Altering a Gene Involved in Nuclear Distribution Increases the Repeat-Induced
Point Mutation Process in the Fungus
Podospora anserina
Khaled Bouhouche, Denise Zickler, Robert Debuchy
1and Sylvie Arnaise
Institut de Ge´ne´tique et Microbiologie, UMR CNRS Universite´ 8621, Universite´ Paris-Sud, F-91405 Orsay Cedex, France Manuscript received September 3, 2003
Accepted for publication January 28, 2004
ABSTRACT
Repeat-induced point mutation (RIP) is a homology-dependent gene-silencing mechanism that intro-duces C:G-to-T:A transitions in duplicated DNA segments.Cis-duplicated sequences can also be affected by another mechanism called premeiotic recombination (PR). Both are active over the sexual cycle of some filamentous fungi, e.g., Neurospora crassaand Podospora anserina. During the sexual cycle, several developmental steps require precise nuclear movement and positioning, but connections between RIP, PR, and nuclear distributions have not yet been established. Previous work has led to the isolation ofami1, theP. anserinaortholog of theAspergillus nidulans apsAgene, which is required for nuclear positioning. We show here thatami1is involved in nuclear distribution during the sexual cycle and that alteration of ami1delays the fruiting-body development. We also demonstrate thatami1alteration affects loss of transgene functions during the sexual cycle. Genetically linked multiple copies of transgenes are affected by RIP and PR much more frequently in anami1mutant cross than in a wild-type cross. Our results suggest that the developmental slowdown of theami1mutant during the period of RIP and PR increases time exposure to the duplication detection system and thus increases the frequency of RIP and PR.
T
HREE homology-dependent gene-silencing mech- fied inN. crassa. Mutations in this gene result in a com-anisms affecting the sexual cycle in Euascomycete plete loss of RIP (Freitag et al. 2002). RID encodes a fungi have been identified: meiotic silencing by unpaired DNA methyltransferase homolog, assumed to be involved DNA (Shiu et al.2001), repeat-induced point mutation in C-to-T transition. InN. crassa, RIP is associated with (RIP), and methylation induced premeiotically (re- PR between tandem repeats (Selkeret al.1987;Irelan viewed byCogoni2001). A fourth mechanism, resulting et al. 1994). PR of tandem repeats has been also de-in gene loss rather than silencde-ing, is based on premeiotic scribed inP. anserina(Picardet al.1987; Coppin-Ray-recombination (PR) betweencis-duplicated sequences and nalet al.1989) but RIP has not been investigated during deletion of the interstitial sequence (reviewed bySelker these studies. PR occurs between fertilization and meio-1990). RIP appears to be a defense genome mechanism sis in P. anserina (Picard et al.1987) and in N. crassa whereby multicopy sequences within one nucleus are (Selkeret al.1987), and the occurrence of RIP has also affected by C-to-T transitions. Although RIP-like muta- been established between these two stages inN. crassa tions are present in almost all identified fungal transpos- (reviewed bySelker1990). Development from fertiliza-able elements (reviewed byDaboussiandCapy2003), tion to meiosis requires tightly regulated nuclear move-functional machinery for RIP has been demonstrated ments (Zickleret al.1995). The relationship between only in Neurospora crassa (Cambareri et al. 1989), Po- homology-dependent gene-silencing mechanisms anddospora anserina(Hamannet al.2000;Graiaet al.2001), the early steps of the sexual cycle, notably nuclear
distri-Magnaporthe grisea(Ikedaet al.2002), andLeptosphaeria bution, has not yet been investigated.
maculans(IdnurmandHowlett2003). RIP character- TheP. anserina ami1gene was previously shown to be
istics have been extensively investigated inN. crassa(re- required for nuclear distribution at different stages of viewed bySelker1990, 2002). RIP operates in a pairwise
the fungal life cycle (Graiaet al.2000). The predomi-fashion on linked and unlinked DNA repeats, but is more
nant phenotype of anami1mutant is male sterility, due efficient on linked duplications. While RIP is frequently
to faulty nuclear migration into male cells (microconi-observed in N. crassa, only two RIP events have been
dia). ami1encodes an ortholog of ApsA ofA. nidulans
reported so far inP. anserina, suggesting that RIP is rare
(FischerandTimberlake1995). ApsA is assumed to be in this species (Hamannet al.2000;Graiaet al.2001).
a positional regulatory protein because the corresponding A single gene (RID) implicated in RIP has been
identi-mutants display nuclear misplacement rather than im-paired nuclear migration (reviewed by Fischer 1999).
ApsAcomplements the male sterility of theami1-1mutant, 1Corresponding author:Institut de Ge´ne´tique et Microbiologie,
Baˆti-suggesting that ApsA/AMI1 may have similar molecular
ment 400, Universite´ Paris-Sud, F-91405 Orsay Cedex, France.
E-mail: [email protected] functions. Recently, a genetic interaction was
strated between apsA and nud F, a gene involved in the dynein pathway, providing the first evidence of a connection between the putative regulatory protein and the cytoplasmic dynein pathway (Efimov2003). How-ever, the effect of AMI1/ApsA alterations on the sexual cycle have not been described.
Analyses of the phenotypes of a partial deletion of
ami1 show that nuclei are abnormally distributed all along the fruiting-body development in this⌬ami1 ho-mozygous cross. Unexpected interactions betweenami1, RIP, and PR were also identified. The frequency of prog-eny affected by RIP and PR was highly increased in ⌬ami1 homozygous crosses and this phenotype corre-lates with a developmental retardation at the early steps of the sexual cycle. These results suggest that RIP and PR phenotypes may be due to an increased exposure time to detection of duplicated sequences during the mutant sexual cycle.
MATERIALS AND METHODS
Plasmids:pCBSMR1 and pPAH1 contain the target genes for measuring the frequency and nature of inactivating events inP. anserina. pCBSMR1 is a derivative of pCB1004 (Carroll
et al.1994) and thehphgene of this vector was used as a target
Figure 1.—Construction of p4-4-5Ble. The p4-5 plasmid gene. It encodes a hygromycin phosphotransferase
determin-(A), which contains a 4.6-kbEcoRI fragment from the ami1 ing the resistance to hygromycin, which serves as the selectable
coding sequence (Graiaet al.2000), has been digested with marker upon transformation. pCBSMR1 also contains the
KpnI andBsaAI. This fragment ofami1has been replaced by SMR1gene from themat⫺mating-type idiomorph (Arnaise
a 1.5-kbKpnI-BsaAI fragment of pPable (CoppinandDebuchy
et al.2001a). The pPAH1 plasmid is based on pUC18 with a
2000) containing a phleomycin resistance gene (Calmelset 3.9-kbPstI genomic fragment fromP. anserinacontaining the
al. 1991) under control of thegpd promoter ofP. anserina pah1gene (Arnaiseet al.2001b).pah1encodes a
homeodo-(RidderandOsiewacz1992), resulting in p4-4-5ble (B). main protein and the inactivation of this target gene in P.
anserinaresults in colonial growth (Arnaiseet al.2001b). p4-4-5ble contains a partial deletion in theami1gene and
was used for replacing the residentami1⫹gene ofP. anserina the binucleate ascospores containmat⫹andmat⫺nuclei and yield self-fertile mycelia. Homokaryotic strains are self-sterile through homologous recombination. The construction of
p4-4-5ble is presented in Figure 1. and must be crossed with a complementary strain. Three kinds
of crosses were used in this study. If mating partners produced Cloning of the complete ami1 gene required the use of
pUN121, a low-copy vector with a positive selection for recom- fertile microconidia, they were cultured in the same petri dish and microconidia were spread with sterile water from one binant (Nilsson et al.1983). An 8-kb fragment containing
the ami1 gene was amplified by PCR with the high-fidelity strain over the other (spermatization). If mating partners were male sterile, either small agar blocks from a culture of each DNA polymerasePfuTurbo(Stratagene, Amsterdam) between
primers N1 (5⬘-CCCTTAAGGTATACGCTGTCCTTTGACAC parent were deposited side by side on the same petri dish and mating occurred after growing at the junction of compatible CAAG-3⬘) and X1 (5⬘-GATGATCAGATCTGTAACCACGCAA
ATCAAGAC-3⬘) from cosmid N10 (Picard et al.1991) and mycelia (side-by-side culture) or the mycelium of one strain was fragmented in a mixer and spread on the mycelium of cloned in theSmaI site of pUN121, resulting in pUN121ami1.
The sequence ofami1from pUN121ami1 indicated that two the compatible strain.
Construction of theami1ⴙ⌬pah1strains:While examining mutations had been introduced during the amplification
pro-cess, leading to the replacement of methionine 939 by a valine the interaction between the homeobox genepah1(Arnaise
et al.2001b) andami1(Graiaet al.2000), we observed that and deletion of a glutamine in a polyglutamine track at
posi-tion 1790. The ami1 gene of pUN121ami1 was noted as fertilization of a double-mutantami1-1⌬pah1by wild-type mi-croconidia resulted in weak sporulation and perithecia with-ami1M939V,⌬Q1790, but these mutations did not impair the function
ofami1in complementation tests. out beak (our unpublished result). This phenotype has been
used for screening hundreds of putative candidates with a P. anserinagrowth conditions, transformation, genetic
analy-sis, and crosses:P. anserinais a Euascomycete whose life cycle knockout of ami1 in a ⌬pah1 context. The deletion of the pah1gene and its replacement by thehphgene in anami1⫹ and general methods for genetic analysis have been described
(Esser1974). Transformations were performed as previously mat⫹ strain has been described previously (Arnaise et al. 2001b). The resulting ami1⫹ mat⫹ ⌬pah1 primary trans-described (Berteaux-Lecellieret al.1995). All strains are
derived from the wild-type S strain. Asci contain four twinned formant was crossed with anami1⫹mat⫺strain, and theami1⫹ mat⫺ ⌬pah1andami1⫹mat⫹ ⌬pah1strains were identified in binucleate ascospores (RajuandPerkins1994). Thus, three
ascospores per ascus are sufficient to determine genetic segre- the progeny. Theami1⫹smr1::ura5 SMR1ec⫹hphec⫹⌬pah1strain
has been constructed as follows. The disruption of theSMR1 gations. A few asci contain three binucleate and two
resultingami1⫹smr1::ura5strain was referred to assmr1-r1in ⌬ami1 mat⫺ ⌬pah1 pah1ec⫹strain was identified by restoration
of the male fertility. The ami1M939V,⌬Q1790 transgene from
the original article;Arnaiseet al.1997).SMR1belongs to the
mat⫺ idiomorph (Debuchy et al. 1993) and its disruption ami1M939V,⌬Q1790 ⌬ami1 mat⫹ was transferred successively by
crossing in⌬ami1 mat⫺and then in⌬ami1 mat⫹ ⌬pah1, re-results in a complete arrest of the sexual cycle (Arnaiseet al.
1997). This defect is complemented by the SMR1 gene of sulting inami1M939V,⌬Q1790⌬ami1 mat⫹ ⌬pah1, which was
identi-fied by restoration of male fertility. pCBSMR1 (Arnaiseet al.2001a). Theami1⫹smr1::ura5strain
has been crossed with a ami1⫹ mat⫹ strain containing DNA procedures:Genomic DNA for PCR or hybridization experiments was extracted according to the CTAB method pCBSMR1 and the progeny have been analyzed for aami1⫹
smr1::ura SMR1ec⫹ hphec⫹ strain. This latter strain has been (RogersandBendich1988). Hybridizations were performed
as described previously (Church and Gilbert 1984). The crossed withami1⫹mat⫹ ⌬pah1 to obtainami1⫹ smr1::ura5
SMR1ec⫹hphec⫹⌬pah1. probes were prepared using the Megaprime DNA labeling
system (Amersham Biosciences). The 1988-bppah1probe was Construction of the⌬ami1strains:The plasmid p4-4-5ble
has been introduced by transformation in an ami1⫹ mat⫹ obtained by PCR with SA13 (5⬘-GGCTACCGACGTGGT CAAGC-3⬘) and SA17 (5⬘-TCCTCTCAAAACGCCGCAG) on ⌬pah1strain and examination of 600 transformants for weak
sporulation and perithecia without beak allowed us to identify wild-type genomic DNA. The 1039-bphphprobe was obtained by PCR with KB13 (5⬘-CCCAAGCATCAAATGAAAAAGCC-3⬘) 81 candidates that displayed this phenotype. Theami1-1
mu-tant was male sterile (Graiaet al.2000), suggesting that the and KB14 (5⬘-CATTTCCTTTGCCCTCGGACGAGTGC-3⬘) on the pCBSMR1 plasmid. To sequencepah1transgenes, we first partial deletion ofami1should have a similar phenotype. We
retained eight male-sterile strains among the 81 transformants amplified their coding sequence from genomic DNA by PCR with SA13 and SA17 or SA13 and SA8 (5⬘-CAGACATGGAG for further examinations. The replacement of a part ofami1
byblewas tested by PCR with two primer pairs. Each pair of AGCGTGTG-3⬘), and the PCR product was purified and se-quenced with one of these primers: SA8, SA13, SA1 (5⬘-CCG primers contain one primer in theblegene, KB4 (5⬘-GAAGGC
TTTAATTTGCAAGC-3⬘) or KB3 (5⬘-GTGGAAGGGAAGGGAT TTCGTGAGAGGATAGCG-3⬘), SA7 (5⬘-GTTCATGCACCGTC ATACCG-3⬘), or KB16 (5⬘-GCGAGAAGCTTGCAGGTCTTC-3⬘). GCTC-3⬘), and the other primer, KB1 (5⬘-CCTGCCAGTCGGA
TCCACGA-3⬘) or KB2 (5⬘-GGGCATCCGCTTGCCATAGC-3⬘), To sequencehphtransgenes, we first amplified their coding sequence with KB13 and KB14, and the PCR product was is targeted to the genomic sequences flanking the 4.6-kbEcoRI
fragment, which contains either the wild-type or the deleted purified and sequenced with one of these primers: KB13,
KB14, hph-s-27 (5⬘-CCACTAGTCTGTCGAGAAGTTTCT
ami1gene. PCR fragments of the expected size were obtained
GATC-3⬘), or hph-a-997 (5⬘-GGACTAGTGGCGTCGGTTTC with the two pairs of primers for 3 transformants, indicating
CACTATCG-3⬘). that the disrupted gene of the p4-4-5ble plasmid has integrated
at its homologous locus by a double crossing over, resulting in the replacement of the wild-typeami1gene by the disrupted
RESULTS copy. The primary transformants were purified by crossing
with a tester strain of opposite mating type andmat⫹
phleo-⌬ami1strains display abnormal nuclear distribution mycin-resistant strains were recovered. The replacement of
and retardation during the sexual cycle:P. anserinais a ami1was tested by Southern blotting in 2 transformants.
Geno-mic DNA was digested withEcoRI and probed with the 8-kb heterothallic fungus with two mating-type idiomorphs,
fragment containing theami1 gene. An ami1⫹ mat⫹ strain mat⫹andmat⫺(Coppinet al.1997). Theami1-1mutant gave three bands at 22, 7.6, and 4.6 kb. The 4.6-kb fragment is tightly linked to themat⫹idiomorph and noami1-1 should be missing in a ⌬ami1 strain and replaced by two
mat⫺strain was obtained previously (Graiaet al.2000).
hybridizing bands of 2 and 1.1 kb. Only one transformant
We constructed⌬ami1 mat⫹and⌬ami1 mat⫺strains by
displayed the expected pattern for a⌬ami1 mat⫹strain. The
procedure described for the construction of⌬ami1 mat⫹was replacing part of theami1coding sequence with theble
followed for the construction of ⌬ami1 mat⫺ and ⌬ami1 gene in mat⫹ and mat⫺ strains (see materials and smr1::ura5 SMR1ec⫹hphec⫹strains after the transformation of
methods). The partial deletion within ami1 left a 3⬘
ami1⫹mat⫺ ⌬pah1andami1⫹smr1::ura5 SMR1ec⫹hphec⫹⌬pah1
end encoding 539 residues of the 1882 deduced from
strains by plasmid p4-4-5ble. Each of these⌬ami1strains was
the entire ami1 gene. Both mutant strains produced
constructed independently, since theami1gene is physically
anucleate microconidia, as does theami1-1mutant,
re-tightly linked to the mating-type locus.
Construction of theami1ⴙstrains containing the hygromy- sulting in male sterility. This defect is complemented cine resistance transgene:The SMR1ec⫹hphec⫹transgenes of
by theami1gene from pUN121ami1 (data not shown).
⌬ami1 smr1::ura5 SMR1ec⫹hphec⫹were introduced after
succes-The absence of functional microconidia in anami1
mu-sive crosses into anami1⫹smr1::ura5strain resulting inami1⫹
tant strain does not preclude mating, because the myce-smr1::ura5 SMR1ec⫹hphec⫹. We checked by Southern blotting
lium can also act as a donor of nuclei to the female
that the SMR1ec⫹ hphec⫹ transgenes were not altered in the
latter strain after the successive crosses. organs of opposite mating type (Graia et al. 2000).
Construction of the strains containing thepah1transgene: Mating between⌬ami1 strains was obtained by spread-The pPAH1 plasmid was introduced by transformation in an ing fragmented mycelium over a sexually compatible ami1⫹mat⫹ ⌬pah1strain, giving rise to theami1⫹mat⫹ ⌬pah1
strain. When the same protocol was used with wild-type pah1ec⫹strain (Arnaiseet al.2001b). Then thepah1ec⫹
trans-strains, ascospore projection occurred 4 days after
fertil-gene was introduced by crossing in a⌬ami1 mat⫺ ⌬pah1strain,
resulting in⌬ami1 mat⫺ ⌬pah1 pah1ec⫹. ization. In a⌬ami1 mat⫹ ⫻ ⌬ami1 mat⫺cross, ascospore Construction of the strains for the complementation test: projection was delayed, beginning the seventh day after Plasmids pUN121ami1 and pUChygro (Ruprich-Robertet fertilization. To determine precisely when delay oc-al.2002) were cotransformed in⌬ami1 mat⫹.
Hygromycin-curred, we did a detailed cytological analysis of the
fruit-resistant (HygR) strains were screened for restoration of male
ing-body development in both⌬ami1and wild-type
ho-fertility, yieldingami1M939V,⌬Q1790 ⌬ami1 mat⫹. This strain was
Figure 2.—Asci and ascospore formation in wild type and⌬ami1.(A) Progressive crozier (ar-rowhead) and ascus development (length of the three arrows indicates the progression from karyo-gamy to pachytene) on an ascogenous hypha in a wild-type perithecium (already published in Zickleret al.1995). (B–E)⌬ami1.(B) Abnormal crozier formation. Some croziers are pluri-nucleated (arrowhead), some contain only one nucleus (thin arrow), and some are anucleated (thick arrow). Note also that the cell below the empty crozier shows three nuclei. (C) Example of abnormal distribution of nuclei during crozier formation. Some cells are plurinucleated (arrow-heads) while others contain one (small arrow) or no nuclei (large arrow). (D) Postmeiotic mitosis with all four spindles formed in the ascus center when, in wild type, the fourspindles are aligned in two well-separated pairs. Large arrow points to a group of chromosomes; small arrow indicates a spindle pole body. (E) Four ascospores: two are binucleated (thick arrows), one contains four nuclei (arrowhead), and one is anucleated (thin arrow, bottom). The top ascus contains only abnormal spores (thin arrow, top). (F) Two wild-type asci. The top ascus contains four binucleate ascospores (large arrows). The bottom ascus (not complete) contains one binucleate and two uninucleate ascospores (small arrows; already published in Zickleret al.1995). Bar, 5m.
do not fuse immediately after fertilization: they divide Progeny of ⌬ami1 homozygous crosses are also conse-quently reduced. A self-fertile heterokaryotic mycelium several times in the female cells beforemat⫹andmat⫺
nuclei migrate into specialized binucleated ascogenous issued from an ascospore containing⌬ami1 mat⫹and
⌬ami1 mat⫺nuclei yielded 70,000 ascospores per petri
hyphae. After a few divisions, the tip cell of the
ascogen-ous hyphae differentiates into a hook-shaped cell, the dish, while a wild-type ami1⫹mat⫺/ami1⫹ mat⫹ strain produced 470,000 ascospores in similar conditions. crozier, in which the two nuclei undergo coordinate
mitosis yielding, after septal formation, two uninucleate ⌬ami1enhances loss of function of transgenes: To assay the frequencies of transgene loss of function, we basal and lateral cells and a binucleate ascus mother
cell (Figure 2A). Karyogamy and meiosis occur in the used the hph and pah1 genes, which encode a hygro-mycin phosphotransferase (Carrollet al.1994) and a ascus mother cell and, after a postmeiotic mitosis,
asco-spores are delineated (for details see Zickler et al. P. anserina homeodomain protein (Arnaise et al.
2001b), respectively. The structure ofpah1andhph ec-1995). In a wild-type perithecium, croziers and the first
meiotic-prophase asci appear 48 hr after fertilization topic integrations were analyzed by Southern blotting: they contained at least three copies ofpah1(Figure 3) (Figure 2A), and by 72 hr, perithecia contain croziers,
asci at different stages of meiosis, and a few asci with and two copies ofhph(data not shown). The multiple copies ofpah1remained associated and identical to the just membraned ascospores. Ascospore projection starts
4 days after fertilization. In homozygous ⌬ami1 peri- parental pattern in 19 independent ascospores resulting from anami1⫹mat⫺ ⌬pah1⫻ami1⫹mat⫹ ⌬pah1 pah1ec⫹ thecia, the first croziers and prophases occur 72 hr after
fertilization, indicating at least a 24-hr developmental cross (data not shown). The multiple copies ofhph re-mained associated and identical to the parental pattern delay compared to that of wild type. The first ascospores
were projected 4–5 days later. When compared to wild in 16 independent ascospores resulting from a cross be-tweenami1⫹ smr1::ura5 SMR1ec⫹hphec⫹ andami1⫹ mat⫹ type, this indicates that meiosis and spore formation
are also delayed by 2–3 days: in wild type, ascospore (data not shown). During this analysis, we found that one copy ofhphwas missing in one strain. However, all projection starts 2 days after crozier formation. These
data suggest that mutant development was delayed at ascospores issued from the same ascus also displayed the loss of onehphcopy, indicating unequivocally that all stages of the sporulation process. In parallel with
this delay, we observed abnormal nuclear distribution this loss of onehphcopy was a premeiotic deletion event rather than a segregation event. These data indicated before and during crozier formation (Figure 2, B and
C). Moreover, nuclear distribution is also very irregular that the multiple copies of each transgene were and remained genetically linked in anami1⫹ background. during both meiotic and postmeiotic divisions (Figure
2D), leading to the formation of abnormal ascospores We next analyzed the transgene stabilities in a⌬ami1
genetic background. The inactivation ofhphwas tested (Figure 2E) when compared to wild type (Figure 2F).
Each developmental step is likely delayed and/or defec- in a⌬ami1 smr1::ura5 SMR1ec⫹hphec⫹⫻ ⌬ami1 mat⫹cross. The progeny of an ami1⫹ smr1::ura5 SMR1ec⫹ hphec⫹ ⫻ tive because of nuclear misplacement, and probably
Figure3.—Analysis ofpah1 se-quence rearrangements in the progeny from a ⌬ami1 homozy-gous cross. Six asci (designated by their reference numbers 9, 13, 14, 29, 35, and 37) from the ⌬ami1 mat⫺ ⌬pah1 pah1ec⫹⫻ ⌬ami1 mat⫹
⌬pah1cross (Table 3, type I) were analyzed by Southern blot and compared to the parental hybriza-tion pattern (lane P). The four strains issued from the four asco-spores of each ascus are indicated as a, b, c, and d. DNA was digested withBstBI, which cuts once within thepah1gene, transferred to ny-lon membranes, and probed with apah1fragment corresponding to the coding sequence to detect ectopicpah1transgenes and their rearrangements. All strains issued from the four ascospores of each ascus display identical rearrangements indicating that they occurred premeiotically. For one strain (14d), DNA extraction has failed; however, this does not preclude the analysis of the ascus (seematerials and methods). Thepah1transgenes were amplified from genomic DNA of each one of these strains and sequenced. In asci 13, 14, 29, and 37, identical C:G-to-T:A transitions were found in the four strains issued from the four ascospores of the same ascus, indicating that they occurred premeiotically. The patterns of mutation were specific to each ascus and the number of RIP mutations for asci 13, 14, 29, and 37 were 15, 56, 4, and 19, respectively. Strains from asci 9 and 35 did not display any mutation.
regation of the hph marker, but unexpectedly both complementation of transgene loss of function. Both crosses produced three types of tetrads: those displaying crosses exhibited a third type of tetrads consisting only
of hygromycin-sensitive (HygS) ascospores. Moreover, first- or second-division segregation of thepah1 trans-gene and tetrads containing four ascospores giving rise the frequency of these HygS tetrads was higher in the
⌬ami1homozygous cross than in theami1⫹ cross. The to colonial growth (Table 2). The frequency of these tetrads, which was probably due to a loss of function of increase of HygS tetrads in a ⌬ami1homozygous cross
cannot be calculated confidently at the first day of asco- thepah1transgene, was 18 times higher in⌬ami1than inami1⫹homozygous crosses at the first day of projec-spore projection, but HygS tetrads frequency is seven
times higher in the ⌬ami1 homozygous cross than in tion. The frequency of tetrads with four colonial strains increased in anami1⫹cross and their frequencies were theami1⫹cross at the fifth day (Table 1).
To further test the increased loss of function of a similar after the fourth day of ascospore projection in ⌬ami1 andami1⫹ homozygous crosses.
transgene in a⌬ami1 background, we usedpah1, aP.
anserinahomeobox gene, whose deletion results in colo- We tested whether this phenotype resulted fromami1
mutationper seby complementation experiments with nial growth (Arnaiseet al.2001a). The loss of function
of the pah1 transgenes was scored in a cross between the ami1M939V,⌬Q1790gene from plasmid pUN121ami1. A total of 80 tetrads resulting from a cross between
⌬ami1 mat⫺ ⌬pah1 pah1ec⫹ and ⌬ami1 mat⫹ ⌬pah1
strains and compared with the referenceami1⫹ mat⫺ ami1M939V,⌬Q1790 ⌬ami1 mat⫹ ⌬pah1 and ami1M939V,⌬Q1790
⌬ami1 mat⫺ ⌬pah1 pah1ec⫹ strains were analyzed for
⌬pah1 ⫻ ami1⫹ mat⫹ ⌬pah1 pah1ec⫹ cross. All strains
were deleted for the endogenous pah1 gene to avoid loss of function ofpah1ec⫹
at the first day of ascospore
TABLE 1
Effect ofami1partial deletion on loss of function in linked copies of thehphhygromycin phosphotransferase gene during the sexual cycle ofP. anserina
% loss-of-function frequency ata
Relevant genotype Day 1 Day 5 Day 6
ami1⫹hphec⫹⫻ami1⫹b 0 (0/30) 10 (3/30) 69 (29/42)
⌬ami1 hphec⫹⫻ ⌬ami1c 25 (5/20) 72 (21/29) 89 (17/19)
aTetrads of crosses described in the text were harvested at days 1, 5, and 6 of ascospore projection, analyzed
on growth medium containing hygromycin, and loss-of-function frequency determined as (4 HygS:0 HygR/
total)⫻100.
TABLE 2
Effects ofami1partial deletion on loss of function in linked copies of thepah1homeobox gene during the sexual cycle ofP. anserina
% loss-of-function frequency ata
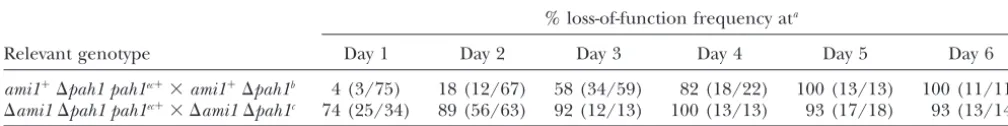
Relevant genotype Day 1 Day 2 Day 3 Day 4 Day 5 Day 6
ami1⫹⌬pah1 pah1ec⫹⫻ami1⫹⌬pah1b 4 (3/75) 18 (12/67) 58 (34/59) 82 (18/22) 100 (13/13) 100 (11/11)
⌬ami1⌬pah1 pah1ec⫹⫻ ⌬ami1⌬pah1c 74 (25/34) 89 (56/63) 92 (12/13) 100 (13/13) 93 (17/18) 93 (13/14) aTetrads of crosses described in the text were harvested from day 1 to day 6 of ascospore projection, analyzed on growth
medium, and loss-of-function frequency determined as (4 colonial:0 wild-type growth/total)⫻100.
bAscospore projection began 4 days after spermatization (seematerials and methods). cAscospore projection began 10 days after side-by-side culture (seematerials and methods).
projection. Of these tetrads, 5% contained four asco- appearance of new bands, suggesting either that RIP has altered the restriction site used for the digestion spores germinating with a colonial growth, a frequency
similar to the frequency obtained in a wild-type cross of the genomic DNA or that a rearrangement, possibly combined with a deletion, has occurred (Figure 3, asci 13 (see Table 2). This complementation test confirmed
that the loss of function is due to⌬ami1. and 14). Strains from asci 9 and 35 (Figure 3) did not display any RIP mutations in the pah1 coding region
⌬ami1increases both RIP and premeiotic
recombina-tion frequency:InP. anserina, two transgene inactivation but nevertheless showed differences in parental hybrid-ization pattern. RIP or PR were also found in thepah1
mechanisms have been described: RIP (Hamannet al.
2000; Graia et al. 2001) and PR (Picard et al. 1987; ectopic locus in 5 of 10 strains growing with a wild-type phenotype and issued from a⌬ami1homozygous cross Coppin-Raynalet al.1989). To determine which
mech-anism accounted for the increase of transgene inactiva- (Table 3, type II). Therefore, 75% (9/12) of colonial tetrads and 20% (2/10) of tetrads with wild-type growth tion in⌬ami1homozygous crosses, we analyzed the
mo-lecular events leading to the loss of function of the displayed RIP. As colonial and wild-type growing tetrads represented 74 and 26% of the progeny on the first day transgene by sequencing and Southern blotting.
Thepah1ectopic locus was analyzed in 12 ascospores of ascospore projection (Table 2), the frequency of RIP in the progeny at this stage can be estimated at 60%. from independent asci in a ⌬ami1 homozygous cross
on the first and second day of projection. We amplified A similar rationale indicates that PR affected 75% of the progeny of a ⌬ami1 homozygous cross at the first the pah1 transgenes from the genomic DNA and the
PCR product was sequenced. Nine strains obtained from day of ascospore projection. As reference, we analyzed the molecular events occurring at thepah1ectopic locus the 12 ascospores displayed transgene mutations, all
restricted to C:G-to-T:A transitions (Table 3, type I). in the progeny of a ami1⫹ homozygous cross. Three strains issued from three different tetrads with genetic For 8 strains among the 9 RIPed strains, RIP mutations
corresponded to superimposed T and C or G and A evidence of loss ofpah1function contained a rearranged
pah1 sequence and one of them has undergone RIP peaks on the electrophoregram, suggesting that at least
two copies ofpah1have been amplified. Southern blot (Table 3, type III). In contrast, the 19 analyzed strains exhibiting a wild-type growth phenotype did not display analysis of genomic DNA from the 12 strains indicated
that 11 strains have undergone transgene deletion or RIP or rearrangements (Table 3, type IV). The fre-quency of RIP and PR in an ami1⫹ cross is therefore rearrangements. To test whether identical mutations
and deletion or rearrangements could be found in the 1.3 and 4%, respectively.
Furthermore, we have analyzed thehphmarker in the other ascospores from the same tetrad, all ascospores
of 6 of the 12 tetrads were also examined. All ascospores progeny obtained from a⌬ami1homozygous cross and from a referenceami1⫹ cross. In both crosses, all nine of each tetrad displayed identical mutations (data not
shown) and identical Southern blot patterns (Figure 3), tested strains issued from tetrads with exclusively HygS ascospores contained a RIPedhphsequence, and six of indicating that these events occurred premeiotically, as
expected for RIP and PR. Previously, PR was found only them also showed PR (Table 3, types V and VII). A single HygR strain issued from a ⌬ami1 homozygous on a 40-kb tandem duplication inP. anserina
(Coppin-Raynal et al. 1989). We show here that duplications cross was analyzed (Table 3, type VI). It displayed one RIP mutation. Three HygRstrains issued from anami1⫹ obtained with smaller inserts can also be the target of
PR. In some cases, PR resulted in the loss of one or cross showed no RIP and no PR (Table 3, type VIII). These results demonstrate that the high efficiency of several bands, suggesting that a deletion of transgenes
TABLE 3
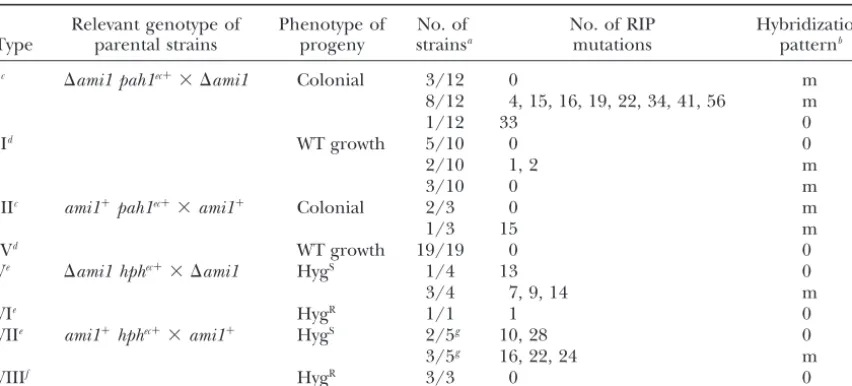
Molecular characterization of the events leading to the loss of function ofpah1andhphtransgenes in⌬ami1andami1ⴙhomozygous crosses
Relevant genotype of Phenotype of No. of No. of RIP Hybridization
Type parental strains progeny strainsa mutations patternb
Ic ⌬ami1 pah1ec⫹⫻ ⌬ami1 Colonial 3/12 0 m
8/12 4, 15, 16, 19, 22, 34, 41, 56 m
1/12 33 0
IId WT growth 5/10 0 0
2/10 1, 2 m
3/10 0 m
IIIc ami1⫹pah1ec⫹⫻ami1⫹ Colonial 2/3 0 m
1/3 15 m
IVd WT growth 19/19 0 0
Ve ⌬ami1 hphec⫹⫻ ⌬ami1 HygS 1/4 13 0
3/4 7, 9, 14 m
VIe HygR 1/1 1 0
VIIe ami1⫹hphec⫹⫻ami1⫹ HygS 2/5g 10, 28 0
3/5g 16, 22, 24 m
VIIIf HygR 3/3 0 0
aAscospores were collected at the first and second day of ascospore projection, unless stated otherwise. bDifference from the parental pattern is indicated as identical to parental (0) or modified (m).
cpah1transgenes were amplified with primers SA13 and SA8, and 800 bp of thepah1coding sequence were
determined on each strain by sequencing the PCR products with SA13 and SA8.
dpah1transgenes were amplified with primers SA13 and SA17, and 1500 bp of thepah1coding sequence
were determined on each strain by sequencing the PCR products with primers KB16, SA1, and SA7.
ehphtransgenes were amplified with primers KB13 and KB14 and 900 bp of thehphcoding sequence were
determined on each strain by sequencing the PCR products with primers Spehph-s-27 and Spehph-a-997.
fhphtransgenes were amplified with primers KB13 and KB14 and 500 bp of thehphcoding sequence were
determined on each strain by sequencing the PCR products with KB13.
gAscospores collected at 5 and 6 days after the beginning of ascospore projection.
⌬ami1cross, 60% of tetrads were affected by RIP in the formation has been available in this fungus for 15 years. Three features may explain why RIP has gone
unde-pah1gene as compared to 1.3% in the ami1⫹ cross on
the first day of ascospore production, thus a 46-fold tected inP. anserinafor such a long time. First, we have been unable to detect RIP on unlinked copies of the increase. PR affected 75% of the tetrads in a⌬ami1cross
while 4% of the tetrads were affected by this event in a pah1 gene (data not shown), even in a ⌬ami1 back-ground, which clearly increases the probability of RIP.
ami1⫹ cross, indicating a 19-fold increase in a ⌬ami1
background. Although the sampling was less extensive This observation suggests that RIP on unlinked copies is very inefficient inP. anserina, reducing the chance to for the loss of function of the hph gene, RIP and PR
were clearly increased in a⌬ami1cross as compared to detect a RIP event. Second, after the introduction of a transgene inP. anserina, any transformant not displaying a wild-type cross. The frequency of RIP mutations
can-not be calculated accurately, since the exact number of an active transgene is usually discarded. Finally, most of the genetic work onP. anserinais performed on five transgene copies in each strain is not known. Only an
overestimation of RIP frequency can be calculated. The ascospore asci, which are produced predominantly at the beginning of ascospore projection, at a time of mini-average RIP nucleotide mutation found in an ami1⫹
context is 1.8 and 2% forpah1andhphgenes, respec- mal RIP efficiency.
Previous work indicated that the ami1 gene was in-tively. The average RIP nucleotide mutation in a⌬ami1
context is 3 and 1% forpah1andhphgenes, respectively. volved in nuclear distribution and positioning (Graia
et al. 2000). This study shows that it is also involved
during the sexual cycle. Interestingly, as ⌬ami1 was
DISCUSSION
found to increase RIP and PR efficiency, it raises the question of the relation between nuclear distribution, We show here that RIP, which was considered rare in
P. anserina (Graia et al. 2001), can be easily detected RIP, and PR. Abnormal nuclear positioningper semight
trigger high RIP and PR, suggesting that the level of RIP on linked multiple sequences at the end of ascospore
projection in aami1⫹strain. It is thus surprising that only and PR might be directly controlled by developmental events. Several lines of evidence indicate that the in-two RIP events have been thus far detected inP. anserina
indirect consequence of abnormal nuclear distribution. expelled ascospores are likely to correspond to croziers observed at 72 hr in the wild-type cross and thus could We have demonstrated that development is delayed in
⌬ami1 homozygous crosses, notably before meiosis, have been submitted to a more prolonged detection of duplication. This modulation of the RIP efficiency which marks the end of RIP and PR occurrence. This
correlation suggests that the developmental delay of between early- vs. late-expelled ascospores may allow duplications to escape RIP in early progeny while they a ⌬ami1 homozygous cross increases the exposure of
duplicated sequences to RIP and PR, resulting in a would be inactivated in late progeny, as proposed by Singer et al. (1995). Such variation in RIP efficiency greater frequency of these events in the progeny.
Ac-cording to this rationale, we predict that any mutations could favor the selection of beneficial duplication in early progeny and the inactivation of detrimental dupli-that slow down initial fruiting-body development should
increase RIP and PR efficiency. Accordingly, the high cation in late progeny. This mechanism and the very low efficiency of RIP on unlinked duplication lead us efficiency of RIP inN. crassamay be attributed, at least
partially, to a specific developmental feature. In N. to predict that RIP has not retarded the creation of new genes through genomic duplication in P. anserina, in
crassa, the first ascospores were found 9–10 days after
fertilization (Singer et al.1995), in contrast to 4 days contrast to its effect inN. crassa(Galaganet al.2003). The comparative genomic analysis from N. crassaand after fertilization in wild-typeP. anserina. This
develop-mental delay in N. crassa may have similar effects on P. anserinawill thus be interesting and may allow us to assess more precisely the impact of various RIP modal-RIP as did the⌬ami1mutation inP. anserina, resulting
in a high proportion of RIPed progeny. ities on genome evolution of the two fungi.
In contrast to the dramatically increased frequency We are greatly indebted to Fatima Graı¨a and Ve´ronique Berteaux-of asci affected by RIP in a ⌬ami1 background, the Lecellier for the gift of p4-5 and for communicating theami1 unpub-lished sequence data. We are grateful to Marguerite Picard for her
frequency of RIP mutation is not modified by ⌬ami1.
constant interest in this work. We thank our colleagues for helpful
The previously reported average RIP frequency in P.
discussions. K.B. was supported by scholarship no. 00302 of the
Minis-anserinawas 1% (Graiaet al.2001), similar to the RIP
te`re de la Recherche et des Nouvelles Technologies.
frequency found in this study, in eitherami1⫹or⌬ami1
backgrounds. RIP frequency may be limited by the fact that PR might result in the loss of the duplicated
se-LITERATURE CITED
quences required for RIP. According to this hypothesis,
Arnaise, S., R. DebuchyandM. Picard, 1997 What is abona fide
the strains that have conserved the parental
multiple-mating-type gene? Internuclear complementation of mat mutants
band pattern should display a very high mutation rate
inPodospora anserina.Mol. Gen. Genet.256:169–178.
in⌬ami1crosses; however, these strains do not show a Arnaise, S., D. Zickler, S. Le Bilcot, C. PoisierandR. Debuchy,
2001a Mutations in mating-type genes of the heterothallic
fun-higher mutation rate than strains that have undergone
gusPodospora anserinalead to self-fertility. Genetics159:545–556.
PR (see Table 3). Another explanation can account for
Arnaise, S., D. Zickler, C. PoisierandR. Debuchy, 2001b pah1:
the absence of mutation increase in ⌬ami1, in agree- a homeobox gene involved in hyphal morphology and microconi-ment with a model proposed by Selker (1990). The diogenesis in the filamentous ascomycetePodospora anserina.Mol.
Microbiol.39:54–64.
detection of duplication may take place during the G1
Berteaux-Lecellier, V., M. Picard, C. Thompson-Coffe, D.
phase, which could be lengthened as a consequence of Zickler, A. Panvier-Adoutteet al., 1995 A nonmammalian development delay in a⌬ami1cross. Consequently, the homolog of thePAF1gene (Zellweger syndrome) discovered as a gene involved in caryogamy in the fungusPodospora anserina.
number of progeny affected by RIP is expected to
in-Cell81:1043–1051.
crease, as observed in a⌬ami1cross. The mutation pro- Calmels, T., M. Parriche, H. DurandandG. Tiraby, 1991 High cess may occur during the nuclear division, the length efficiency transformation of Tolypocladium geodesconidiospores
to phleomycin resistance. Curr. Genet.20:309–314.
of which should be unaffected by development delay.
Cambareri, E. B., B. C. Jensen, E. SchabtachandE. U. Selker,
If there are no more nuclear divisions during the early 1989 Repeat-induced G-C to A-T mutations inNeurospora. Sci-development stages of the fruiting body in a ⌬ami1 ence244:1571–1575.
Carroll, A. M., J. A. SweigardandB. Valent, 1994 Improved
homozygous cross than there are in a wild-type cross,
vectors for selecting resistance to hygromycin. Fungal Genet.
no increase of mutation is expected in a⌬ami1cross. Newsl.41: 22.
If⌬ami1increases RIP and PR by a developmental retar- Church, G. M., andW. Gilbert, 1984 Genomic sequencing. Proc. Natl. Acad. Sci. USA81:1991–1995.
dation, it should be possible to find some conditions in
Cogoni, C., 2001 Homology-dependent gene silencing mechanisms
which fruiting-body development was delayed and RIP/
in fungi. Annu. Rev. Microbiol.55:381–406.
PR were increased. InP. anserina, as inN. crassa(Singer Coppin, E., andR. Debuchy, 2000 Co-expression of the mating-type genes involved in internuclear recognition is lethal inPodospora
et al. 1995), RIP is increased in late-expelled ascospores
anserina.Genetics155:657–669.
relative to early ascospores. These late-expelled ascospores
Coppin, E., R. Debuchy, S. ArnaiseandM. Picard, 1997 Mating
in a wild-type cross could be considered as phenocopies types and sexual development in filamentous ascomycetes.
Micro-biol. Mol. Biol. Rev.61:411–428.
of the early-projected ascospores in a ⌬ami1
homozy-Coppin-Raynal, E., M. PicardandS. Arnaise, 1989
Transforma-gous cross. In both cases, detection of transgene
duplica-tion by integraduplica-tion inPodospora anserina. III. Replacement of a
tion is increased while RIP mutation efficiency remains chromosome segment by a two-step process. Mol. Gen. Genet.
219:270–276.
late-Daboussi, M.-J., andP. Capy, 2003 Transposable elements in fungi. Nilsson, B., M. Uhlen, S. Josephson, S. GatenbeckandL.
Phil-Annu. Rev. Microbiol.57:275–299. ipson, 1983 An improved positive selection plasmid vector
con-Debuchy, R., S. ArnaiseandG. Lecellier, 1993 Themat⫺allele structed by oligonucleotide mediated mutagenesis. Nucleic Acids
ofPodospora anserinacontains three regulatory genes required Res.11:8019–8030.
for the development of fertilized female organs. Mol. Gen. Genet. Picard, M., R. Debuchy, J. JulienandY. Brygoo, 1987
Transforma-241:667–673. tion by intergration inPodospora anserina. II. Targeting to the
Efimov, V. P., 2003 Roles of NUDE and NUDF proteins ofAspergillus resident locus with cosmids and instability of the transformants.
nidulans: insights from intracellular localization and overexpres- Mol. Gen. Genet.210:129–134.
sion effects. Mol. Biol. Cell14:871–888. Picard, M., R. DebuchyandE. Coppin, 1991 Cloning the mating
Esser, K., 1974 Podospora anserina, pp. 531–551 inHandbook of Genet- types of the heterothallic fungus Podospora anserina:
develop-ics, edited byR. C. King. Plenum, New York/London. mental features of haploid transformants carrying both mating
Fischer, R., 1999 Nuclear movement in filamentous fungi. FEMS types. Genetics128:539–547.
Microbiol. Rev.23:39–68. Raju, N. B., andD. D. Perkins, 1994 Diverse programs of ascus
Fischer, R., andW. E. Timberlake, 1995 Aspergillus nidulans apsA development in pseudohomothallic species ofNeurospora,
Gelasi-(anucleate primary sterigmata) encodes a coiled-coil protein re- nospora, andPodospora.Dev. Genet.15:104–118.
quired for nuclear positioning and completion of asexual devel- Ridder, R., andH. D. Osiewacz, 1992 Sequence analysis of the gene opment. J. Cell Biol.128:485–498. coding for glyceraldehyde-3-phosphate dehydrogenase (gpd) of
Freitag, M., R. L. Williams, G. O. KotheandE. U. Selker, 2002 Podospora anserina: use of homologous regulatory sequences to
A cytosine methyltransferase homologue is essential for repeat- improve transformation efficiency. Curr. Genet.21:207–213. induced point mutation inNeurospora crassa.Proc. Natl. Acad. Rogers, S. O., andA. J. Bendich, 1988 Extraction of DNA from Sci. USA99:8802–8807. plant tissues, pp. 1–10 inPlant Molecular Biology Manual, edited
Galagan, J. E., S. E. Calvo, K. A. Borkovich, E. U. Selker, N. D.
by S. B.Gelvin, R. A.Schilperoortand D. P. S.Verma. Kluwer
Read et al., 2003 The genome sequence of the filamentous
Academic Publishers, Dordrecht, The Netherlands. fungusNeurospora crassa.Nature422:859–868.
Ruprich-Robert, G., D. Zickler, V. Berteaux-Lecellier, C. Velot
Graia, F., V. Berteaux-Lecellier, D. ZicklerandM. Picard, 2000
andM. Picard, 2002 Lack of mitochondrial citrate synthase
ami1, an orthologue of theAspergillus nidulans apsAgene, is
in-discloses a new meiotic checkpoint in a strict aerobe. EMBO J. volved in nuclear migration events throughout the life cycle of
21:6440–6451.
Podospora anserina.Genetics155:633–646.
Selker, E. U., 1990 Premeiotic instability of repeated sequences in
Graia, F., O. Lespinet, B. Rimbault, M. Dequard-Chablat, E.
Cop-Neurospora crassa.Annu. Rev. Genet.24:579–613.
pinet al., 2001 Genome quality control: RIP (repeat-induced
Selker, E. U., 2002 Repeat-induced gene silencing in fungi. Adv.
point mutation) comes toPodospora.Mol. Microbiol.40:586–595.
Genet.46:439–450.
Hamann, A., F. FellerandH. D. Osiewacz, 2000 The degenerate
Selker, E. U., E. B. Cambareri, B. C. Jensenand K. R. Haack,
DNA transposonPatand repeat-induced point mutation (RIP)
1987 Rearrangement of duplicated DNA in specialized cells of inPodospora anserina.Mol. Gen. Genet.263:1061–1069.
Neurospora.Cell51:741–752.
Idnurm, A., andB. J. Howlett, 2003 Analysis of loss of
pathogenic-Shiu, P. K., N. B. Raju, D. ZicklerandR. L. Metzenberg, 2001
ity mutants reveals that repeat-induced point mutations can occur
Meiotic silencing by unpaired DNA. Cell107:905–916. in the Dothideomycete Leptosphaeria maculans. Fungal Genet.
Singer, M. J., E. A. Kuzminova, A. Tharp, B. S. MargolinandE. U.
Biol.39:31–37.
Ikeda, K., H. Nakayashiki, T. Kataoka, H. Tamba, Y. Hashimoto Selker, 1995 Different frequencies of RIP among early vs. late
et al., 2002 Repeat-induced point mutation (RIP) inMagnaporthe ascospores ofNeurospora crassa.Fungal Genet. Newsl.42:74–75.
grisea: implications for its sexual cycle in the natural field context. Zickler, D., S. Arnaise, E. Coppin, R. DebuchyandM. Picard, Mol. Microbiol.45:1355–1364. 1995 Altered mating-type identity in the fungusPodospora
anse-Irelan, J. T., A. T. HagemannandE. U. Selker, 1994 High fre- rina leads to selfish nuclei, uniparental progeny and haploid
quency repeat-induced point mutation (RIP) is not associated meiosis. Genetics140:493–503. with efficient recombination in Neurospora. Genetics138:1093–