ABSTRACT
CORDOVA NOBOA, HERNAN ALEJANDRO. Understanding Corn Variability for Broilers. (Under the direction of Dr. Edgar O. Oviedo Rondón).
Corn kernel hardness is an important agronomic trait that could be affected by
environmental conditions and post-harvest practices such as drying and storage. This
dissertation discuss current literature related to corn variability factors and original research
showing the effects of two corn hybrids varying in endosperm hardness (average vs. hard)
and three post-harvest drying temperatures (35, 80, and 120°C) on corn nutritional
composition, geometric mean (dgw) and standard deviation (Sgw) of particle size, and pellet
quality. Likewise, the effect of storage time on the parameters previously listed will be
presented. The supplementation of diets containing these corn hybrids with an exogenous
amylase was also evaluated, to investigate its effects on broiler live performance, organ
development, nutrient digestibility and energy utilization.
In the first chapter the literature presents an overview of factors that influence corn
nutritional value with emphasis in kernel hardness, and feed manufacturing processes such as
grain drying temperature and grinding. The second chapter describes the differences on corn
nutritional composition among two corn hybrids dried at three temperatures, analyzed by
different NIRS prediction models. The non-standardized spectra from whole grain and
ground samples were obtained at two storage times (2 and 24 wk). It was concluded that
endosperm hardness and drying temperatures influenced the nutritional value of corn.
Likewise, storage time increased the estimated value of crude protein and reduced starch
content in all treatments except for corn with average kernel hardness and dried at 120°C. In
roller mill or hammermill at different grinding settings. The effects of kernel hardness and
drying temperatures on dgw and Sgw were studied. This evaluation was conducted in fresh
corn (less than 2 wk of storage) and at 30 wk of storage. Results showed a larger dgw for corn
with harder endosperm and dried at 120°C. This study also demonstrated that as storage time
increase the dgw is reduced regardless of grinding method. For chapter four, the
supplementation with an exogenous amylase on broiler live performance, nutrient
digestibility and energy utilization was investigated. For this study a factorial arrangement of
two corn kernel hardness, three drying temperatures, and two amylase supplementation (0 vs.
133 g/ton) levels were considered. Two parallel studies were carried out in 96 floor pens and
cages. Live performance results showed that at 40d the best FCR was observed when the
average hardness kernels were dried at 120°C. Additionally, the supplementation with
amylase improved FCR in 2 points, but only when feeding diets comprised by corn with
average hardness.
Considering that there were no improvements when supplementing amylase for chickens
fed hard kernel corn, and that corn dried at 120°C yielded larger dgw, a second experiment
was conducted. The effects of hard kernel corn dried at two temperatures (35 vs. 120°C),
ground at two particle sizes (coarse and fine), and three amylase inclusion levels (0, 133, and
266 g/ton) were evaluated on live performance, organ development, nutrient digestibility, and
energy utilization. Results showed that high drying temperature depressed ileal starch
digestibility especially for young chicks. Feeding coarse corn did not improve live
performance, but it promoted gizzard development. Finally, at 42d; FCR was improved when
The results obtained in these experiments suggested that endosperm hardness, and drying
temperatures influenced corn nutritional composition, dgw and Sgw. Likewise, it affected
broiler live performance, nutrient digestibility and energy utilization. Some improvements
were observed on these parameters by adding an exogenous amylase in the feed. However,
these factors were not independent and so must be manipulated strategically to improve corn
© Copyright 2019 by Hernán Alejandro Córdova Noboa
Understanding Corn Variability for Broilers
by
Hernán Alejandro Córdova Noboa
A dissertation submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the Degree of
Doctor of Philosophy
Animal and Poultry Science
Raleigh, North Carolina
2019
APPROVED BY:
___________________________ ___________________________
Dr. Edgar O. Oviedo Rondón Dr. Peter Ferket
Co-Chair of Advisory Committee Co-Chair of Advisory Committee
___________________________ ___________________________
DEDICATION
I dedicate this dissertation to my beloved parents Bolivar Cordova and Cecilia Noboa who
supported and helped me in each step of the way. Also, to my brothers Juan Carlos and Erik
for being my constant motivation. Lastly, I would like to dedicate this dissertation to my
BIOGRAPHY
Hernan Alejandro Cordova Noboa is the first of the three siblings of Bolivar Cordova and
Cecilia Noboa. He was born in Quito-Ecuador and lived there for 29 years before he moved to
Raleigh-North Carolina to continue his education. He obtained his Bachelor of Science degree
with a Major in Farming Science from the Army University of Ecuador in 2011. At the same
year he started working with the National Food Processor Company – PRONACA in the
department of Animal Nutrition as an assistant. He received a Specialist in Animal Nutrition
graduate degree in 2015 from Central University of Ecuador. In August 2015, he was accepted
to start his master’s degree in Poultry Science at North Carolina State University where he joined Dr. Oviedo’s lab as a graduate student and research assistant. In May 2017, he received
his Master of Science diploma and he started his doctorate program in September of the same
year. The Graduate Certificate in Feed Science was awarded to him in December 2017. He
unconditionally passed his dissertation defense in October 2019 and is waiting to receive his
diplomas of Doctor of Philosophy in Animal and Poultry Science, and Nutrition. After his
ACKNOWLEDGMENTS
First, I would like to thank God for my life and the people that is included in it. I would
like to thank my committee members for all their expertise, comments and suggestions for the
present study. A special thanks to Dr. Edgar Oviedo for being my advisor and to accept me as
part of his lab so I could have had this amazing personal and professional experience. Also, I
want to thank Dr. Peter Ferket, Dr. Adam Fahrenholz, and Dr. Consuelo Arellano, for their
precious time expended in reading, commenting and expressing their thoughts through the
entire process. I would like to express my gratitude to all of you for all the support and for your
patience.
I would like to acknowledge and thank my family at the Prestage Department of Poultry
Science for the help and friendship during these two years. A special acknowledgment to all
the staff members of the Chicken Education Unit (Ryan, Lorie, Maddie, and Jeff), and the Feed
Mill Facility for their assistance during my research trials and their continued support and help.
I am very grateful to all the undergrad (Baxter Fernandez, Benjamin Herndon, Charity
Robinett, Alexandria Blanchard, Katherina Wilhelm, Lauren Barker, Zack Bellomo, and
Abigail Dupuis), visitor scholars (Lina Peñuela, Miguel Chico, Yilmar Matta, Andres Ortiz,
Juan David Martinez, Daniela Buitrago, Sebastian Hoyos, Jose Yanquen, and Lorena
Castellanos), research associates (Ramon Malheiros, Camilo Ospina, Deellila Hodgson, and
Rasha Qudsieh) and Faculty (Kimberly Livingston) for their help through this whole time.
Finally, I would like to thank the companies that helped with this project: AB Vista,
DSM, Adisseo, Cargill Inc., Pegasus Science, Trouw Nutrition, Evonik and a special
acknowledgement to the North Carolina Department of Agriculture NCDA & CS Cherry
TABLE OF CONTENTS
LIST OF TABLES ... viii
LIST OF FIGURES ... xiv
CHAPTER I: Literature Review ...1
1.1. Introduction ...1
1.2. Genetic selection ...2
1.3. Maize (Dent corn) ...3
1.4. Kernel Hardness ...4
1.5. Vitreousness and corn quality ...6
1.6. Carbohydrates ...7
1.7. Starch ...7
1.8. Corn as feedstuff ...8
1.9. Nutritional importance and quality of corn as a feedstuff ...8
1.10. Starch dynamics of digestion in poultry ...9
1.11. Processing of corn in feed manufacturing ...11
1.12. Drying process ...11
1.13. Particle size reduction ...14
1.14. Agglomeration - pelleting ...16
1.15. Near-infrared spectroscopy (NIRS) in the feed industry ...17
1.16. Use of exogenous carbohydrases ...19
1.17. References ...22
CHAPTER II: NIRS analysis to assess the nutritional value of two corn hybrids dried at different temperatures: a comparative analysis of different calibration models ...33
2.1. Abstract ...33
2.2. Introduction ...35
2.3. Materials and methods ...38
2.4. Results ...40
2.5. Discussion ...47
2.6. Conclusion ...52
2.7. Acknowledgements ...53
CHAPTER III: Effects of corn kernel hardness and grain drying conditions on particle
size when grinding using a roller mill or hammermil ...97
3.1. Abstract ...97
3.2. Introduction ...99
3.3. Materials and methods ...101
3.4. Results ...106
3.5. Discussion ...110
3.6. Conclusion ...114
3.7. References ...116
CHAPTER IV: Corn kernel hardness, drying temperature and amylase supplementation affects live performance and nutrient utilization of broilers...146
4.1. Abstract ...146
4.2. Introduction ...148
4.3. Materials and methods ...152
4.4. Results ...157
4.5. Discussion ...160
4.6. Conclusion ...165
4.7. References ...166
CHAPTER V: Corn drying temperature, particle size, and amylase supplementation influences growth performance, digestive tract development, and nutrient utilization of broilers ...189
5.1. Abstract ...189
5.2. Introduction ...191
5.3. Materials and methods ...193
5.4. Results ...200
5.5. Discussion ...204
5.6. Conclusion ...208
CHAPTER VI: Final conclusions ...234
6.1. NIR results ...234
6.2. Particle size ...234
6.3. Live performance and organ development ...235
LIST OF TABLES
Table 2.1 DM, proximate composition, starch and carbohydrate fractions, phytate, mycotoxin content and water activity (Aw) of two un-dried corn hybrids
varying in kernel hardness ...………..………..…. 65
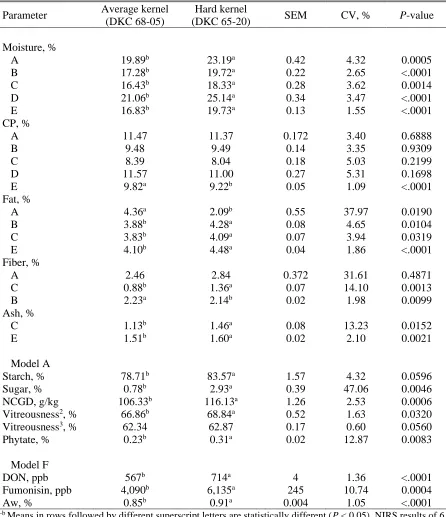
Table 2.2 Kernel hardness and drying temperature effects on moisture and DM content in whole corn samples analyzed with different NIRS calibration models (A-E) ………..…... 66
Table 2.3 Kernel hardness and drying temperature effects on proximate composition in whole corn samples analyzed with different NIRS calibration models (A-F).………..67
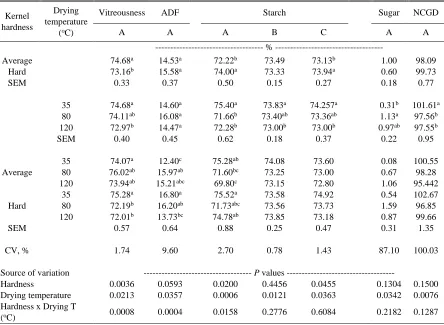
Table 2.4 Kernel hardness and drying temperature effects on vitreousness, starch and starch fractions, and sugar content in whole corn samples analyzed with
different NIRS calibration models (A-F) ……….……….… 68
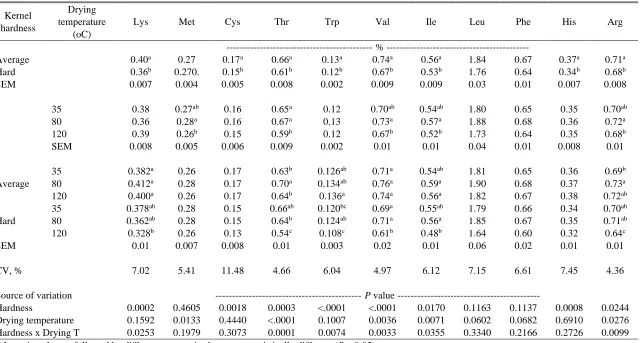
Table 2.5 Kernel hardness and drying temperature effects on total amino acids content in whole corn samples, analyzed with NIRS calibration model D ………..…. 69
Table 2.6 Kernel hardness and drying temperature effects on standardized ileal amino acid digestibility for poultry in whole corn samples, analyzed with NIRS
calibration model D.………...…...70
Table 2.7 Kernel hardness and drying temperature effects on standardized ileal amino acid digestibility for swine in whole corn samples, analyzed with NIRS
calibration model D ...………..………….. 71
Table 2.8 Kernel hardness and drying temperature effects on energy and phosphorous in whole corn samples, analyzed with NIRS calibration model D ……..…...72
Table 2.9 Kernel hardness and drying temperature effects on CP and protein solubility index (PSI) in ground corn samples analyzed with different NIRS calibration models (A-F) ……….……….73
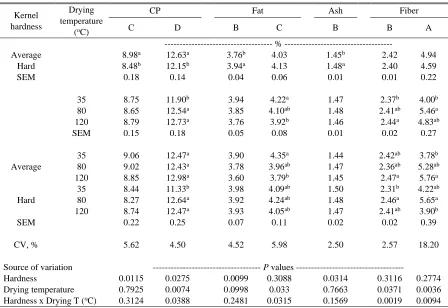
Table 2.10 Kernel hardness and drying temperature effects on proximate composition in ground corn samples analyzed with different NIRS calibration models (A-F) …... 74
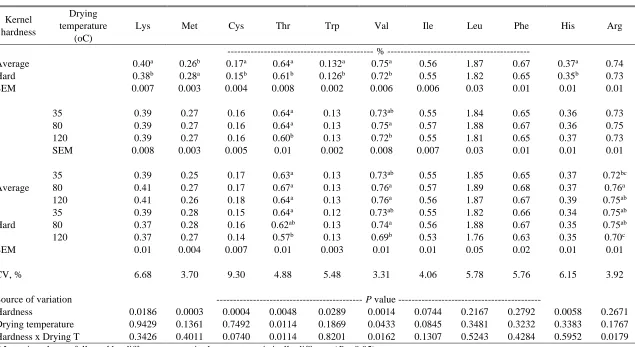
Table 2.12 Kernel hardness and drying temperature effects on total amino acid content for swine in ground corn samples, analyzed with NIRS calibration model D ………. 76
Table 2.13 Kernel hardness and drying temperature effects on standardized ileal amino acids digestibility for poultry in whole corn samples, analyzed with NIRS calibration model D ………..……...….. 77
Table 2.14 Kernel hardness and drying temperature effects on standardized ileal amino acids digestibility for swine in whole corn samples, analyzed with NIRS
calibration model D………..………...78
Table 2.15 Kernel hardness and drying temperature effects on energy, total, phytate, non-phytate, and digestible phosphorous in ground corn samples analyzed with different NIRS calibration models (A-D) ………...……….…..79
Table 2.16 Kernel hardness and drying temperature effects on corn fatty acid profile in ground samples analyzed with NIRS calibration model B ...………..…..….80
Table 2.17 Kernel hardness and drying temperature effects on mycotoxin content, and water activity in ground corn samples analyzed with NIRS calibration model F ………...…...81
Table 2.18 Kernel hardness and drying temperature effects on proximate composition, and protein solubility in whole stored (6mo) grain samples analyzed with
different NIRS calibration models (A-F) ………..……….………....82
Table 2.19 Kernel hardness and drying temperature effects on vitreousness, starch, arabinose (A), xylose (X), and glucose in whole stored (6mo) grain samples analyzed with different NIRS calibration models (A-F) …………...……....….83
Table 2.20 Kernel hardness and drying temperature effects on energy, total, phytate, non-phytate, and digestible phosphorous in whole stored (6mo) grain samples analyzed with different NIRS calibration models (A-F) ………...….84
Table 2.21 Kernel hardness and drying temperature effects on standardized ileal amino acids digestibility for poultry in whole stored (6mo) corn samples, analyzed with NIRS calibration model D ………..………...…….…...85
Table 2.22 Kernel hardness and drying temperature effects on standardized ileal amino acids digestibility for swine in whole stored (6mo) corn samples, analyzed with NIRS calibration model D ………..………...….……...86
Table 2.24 Kernel hardness and drying temperature effects on proximate composition in ground stored (6mo) corn samples analyzed with different NIRS
calibration models (A-F) at 6 months of storage ………...…....88
Table 2.25 Kernel hardness and drying temperature effects on vitreousness, ADF, NDF, starch, NPS, arabinose (A), and xylose (X), in ground stored (6mo) corn samples analyzed with different NIRS calibration models (A-F) ……...89
Table 2.26 Kernel hardness and drying temperature effects on total amino acids content in ground stored (6mo) corn samples, analyzed with NIRS calibration model D ...90
Table 2.27 Kernel hardness and drying temperature effects on standardized ileal amino acid digestibility for poultry in ground stored (6mo) corn samples, analyzed with NIRS calibration model D ……….………....91
Table 2.28 Kernel hardness and drying temperature effects on standardized ileal amino acid digestibility for swine in ground stored (6mo) corn samples, analyzed with NIRS calibration model D ………..………...…92
Table 2.29 Energy and nutrient content comparison of two corn hybrids post-harvest with different drying temperatures ……….………..………..93
Table 2.30 Kernel hardness and drying temperature effects on mycotoxin content, and water activity in whole and ground stored (6mo) samples1 analyzed with NIRS calibration model F ..………..….……….94
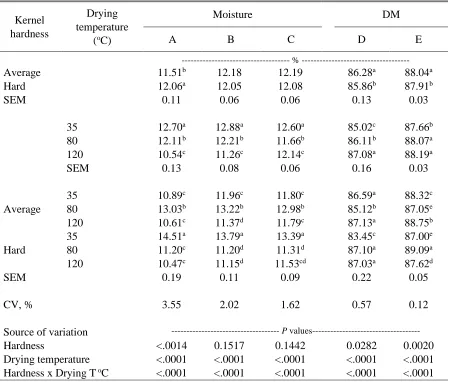
Table 3.1 Effect of corn kernel hardness and drying temperature on the particle size geometric mean (dgw) of fresh corn when grinding in hammermill and roller mill at different settings ………..……….…...127
Table 3.2 Effect of corn kernel hardness and drying temperature on the particle size distribution expressed as standard deviation (Sgw) of fresh corn, when
grinding in hammermill and roller-mill and at different settings ..…………..128
Table 3.3 Effect of corn kernel hardness and drying temperature on the particle size geometric mean (dgw) of 30wk stored corn when grinding in hammermill and roller-mill at different settings ...……….…….….129
Table 3.5 Effect of corn kernel hardness and grain drying temperature on corn particle size geometric mean (dgw), and particle size distribution expressed as standard deviation (Sgw) while grinding in a hammermill1 at 2, 4 and 8 wk of grain storage, respectively ………...………131
Table 3.6 Effect of particle size and grain drying temperature on corn particle size geometric mean (dgw) and distribution expressed as standard deviation (Sgw)while grinding in a hammermill at 12, 17 and 20 wk of grain
storage, respectively ……….………...……….132
Table 3.7 Effect of storage time on particle size geometric mean (dgw) while grinding in a hammermill at different speeds for fresh corn and stored corn at 2, 4, 12, 17, 20, and 30 wk ………..…….………133
Table 3.8 Effect of storage time on particle size distribution expressed as standard deviation (Sgw) while grinding in a hammermill at different speeds for fresh corn and stored corn at 2, 4, 12, 17, 20, and 30 wk ……….…...134
Table 3.9 Effect of storage time on particle size distribution (Dgw) while grinding in a roller mill at different roller gap settings for fresh1 corn and stored corn at 30 wk ……….……….…..135
Table 3.10 Effect of kernel hardness and grain drying temperature on pellet durability index (PDI) of starter 1, grower 1, and finisher 1 broiler diets with corn stored for 2, 4 and 8 wk, respectively ……….………...136
Table 3.11 Effect of particle size and grain drying temperature on pellet durability index (PDI) of starter 2, and grower 2 broiler diets, with corn stored for 12, and 17 wk, respectively ……….………...…...137
Table 3.12 Effects of corn kernel hardness and drying temperature on production rate of feed (lb/30s) and temperature variation (ΔT) between conditioner and hot pellets for starter 1, grower 1, and finisher 1, broiler diets
produced at 2, 4, and 8 wk of grain storage. ……….…………..… 138
Table 3.13 Effects of corn kernel hardness and drying temperature on production rate of feed (lb/30s) and temperature variation (ΔT) between conditioner and hot pellets for starter 2, and grower 2 broiler diets produced at 12, and 17 wk of corn storage …..……….………….…. 139
Table 3.14 Effect of corn kernel hardness and grain drying temperature on
vitreousness and starch content ……….…………...140
Table 4.2 Ingredient composition of starter, grower, finisher and withdrawal basal diets for Ross-708 male broilers ………..………….……….. 178
Table 4.3 Calculated and analyzed nutrient content of basal starter, grower, and
finisher diets for Ross-708 male broilers ….………179
Table 4.4 Declared and analyzed activities of amylase in the experimental diets...…….180
Table 4.5 Effect of corn kernel hardness, grain drying temperature, and enzyme
supplementation on live performance of Ross 708 male broilers at 14 d in floor pens (Experiment 1)……….………... 181
Table 4.6 Effect of corn kernel hardness, grain drying temperature, and enzyme supplementation on live performance of Ross 708 male broilers at 28 d in floor pens (Experiment 1)………..………...………….……...182
Table 4.7 Effect of corn kernel hardness, grain drying temperature, and enzyme
supplementation on live performance of Ross 708 male broilers at 40 d in floor pens (Experiment 1)……….…………..……….…… 183
Table 4.8 Effect of corn kernel hardness, grain drying temperature, and enzyme supplementation on live performance at 14 d of Ross 708 male broilers in battery cages (Experiment 2)………...… 184
Table 4.9 Effect of corn kernel hardness, grain drying temperature, and enzyme supplementation on nutrient digestibility at 16d in battery cages
(Experiment 2)………..185
Table 5.1 Moisture content of corn before and after drying treatments………...220
Table 5.2 Ingredient composition of starter, grower, finisher and withdrawal basal diets for Cobb-500 male broilers..……….………...221
Table 5.3 Calculated and analyzed nutrient content of basal starter, grower, and
finisher diets for Cobb-500 male broilers……….……….………...222
Table 5.4 Particle size geometric mean (Dgw) and distribution (Sgw) for dietary feeds ……….………....223
Table 5.5 Declared and analyzed activities of amylase in the experimental diets….…...224
Table 5.6 Effect of corn drying temperature, particle size and enzyme
Table 5.7 Effect of corn drying temperature, particle size and enzyme
supplementation on live performance of Cobb 500 male broilers at 28 d of age in floor pens (Experiment 1) ………..….…..226
Table 5.8 Effect of corn drying temperature, particle size and enzyme
supplementation on live performance of Cobb 500 male broilers at 40 d of age in floor pens (Experiment 1)…………..………....…227
Table 5.9 Effect of corn drying temperature, particle size and enzyme
supplementation on live performance of Cobb 500 male broilers at 14 d of age in battery cages (Experiment 2) ……….……….……. 228
Table 5.10 Effect of corn drying temperature, particle size and enzyme
supplementation on gastrointestinal organ development of Cobb 500 male broilers at 42 d of age in floor pens (Experiment 1). …………..……...229
Table 5.11 Effect of grain drying temperature, particle size, and enzyme supplementation on nutrient digestibility at 42d in floor pens
(Experiment 1)………..………....230
Table 5.12 Effect of grain drying temperature, particle size, and enzyme supplementation on nutrient digestibility at 16d in battery cages
LIST OF FIGURES
Figure 1.1 Whole corn kernel with the indication of endosperm, germ, bran and tip cap (Ai and Jane, 2016) ………..……….…...3
Figure 1.2 Sequence of events in the digestion of starch in non-ruminants. Starch is normally in granular form and pancreatic alpha-amylase progressively hydrolyse constituent amylose and amylopectin to maltose, maltotriose and alpha-limited dextrins (Moran, 1985)……….……….11
Figure 2.1 Effects of two corn kernel hardness (average and hard), and three drying temperatures (35, 80, and 120°C) on starch content (g/100 g, DM) analyzed by three different NIRS prediction models (colors represent each model) at 2 and 24 wk of grain storage………...95
Figure 2.2 Effects of two corn kernel hardness (average and hard), and three drying temperatures (35, 80, and 120°C) on crude protein content (g/100 g, DM) analyzed by four different NIRS prediction models (colors represent each
model) at 2 and 24 wk of grain storage………...96
Figure 3.1 Lineal regression effect (P < 0.0001) of corn storage time on particle size geometric mean (Dgw) in hard corn kernel when grinding at 1,400 rpm in a hammermill. Particle size, Dgw = 1,077.5292 -
14.679065*wk (R2 = 0.67) ………...……….. 141
Figure 3.2 Lineal regression effect of corn storage time on particle size geometric mean (Dgw) in average (P < 0.05; R2 = 0.23) and hard corn kernel (P < 0.05) when grinding at 3,600 rpm in a hammermill. For average corn kernel: Particle size, Dgw = 443.2 - 0.7908371*wk (R2 = 0.23). For hard corn kernel: Particle size, Dgw = 463.72019 - 1.349858*wk
(R2 = 0.17) ……..……….…….……...142
Figure 3.3 Lineal regression effect of corn storage time on particle size geometric mean (Dgw) in corn dried at 35 (P ≤ 0.001), 80 (P < 0.001), and 120°C (P < 0.001) when grinding at 3,600 rpm in a hammermill. For corn dried at 35°C: Particle size, Dgw = 502.58681 - 2.6391391*wk (R2 = 0.67). For corn dried at 80°C: Particle size, Dgw = 424 - 0.6219908*wk (R2 = 0.95). For corn dried at 120°C: Particle size, Dgw = 298.075 +
8.8007143*wk (R2 = 0.81) ………...………...142
Figure 3.5 Distribution of particle size for two corn hybrids varying in kernel hardness (average and hard) and dried at three temperatures (35, 80, and 120°C), while grinding in a hammermill (12-12 screen combination and 1,600 rpm) at 4 wk post-harvest………...…………...144
Figure 3.6 Distribution of particle size for two corn hybrids varying in kernel hardness (average and hard) and dried at three temperatures (35, 80, and 120°C), while grinding in a hammermill (12-12 screen combination and 1,600 rpm) at 6 wk post-harvest…………...………145
Figure 4.1 Effect of corn kernel hardness and three drying temperature on the proportion of amylose within the starch. Means not sharing a common superscript (a-c) are significantly different (n=3; P < 0.01) by
Tukey’s test ……...……….………..186
Figure 4.2 Effect of corn kernel hardness and drying temperature on starch gelatinization. Means not sharing a common superscript (a-e) are
significantly different (n=6; P < 0.01) by Tukey’s test……….……...186
Figure 4.3 Effect of corn kernel hardness and drying temperature on FCR at 40 d of age. Means not sharing a common superscript (a-c) are significantly
different (n=8; P < 0.01) by Tukey’s test……….…...….187
Figure 4.4 Interaction effect (P < 0.01) of amylose content and kernel hardness on AMEn at 16d in corn with average and hard kernel hardness. For average corn kernel, AMEn = 4665.7633 - 51.868479*amylose content, R2 = 0.25; For hard corn kernel AMEn = 2823.2909 + 27.631234* amylose content,
R2 = 0.28 ……….……….…………187
Figure 4.5 A. Lineal regression effect of corn in vitro starch gelatinization with FCR at 40d (P < 0.01) on average and hard corn kernel hybrids. For average corn kernel, FCR 0-40d = 1.5743792 - 0.0197937*starch gelatinization. For hard corn kernel, FCR 0-40d = 1.5151513 -
0.0082148*starch gelatinization. B. Interaction effect (P < 0.01) of corn amylose content within starch and kernel hardness on FCR at 40d in corn with average and hard kernel. For average corn kernel, FCR 0-40d = 1.5578868 - 0.0024241* amylose content. For hard corn kernel, FCR 0-40d = 1.447081 + 0.0021232* amylose content……….……..188
Figure 5.1 Effect of two grain drying temperatures on the proportion of amylose within the starch. Means not sharing a common superscript (a-b) are
significantly different (n=3; P < 0.01) by Tukey’s test………232
Figure 5.2 Effect of two grain drying temperatures on in vitro starch gelatinization. Means not sharing a common superscript (a-b) are significantly different
Figure 5.3 Effect of two grain drying temperatures on protein solubilization. Means not sharing a common superscript (a-b) are significantly different
CHAPTER I
LITERATURE REVIEW
Introduction
Corn is the most important cereal grain used in the feed industry worldwide, with an annual
production of 1,000 million of tons (Ai and Jane, 2016; IGC, 2018; USDA, 2019). Depending
on the desired agronomic traits that grain producers want, a variety of maize hybrids are
commercially available for planting (Moore et al., 2008). Corn kernel hardness is principally
a genetic expression, but environment and post-harvest handling (e.g. transportation, drying
and storage) also influence hardness properties (Williams et al., 2009). In corn, hardness
measurement is important for breeding in order to retain trait characteristics (Pratt et al., 1995).
After harvest, corn is usually dried either in the field or employing industrial dryers with hot
air prior to storage and further use. Previous studies reported that drying temperatures usually
reach more than 100°C, mainly to reduce time of energy consumption in grain dryers (Larbier
et al., 1971; Barrier-Guillot et al., 1993; Li et al., 2014; Odjo et al., 2015a). However, either
drying at high temperatures and/or exposing the grain to heat for a long time, regardless the
temperature, could result in the production of indigestible complexes, that could impair the
digestibility of nutrients and consequently depressing broiler live performance (Larbier et al.,
1971; Iji et al., 2003; Cowieson, 2005; Malumba et al., 2008; Huart et al., 2018).
This literature review will discuss topics related with the nutritional value of corn for
broiler chickens, and how it is affected by drying temperatures and its interactive effects with
manufacturing process, and the current technologies used by animal nutritionists worldwide
such as Near Infrared Spectroscopy (NIRS). The effects of amylase supplementation will also
be revised as an alternative to ameliorate the possible negative effects of drying temperatures
and its interactions with kernel hardness.
Genetic selection
Agronomic traits such as grain production, disease and insect resistance, drought tolerance
and storage characteristics, have been the primary criteria for selecting corn hybrids (Moore et
al., 2008). While most of the corn maize is used for human consumption in developing
countries, it is mainly used for industrial purposes and animal feed in developed countries
(FAO, 1992; IGC, 2018). Because of its importance to food and energy security, the genetic
improvement of corn has played a key role in the development of genotypes with high
technological and nutritional values (Wang et al., 1993, IGC, 2018).
A selection pressure by both humans and nature has resulted in various maize types,
generally classified by properties of their grain endosperm. The most common types of maize
include flint, flour, dent, popping, sweet and waxy (Knott et al., 1995). Flour maize endosperm
is made of soft starch with thin pericarps. Dent maize with flinty sides and soft cores of starch
that cause the end of the grains to collapse or dent during drying fall between the flint and flour
types (Zilic et al., 2011).Waxy maize is a starch variant of corn which contains 100%
amylopectin, whereas normal maize contains 75% amylopectin and 25% of amylose (Plumier
Maize (Dent corn)
In the U.S. and in the rest of the world, corn hybrids are primarily dent-endosperm cultivars
(Watson 1987; Correa et al., 2002). Maize kernels are quite large relative to those of other
cereals but similar in composition in that the most dominant component is the starchy
endosperm (Sull et al., 1990). The endosperm accounts for more than 80% of the physical
kernel structure. Within the endosperm, individual compartments (cells) contain the starch as
well as storage proteins and lipids (Williams and Kucheryavskiy, 2016). These compartments
have walls, which are made up of non-starch polysaccharides (β-glucan and arabinoxylan),
protein, and phenolic acids (Fox and Manley, 2009). Maize kernels also have a germ, as well
as outer layers (pericarp and aleurone) that surround the endosperm. The tissues in these outer
layers are alive, unlike the endosperm, and provide important metabolic systems for the
protection of the kernel against pathogens and insects (Evers et al., 1999). In addition, these
tissues drive the biochemical processes to degrade the endosperm when the kernel is required
to germinate (Mestres and Matencio, 1996). A whole corn kernel is composed of 4 different
parts: endosperm (82% to 84% of whole kernel mass), germ (10% to 12%), bran (5% to 6%),
and tip cap (1%) on a dry matter basis, as shown in Figure 1 (Ai and Jane, 2016).
The physical shape of maize and the structures of the individual components have been
shown to affect maize hardness (Evers et al., 1999). Maize can be short and round (popcorn)
or long and flat, with an oval, distal end (flint), or short and flat with a dented distal end (dent).
In general, flint and dent types of maize are more common, with dent maize being softer than
flint maize (Pomeranz et al., 1985). However, whereas the physical shape and size of flint or
dent maize affect hardness, the interesting characteristic of maize is that there can be both soft
and hard endosperm within a single kernel (Pomeranz et al., 1984). The endosperm is the main
internal structure and contributes significantly to hardness. As mentioned, starch and protein
are the most abundant components, and both of these components affect hardness (Gayral et
al., 2016; Milašinović-Šeremešić et al., 2019). However, although protein is much lower in
proportion than starch, it plays the major role in final physical hardness (Caballero-Rothar et
al., 2019).
Kernel hardness
The storage protein in cereals is named prolamin, after the high proline and glutamine
content found in these proteins (Salunkhe et al., 1977). In maize, the protein exists in a layered
ball-like structure or body, instead of forming a matrix, linked by covalent or molecular amino
acid bonding. These protein bodies adhere to the starch granule (Zilic et al., 2011). Protein in
maize comprises around 8-12% of the total composition. Protein content has been correlated
to hardness, and the variation in zein classes has been linked to differences in hardness (Bantte
and Prasanna, 2004). Although the protein content comprises a very low proportion of the total
kernel composition, it would appear that it does play a significant role in influencing hardness
capacity of proteins to adhere to starch granule surfaces creating a protein matrix. The glassy
endosperm is tightly compacted with few or no airspaces (Williams and Kucheryavskiy, 2016).
The floury endosperm on the other hand comprises spherical starch granules that are covered
with a protein matrix without zein (maize storage protein) bodies (Evers et al., 1999;
Caballero-Rothar et al., 2019).
Maize can be categorized into five general classes by kernel hardness, more appropriately
termed vitreousness, which means glassy (Watson, 1987;Williams et al., 2009; Kljak et al.,
2011). These classes are in decreasing order of vitreousness: flint, popcorn, flour, dent, and
sweet (Ratnayake and Jackson, 2003). Flint maize has a hard, starchy layer surrounding the
outer part of the kernel and has an endosperm vitreousness of around 65–75%. Endosperm
hardness is important to producers and processors for the trade of corn, because affects yield
during dry-milling, energy consumption, and dust formation during food processing (Mestres
and Matencio, 1996). It has been reported that the nutrient composition of corn varied among
corn hybrids, and that is was associated with hardness of the kernel (Seetharaman et al., 2001;
Zilic et al., 2011). In addition, previous reports concluded that corn kernel hardness affected
the live performance and nutrient digestibility in broiler chickens (O’Neill et al., 2012), and
that some of the responses were attributed to the interactive effects of kernel hardness with
grain drying temperatures (Kaczmarek et al., 2014).
Some analysis to determine maize hardness require destruction of the sample (Pomeranz
et al., 1984, 1985; Williams et al., 2009). However, NIRS has been used for over 30 years to
estimate maize hardness (Fox and Manley, 2009), and nutrients such as protein and lysine for
breeding programs purposes (Rosales et al., 2011). Considering that drying temperatures of
attributed to kernel hardness, then it is of interest to explore if NIRS is able to detect these
variations in nutrient and physical quality.
Vitreousness and corn quality
Vitreousness provide an indication of endosperm texture (Fox and Manley, 2009).
Although not a direct measure of hardness, vitreousness provides a measure of the endosperm
that is translucent (Watson, 2003). Studies on those areas of the endosperm that are translucent
against the nontranslucent areas show differences in protein content and zein fractions (Fox
and Manley, 2009). Previous studies have correlated other hardness measures with
vitreousness with the more vitreous samples being harder than the less vitreous samples
(Mestres and Matencio, 1996; Caballero-Rothar et al., 2019). In addition, studies have
correlated the level of vitreousness to end-use quality. Therefore, using vitreousness as a
quality parameter in corn (Dombrink-Kurtzman and Bietz, 1993; Gehring et al., 2012).
Furthermore, Kljak et al. (2018), observed that amylose (P < 0.005; r = 0.29) content and
amylose:amylopectin ratio (P < 0.005; r = 0.29), positively correlated with vitreousness, which
it has been also observed by Gayral et al. (2015). Higher amylose concentrations were detected
in vitreous than in floury endosperm. Meaning that corn samples with higher proportion of
vitreous endosperm are expected to have higher amylose content in the entire kernel. The
amylose content (P < 0.005; r = 0.84) as well as the amylose:amylopectin ratio (P < 0.005; r
= 0.86) have been reported to be positively correlated to the TME value for ducks (Zhou et al.,
2010). In this way, the vitreousness in corn is an important parameter to consider when
Carbohydrates
Chemically, carbohydrates vary from the low-molecular-weight sugars, such as glucose,
fructose, and sucrose, to some of the largest natural polymers known, amylopectin and
cellulose. Some of these high molecular-weight entities are made up of a single simple sugar;
for example, the amylopectin fraction of starch is made up essentially of glucose. Other
complex carbohydrates consist of numerous sugars and may contain non-carbohydrates, such
as proteins, lipids, and phenolics (Hoseney, 1984).
Starch
Starch is the most abundant carbohydrate in all cereal grains, constituting about 73% of the
dry matter of the dent maize grain (Cowieson, 2005; Lehmann and Robin, 2007). In cereal
grains, starch is stored inside the granules in the endosperm. The granules consist of
semi-crystalline and amorphous layers and exist in various sizes and shapes (Svihus et al., 2005a).
Amylose and amylopectin are the main building components of layers and both are polymers
of glucose units (Singh et al., 2005; Svihus et al., 2005). Amylopectin, the highly branched component of starch, consists of chains of α-D-glucopyranosyl residues linked together mainly
by 1-4 linkages, with 5 to 6% of 1-6 bonds forming branch points (Gray, 1992; Le-Bail et al.,
2017). Amylose is the smaller of the two fractions and possesses very few branches. The levels
of amylose and amylopectin content found in a corn starch sample will depend on the variety,
and these levels are fundamentally governed by genetics (Wang et al., 1993; Pirgozliev et al.,
2010; Lian et al., 2012). In solution, amylose molecules may adopt a helical form and associate
with organic acids, alcohols, or, more importantly, lipids to form complexes in which a
Corn as feedstuff
Corn is the primary source of energy for domestic animal nutrition (Kljak et al., 2018) and
main feed ingredient for poultry (IGC, 2018; USDA, 2019). Corn is relatively free of viscous
non-starch polysaccharides, one of the major anti-nutritional factors present in most other
cereals such as barley, wheat, and oats (Kaczmarek et al., 2014). In a typical broiler diet
formulation, maize can contribute about 65% of broiler metabolizable energy (ME)
requirements (Baurhoo et al., 2011), which suggests that maize quality differences may lead
to dramatic variance in broiler growth performance and feed costs per unit of production. The
AMEn of maize for broilers may vary by more than 400 kcal/kg and as it is affected by
genetics, agronomic conditions, proximate composition, pre- and post-harvest processing
variables, and the presence of anti-nutritional factors such as phytate, resistant starch, enzyme
inhibitors and insoluble non-starch polysaccharides (Cowieson, 2005; Rutherfurd et al., 2007;
Gehring et al., 2013).
Nutritional importance and quality of corn as a feedstuff
Corn protein is mainly stored in both the endosperm and the germ, which are the two
principal parts of maize kernels (Kljak et al., 2011). But there are large differences in the
characteristics of protein present in the two parts. Generally, the germ contains 35% of
high-quality protein, whereas the endosperm has only 9% of poor-high-quality protein (Wall and
Donaldson, 1975; Mestres and Matencio, 1996). Normally, in corn, the endosperm accounts
for 80-85% of the total dry weight of kernels, while the germ constitutes the remaining
The relative amounts of protein are also dependent on the type, texture, size of kernels,
genotypes, and the environments in which the maize is grown (Baye et al., 2006).
According to the solubility in different solvents, maize proteins in kernels are divided into
several fractions. The prolamins, referred to as the zeins, are the major protein fractions (Wall
and Donaldson, 1975). They are soluble in alcohol, comprising 52% of kernel nitrogen. The
glutelins, which account for 25% of kernel nitrogen, are soluble in dilute alkaline solutions,
while the water-soluble albumins comprise 7% of kernel nitrogen. The globulins are soluble
in salt solutions and consist of 5% kernel nitrogen. Maize proteins have a low nutritional value
because they are deficient in essential amino acids such as lysine and tryptophan (Bantte and Prasanna, 2004; Milašinović-Šeremešić et al., 2019a).
Corn quality and its chemical composition changes due to many factors that includes
genetic variety, agronomic and post-harvest practices such as drying method, and storage
conditions (Cromwell et al., 1999; Uribelarrea et al., 2004; Cowieson, 2005). These effects
have been observed even in hybrids from the same variety (Lasek et al., 2012), leading to
nutritional variation between batch to batch while manufacturing feed (Cowieson, 2005).
Starch dynamics of digestion in poultry
Amylase is the major responsible enzyme for starch digestion in mammals. Although, there is some α-amylase activity present in the saliva of humans and pigs (McDonald, 2002), in
poultry, salivary amylase does not supply any important function, thus pancreatic α-amylase
is the only starch degrading enzyme (Moran, 1982; Wiseman, 2006). A previous study (Nir et
al., 1993) reported that the pancreatic amylase activity was suboptimal in young chickens as
hatchlings is still immature (Uni et al., 1998). Findings by these researchers suggested that
younger chicks may have limitations to utilize gelatinized starch. Although all starch
potentially can be digested by alpha-amylase and the brush border enzymes in the small
intestine (Figure 2), a certain fraction of starch will resist digestion in the small intestine either
because it is trapped within whole plant cells and food matrices (resistant starch, RS1), because
of the crystalline nature of the starch granule (RS2), because the starch is retrograded (RS3) or
because the starch is chemically modified (RS4) (Englyst et al., 1992). This could be
exacerbated by high drying temperatures in corn as it will form resistant starch as observed by
Kaczmarek et al. (2014), thus impacting starch digestibility in broilers.
The digestive rate of starch generally decreases with increased amylose content and
branch- chain length of amylopectin (Jane et al., 2003; Li et al., 2008). Other factors, such as
surrounding by cellulose and hemi-cellulose fibers and/or protein matrices and interacting with
lipids, can also reduce the susceptibility of starch to enzyme hydrolysis (Ezeogu et al., 2005;
Ezeogu et al., 2008; Ai et al., 2013). Zhou et al. (2010) observed (P < 0.05) a strong positive correlation (r = 0.86),between amylopectin:amylose ratio and the TME value when conducting
an experiment in ducks. Ducks fed diets containing maize with high amylose content, tended
to have lower TME than the ones fed treatments with low amylose corn. Similarly, Pirgozliev
et al. (2010) reported that diets containing high amylose, negatively affected the live
performance in broiler chickens. Therefore, it is important to determine if
Figure 1.2. Sequence of events in the digestion of starch in non-ruminants. Starch is normally in granular form and pancreatic alpha-amylase progressively hydrolyse constituent amylose and amylopectin to maltose, maltotriose and alpha-limited dextrins (Moran, 1985).
In broilers, the rate of starch digestion varies among feedstuffs (Weurding et al., 2001).
Weurding et al. (2001) determined that the rates of starch digestion were different between
hammer-milled and roller-milled corn although the extent of digestion was equal (97%). This
meaning that particle size will influence the starch digestion in broilers.
Processing of corn in feed manufacturing Drying process
Before corn could be stored for later use as a feed ingredient, it is dried to reach moisture
between 12 and 15%. This is achieved by different means and technologies worldwide. (Odjo
et al., 2015). The drying process can occur in the field, but it is commonly conducted using
industrial grain dryers with high temperatures that can reach 50oC in crossflow dryers and up
per tonne of grain (Larbier et al., 1971; Kaczmarek et al., 2014a; Li et al., 2014). Elevated
temperatures are used to achieve maximum grain drying energy efficiency and reduce costs
(Barrier-Guillot et al., 1993). Nevertheless, high kernel temperatures may induce the formation
of new chemical bonds within and between substrates that are resistant to digestive enzymes
(Larbier et al., 1971; Iji et al., 2003; Bhuiyan et al., 2010a). In addition, when maize is
harvested with high moisture content and then dried, the nutritional value may be
compromised, with a concomitant effect on dietary AME, BW gain, and FCR (Barrier-Guillot
et al., 1993; Iji et al., 2003; Huart et al., 2018). These adverse effects are associated with
denaturation of heat-labile vitamins, damage to protein via an interaction with reducing sugars,
and retrograde starch formation after heat processing (Malumba et al., 2008; Adeola and
Cowieson, 2011).
Starch gelatinization is a phenomenon manifested as several changes in properties,
including granule swelling and progressive loss of organized structure (detected as loss of
birefringence and crystallinity), increased permeability to water and dissolved substances,
increased leaching of starch components, increased viscosity of the aqueous suspension, and
increased susceptibility to enzymatic digestion (Setiawan et al., 2010). When starch is heated
with excess water, it transitions from an ordered to a disordered structure, what is called
gelatinization. (Rosicka-Kaczmarek et al., 2017; Semeijn and Buwalda, 2017). The term
gelatinization is often used to describe all changes that occur when we heat starch and water,
however, it should be restricted to the loss of birefringence. Once the starch has lost its
birefringence, continued heating to higher temperature with water brings about a continuum of
Past research has associated dietary gelatinized starch both positively and negatively with
pellet quality and broiler performance (Moritz et al., 2001, 2002). However, it has been
speculated that gelatinized starch per se may affect broiler performance aside from its
contribution to pellet binding. Gelatinized starch has been thought to improve enzymatic access
to glucosidic linkages and consequent digestibility (Moran et al., 1989; Colonna et al., 1992).
However, overdoing pelleting may not be positive. Allred et al. (1957) investigated the effects
of reprocessing pelleted feeds. This early study reported a significant improvement in BW gain
and FCR in chicks fed diets that incorporated corn completely processed twice in a pellet
machine. Later research (Sloan et al., 1971; Naber and Touchburn, 1969) evaluating
reprocessed corn-based diets concluded that there was no nutritional benefit to broilers despite
the increased starch gelatinization observed. Moreover, Plavnik et al. (1997) found that feeding
broilers reprocessed corn-based diets resulted in decreased bird performance compared to
broilers fed similar pelleted diets.
Drying temperatures post-harvest cause morphological and structural changes in corn
endosperm due to protein and starch interactions. Variability in the conformation of these
nutrients have been reported to alter enzymatic in vitro digestibility and fermentation of starch
residues (Malumba et al., 2008), impacting nutrient digestibility and live performance (Martins
et al., 2001; Iji et al., 2003; Cowieson, 2005; Huart et al., 2018). In addition, the digestibility
of some essential amino acids specially lysine (Wall and Donaldson, 1975; Rutherfurd et al.,
1997; Odjo et al., 2015), cysteine, tyrosine, and threonine (Kaczmarek et al., 2014a) could be
negatively affected by the production of Maillard reactions (Žilić et al., 2013). In feedstuffs
that have undergone thermal processing or prolonged storage, the free ε-amino group of lysine
proteins, to become nutritionally unavailable. Lysine is converted via Schiff bases and the
Amadori rearrangement to the biologically unavailable deoxyketosyl compound. Whatever the
source of the carbonyl group, the basic reaction rendering the lysine unavailable as a
biochemical component is the same. The portion of undamaged lysine that is thought to be
available for digestion and absorption processes has been defined as reactive lysine
(Torbatinejad et al., 2005; Moughan, 2005). A previous study (Rutherfurd and Moughan,
2007) detected reductions between 27.3 to 36.8% of Lys availability in dried corn. On the other
hand, Kaczmarek et al. (2014) suggested that endosperm hardness could play an important role
in the physical properties that impact protein digestibility. Corn may fracture differently during
grinding depending on kernel hardness, influencing the gizzard function, feed rate passage,
pH, enzymatic activity, and uptake of nutrients in the intestinal lumen.
As drying temperature could possibly modify the endosperm characteristics of corn,
therefore is also of interest to investigate the implications while manufacturing feed, as corn
may fracture different. Particle size has not only important effects in the gastrointestinal tract
of broiler chickens, but it could also impact the effectivity of the exogenous amylase.
Particle size reduction
Both the hammermill and roller mill grinders have traditionally been used to reduce the
particle size of feed grains (Moughan and Hendriks, 2018). Hammermill grinding has typically
created a wider particle size distribution, while roller mill grinding has created a more compact
or narrow particle size distribution (Nir et al., 1995). Although grinding represents a
considerable cost in terms of energy consumption and feedmill capacity, cereals used for
in size. One major reason for this fine grinding is to assure a good pellet quality (Amerah et
al., 2007; Vukmirović et al., 2017a). Fine grinding of cereals has also been considered to be
beneficial for a proper utilization of the feed, but this is a view not supported by most scientific
literature (Svihus et al., 2004).On the other hand, grain particle size is an important factor as
it could influence several aspects of broiler gastrointestinal functionality and physiology (Celi
et al., 2017). Some of these aspects includes nutrient utilization (Benedetti et al., 2011;
Kheravii et al., 2018), growth performance (Nir and Hillel, 1995; Amerah et al., 2007; Zang
et al., 2009; Xu et al., 2015), feed passage rate (Hetland et al., 2002; Svihus et al., 2002),
digestive tract development (Zang et al., 2009; Pacheco et al., 2014), and microbiota profile
(Amerah et al., 2009).
Poultry have been reported to have a preference for larger feed particles (Schiffman, 1968)
and to distinguish the differences in feed particle size by mechanical sensors located in the
beak (Gentle, 1979), which has been observed at all ages (Portella et al., 1988). Traditionally,
fine particles have been associated with larger relative surface area, resulting in greater enzyme
activity in the gastrointestinal tract of poultry (Hetland et al., 2002; Amerah et al., 2007). In
contrast, coarse particles are known to enhance GIT motility, stimulate gizzard function and
reduce pathogen population in the lower intestine, resulting in better overall digestive
efficiency (Pacheco et al., 2013; Xu et al., 2015). The most notably functional contribution of
dietary coarse particles is to improve gizzard development in the chicken, which has been
referred as the pacemaker of gut motility (Duke et al., 1977; Ferket and Gernat, 2006). Feeding
large corn particles has been reported to increase gizzard relative weight (Oliver and Jonker,
1997; Dahlke et al. 2003; Zang et al., 2009; Pacheco et al., 2013, 2014). The gizzard was
improved digestive efficiency. (Duke et al., 1977; Ferket and Gernat, 2006). A well- developed
gizzard reduced digesta pH and passage rate (Nir et al., 1994), enhanced enzymatic digestion
efficiency, and improved energy utilization and nutrient digestibility (Duke, 1992; Amerah et
al., 2007). Therefore, it becomes of interest to explore the interactive effects of corn kernel
hardness, drying temperature and particle size on the gastrointestinal tract development in
broilers.
Agglomeration - pelleting
Some studies have shown improved growth performance with the use of coarsely ground
cereals when the feed is offered in mash form (Reece et al., 1986; Nir, 1994). Birds prefer
more coarse particles and will eat more of a coarsely ground than of a finely ground mash diet
(Reece et al., 1985; Portella et al., 1988; Nir and Hillel, 1994). It is thus necessary to use
pelleted diets to separate the effect of feed structure on feed intake stimulation and the effect
of feed structure on feed utilization. Experiments conducted with pelleted diets containing
cereals ground to different sizes usually shows that a coarse grinding gives similar BW gain
and feed utilization as fine grinding (Reece et al., 1985, 1986; Nir and Hillel, 1995).
Additionally, including coarse ground corn in the feed did not compromise the growth
performance of broilers, even though pellet quality was reduced (Dozier et al., 2010). The
consensus is that pelleting increases performance due to a higher feed intake and increased
feed efficiency. It is not clear, however, whether the increased feed per gain is caused solely
by reduced maintenance requirements, or whether pelleting in itself may improve digestibility
decreases digestibility of starch in comparison to mash feeding, despite an improvement in
feed per gain (Svihus and Hetland, 2001).
Many factors can affect pellet quality in a feed mill, but particle size plays an important
role in pellet press operation (Reece et al., 1986; Amerah et al., 2009; Chewning et al., 2012).
Particles larger than 1,000 microns or 1,500 microns may cause fracture of pellets (Svihus et
al., 2004; Amerah et al., 2007; Dozier et al., 2010), while grinding coarser ingredients will
reduce the PDI of the pellet (Muramatsu et al., 2013). Considering that corn dried at different
temperatures will facture differently during the grinding process depending on kernel hardness,
the evaluation of particle size and its implications on pellet quality becomes of interest.
It is very likely that drying temperatures will have an effect on the nutritional value of corn,
and the responses will be dictated by interaction effects with kernel hardness. Therefore, it is
important to evaluate the nutritional composition of corn after drying at different temperatures
using NIRS in both hybrids.
Near-infrared spectroscopy (NIRS) in the feed industry
The NIR region extends from 800 to 2500 nm, between the visible from 380 to 780 nm
(Graham et al., 2013). The greatest impact on the progression of NIR technology was,
however, in the early 1970s, when Phil Williams started using NIR spectroscopy to measure
protein and moisture contents as a basis for trading wheat (Manley, 2014). It is well known
that strict analytical controls and quality assessment have become of crucial importance in the
animal feed sector, not only due to the extensive legislation concerning the manufacturing and
circulation of compound feedstuffs, but also because of the increasing desire of manufacturers
though, chemical analysis procedures are the most widely accepted reference methods for
determining nutritional composition in grains, these methods frequently are destructive and
require time and large samples of grain are needed to perform different analysis (Baye et al.,
2006). NIRS is now considered equally significant among other major analytical technologies
(Graham et al., 2013).
NIRS is a cost efficient and fast indirect method that has emerged as a popular tool used in
different industries. The most noticeable commercial application of NIRS remains in the fields
of agricultural raw materials, food and feed ingredients and finished food products worldwide
(Manley, 2014; Williams and Kucheryavskiy, 2016). There exist numerous applications of
NIRS to feed compound analysis. Most of them address the use of this technology for
prediction of analytical constituents such as moisture, CP (Delwiche, 1998), crude fiber and
crude fat (Perez-Marin et al., 2004, Xiccato et al., 2003, White and Rouvinen-Watt 2004).
Others deal with more complex parameters such as in vivo digestibility and energy value
(Landau et al., 2006). Vitreousness assessment in corn has been also analyzed by NIRS
technology (Armstrong et al., 2007). As suggested by other authors (Williams and
Kucheryavskiy, 2016) it is important to assess the hardness of whole kernels rapidly to be able
to classify them among different types. Although the effectiveness of the NIRS method
depends to a great extent on the accuracy and precision of the reference method, NIRS
measurements and predictions are considered more reproducible than the ones among wet
chemistry labs (Manley, 2014). Consequently, it becomes of interest to investigate if prediction
models develop in ground samples could be applied to efficiently assess the vitreousness
content in whole corn grain. High drying temperatures could have a negative effect on corn
and digestibility of starch. Therefore, the supplementation with an exogenous amylase was
proposed.
Use of exogenous carbohydrases
Exogenous enzymes are now extensively used throughout the world as feed additives in
non-ruminant diets (Adeola and Cowieson, 2011; Khajali and Slominski, 2012; Horvatovic et
al., 2015). The chemical effects of these enzymes are well understood, but the manner in which
their benefits to the animal are brought about is still under debate. Studies with supplemental
enzymes targeting the degradation of substrates that release energy for poultry have been
increasing (Romero et al., 2014; Vieira et al., 2015). Exogenous carbohydrases, such as
xylanases, amylases, and glucanases, have been reported to improve energy utilization and the
performance of broilers (Olukosi et al., 2008; Kiarie et al., 2013; Stefanello et al., 2015a).
Variation in animal performance from flock to flock is also reduced (Engberg et al., 2004;
Khajali and Slominski, 2012).
Carbohydrases and proteases facilitate the action of endogenous enzymes to plant-cell
contents by degrading soluble and insoluble arabinoxylans (Cowieson, 2005; Adeola and
Cowieson, 2011), as well as augmenting endogenous amylase in young animals (Ritz et al.,
1995; Gracia et al., 2003). The combination of these enzymes could reduce the endogenous
losses of amino acids by altering the production of pancreatic enzymes and mucin secretion
which has been commonly associated (Jiang et al., 2008; Cowieson and Bedford, 2009) with
the anti-nutritional properties of non-starch polysaccharides (NSP). Likewise, these enzymes
capturing around 30% of the undigested starch, amino acids, fat and Ca and around 60% of the
undigested P (Cowieson, 2010).
The benefits observed in wheat-based diets for poultry have been widely reported
(Ruangpanit et al., Engberg et al., 2004; Wiseman, 2006). However, the usefulness of
carbohydrases in corn-soybean meal based diets for poultry is variable (Bedford, 2000; Kiarie
et al., 2013), and even more so when phytase is now ubiquitous in the feed (Klein, 2013).
Phytase per se may reduce the main negative component of fiber which is the phytate. Though
there are many interacting factors that affect the measured response to an exogenous enzyme,
the most influential is the nutritional value of the diet to which it is added (Cowieson, 2010).
Despite the high nutritional level of maize, data has been published confirming the
effectiveness of exogenous amylase to improve nutrient digestibility and live performance in
broiler chickens when feeding corn-based diets (Gracia et al., 2003). However, other
researchers failed to prove the beneficial effects previously observed (Kaczmarek et al., 2014a;
Stefanello et al., 2015b). Inconsistencies in the responses while including amylase in the diet
could be attributed to intrinsic effects of corn such as kernel hardness and the processing of
maize while manufacturing feed. Factors such as particle size after grinding are not reported
and they may explain the lack of consistency in results while supplementing amylase. For
example, corn may break different during milling depending on its post-harvest drying
temperature, resulting in different particle size distribution as it has been proposed in an early
study (Kaczmarek et al., 2009). The possible interaction effects among post-harvest drying
temperature, corn kernel hardness particle size post-grinding and its variability, and exogenous
Based on the literature presented here, there is enough evidence to suggest that the
nutritional value of corn will be influenced by the interactions effects of grain drying
temperature with corn kernel hardness. In addition, the effectiveness of an exogenous amylase
to improve live performance, nutrient digestibility and energy utilization could dependent on
the nutritional composition of corn attributed to kernel hardness and drying temperature. Thus,
the objective of the present project was to evaluate the interaction effects of corn kernel
hardness, drying temperature, particle size post-grinding, and amylase supplementation on
broiler live performance, digestive tract and organ development, nutrient digestibility and
energy utilization in broiler chickens. The results of the experiments conducted herein will be
presented in four chapters, including nutrient variability in corn, feed manufacturing, broiler