Basic and Clinical
275
July, August 2018, Volume 9, Number 4
Research Paper: Assessing the Effects of Opioids on
Pathological Memory by a Computational Model
Mehdi Borjkhani1 , Fariba Bahrami1* , Mahyar Janahmadi2,3
1. Motor Control and Computational Neuroscience Laboratory, School of Electrical & Computer Engineering, College of Engineering, University of Tehran, Tehran, Iran.
2. Neuroscience Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran.
3. Department of Physiology, School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran.
* Corresponding Author:
Fariba Bahrami, PhD
Address: Motor Control and Computational Neuroscience Laboratory, School of Electrical & Computer Engineering, College of Engineering, University of Tehran, Tehran, Iran.
Tel: +98 (21) 61114924
E-mail: [email protected]
Introduction:Opioids hijack learning and memory formation mechanisms of brain and induce
a pathological memory in the hippocampus. This effect is mainly mediated by modifications in glutamatergic system. Speaking more precisely, Opioids presence in a synapse inhibits
blockage of N-Methyl-D-Aspartate Receptor (NMDAR) by Mg2+ , enhances conductance of NMDAR and thus, induces false Long-Term Potentiation (LTP).
Methods: Based on experimental observations of different researchers, we developed a mathematical model for a pyramidal neuron of the hippocampus to study this false LTP. The model contains a spine of the pyramidal neuron with NMDAR, α-Amino-3-hydroxy-5-Methyl-4-isoxazole Propionic Acid Receptors (AMPARs), and Voltage-Gated Calcium Channels (VGCCs). The model also describes Calmodulin-dependent protein Kinase II (CaMKII) and AMPAR phosphorylation processes which are assumed to be the indicators of LTP induction
in the synapse.
Results:Simulation results indicate that the effect of inhibition of blockage of NMDARs by Mg2+ on the false LTP is not as crucial as the effect of NMDAR’s conductance modification by opioids. We also observed that activation of VGCCs has a dominant role in inducing pathological LTP.
Conclusion:Our results confirm that preventing this pathological LTP is possible by three different mechanisms: 1. By decreasing NMDAR’s conductance; and 2. By attenuating VGCC’s mediated current; and 3. By enhancing glutamate clearance rate from the synapse.
Article info:
Received: 07 June 2017 First Revision: 10 July 2017 Accepted: 04 October 2017 Available Online: 01 July 2018
Keywords:
Opioids, Memory of addiction, Synaptic plasticity, Long-Term Potentiation (LTP),
Hippocampus
Citation:Borjkhani, M., Bahrami, F., & Janahmadi, M. (2018). Assessing the Effects of Opioids on Pathological Memory by a Computational Model. Basic and Clinical Neuroscience, 9(4), 275-288. http://dx.doi.org/10.32598/bcn.9.4.275
:
: http://dx.doi.org/10.32598/bcn.9.4.275 Use your device to scan
and read the article online
Funding:See Page 284
276 Borjkhani, M., et al. (2018). Assessing the Effects of Opioids on Pathological Memory by a Computational Model. BCN, 9(4), 275-288.
1. Introduction
pioid consumption like other drugs can lead to behavioral changes (Contet, Kief -fer, & Befort, 2004;Williams, Christie, &
Manzoni, 2001). Some researchers believe
that these changes are due to pathological memory formation which can be called memory of ad-diction (Dong & Nestler, 2014;Kelley, 2004; Nestler,
2001). Hippocampus seems to be one of the targets
subject to the plastic changes due to drug abuse (Bao et
al., 2007;Borjkhani, Mahdavi, & Bahrami, 2014;Cap
-ogna, Gähwiler, & Thompson, 1993;Caudle & Chavkin,
1990;Ga, 1980;Heidari et al., 2013;Hosseinmardi, Az
-imi, Fathollahi, Javan, & Naghdi, 2011;Hosseinmardi, Fathollahi, Naghdi, & Javan, 2009).
At the synaptic level, drug-induced changes might be due to Long-Term Potentiation (LTP)/ Long-Term Depression (LTD) induction. Therefore, analyzing LTP/ LTD induction in synapses under the presence of opioids can help researchers to unveil the formation mechanisms of this pathological memory by drugs (Kauer & Malen -ka, 2007;Lüscher & Malenka, 2011).
Opioids affect hippocampal neurons mainly by enhanc-ing glutamate release in presynaptic neurons (Akaishi, Saito, Ito, Ishige, & Ikegaya, 2000;Williams et al., 2001)
and increasing postsynaptic receptors activities (Kauer & Malenka, 2007;van Huijstee & Mansvelder, 2015;
Williams et al., 2001). All the modifications caused by
opioids result in LTP induction in affected synapses (Bao et al., 2007;Wolf, 2003). It has been shown that one of the causes of LTP induction is Calmodulin-dependent protein Kinase II (CaMKII) phosphorylation process
(Lisman, Yasuda, & Raghavachari, 2012;Sanhueza &
Lisman, 2013).
On the other hand, Ca2+ concentration is the main me-diator in this process. Elevation of Ca2+ concentration in postsynaptic neuron triggers CaMKII phosphoryla-tion process. Ca2+ can enter into the neuron through Voltage-Gated Calcium Channels (VGCCs), N-Methyl-D-Aspartate Receptor (NMDAR) and Ca2+ permeable α-Amino-3-hydroxy-5-methyl-4-isoxazole Propionic Acid Receptors (AMPARs). Thus, these channels and receptors contribute in CaMKII phosphorylation process and LTP induction.
It has been shown that blocking hippocampal VGCCs can prevent opioids dependency (Mishra, Barik, & Ray,
2017). Furthermore, opioid effects on NMDARs and
AMPARs were verified in different research studies (Ca -pogna et al., 1993;Chen & Huang, 1992;Chen & Marine,
1991; Ga, 1980; Kauer & Malenka, 2007;Nestler, 2013;
Peters & De Vries, 2012;Williams et al., 2001). Most of the research and findings in this field are based on ex-perimental observations.
However, computational models offer new tools and approaches for investigating neurophysiological ba-sis of addiction memory formation process. In general,
Highlights
•
Opioids form pathological memories by induction of false LTP.•
Decrease in NMDAR conductance and attenuation of VGCC can prevent pathological LTP.•
Enhancing the glutamate clearance rate from the synapse can prevent false LTP.Plain Language Summary
Drug abuse creates pathological memories in different brain regions like the hippocampus. Information about the context of drug abuse is stored in hippocampus. In withdrawal times, the recall of pathological memories can trigger the relapse. Preventing the relapse is one of the challenges in the field of addiction research. Some researchers believe that if the pathological memory is not formed or weakened, the relapse will be undermined. To get a better understand-ing about the latter, we developed a computational model to simulate different cases. Our simulation results suggest that decreasing the conductance of glutamatergic receptors such as NMDARs or clearing the extra glutamates from the synapse, in other words, changing some of the synaptic characteristics can prevent formation of drug-related memories at the synaptic level. This will decrease the possibility of relapse during withdrawal period.
Basic and Clinical
277
July, August 2018, Volume 9, Number 4
Borjkhani, M., et al. (2018). Assessing the Effects of Opioids on Pathological Memory by a Computational Model. BCN, 9(4), 275-288.
computational modeling of biological systems helps us to study consistency of different experimental findings and to predict the behavior of a given biological system under different constraints. That is why computational modeling has recently attracted the interests of many researchers in the field of neuroscience (Amiri, Bahra -mi, & Janahmadi, 2012;Amiri, Montaseri, & Bahrami, 2011; Li et al., 2016;Tewari & Majumdar, 2012b;Vol -man, Bazhenov, & Sejnowski, 2012).
Therefore, in this paper, using experimental findings we introduce a minimal computational model to assess opioid-induced LTP in CA1 region of the hippocampus. This model can also be used for analyzing the effects of other drugs by minor modifications. In the first part of the paper, opioids effect on hippocampal neurons will be discussed. The second part is dedicated to introduce mathematical modeling approach. The third part pres-ents simulation results. Discussion and conclusion will be presented in the final part.
2. Methods
2.1 Neurophysiological background
According to many research studies, opioid-induced modifications in glutamatergic synapses are responsible for pathological memory formation related to the addic-tion (Harris, Wimmer, Byrne, & Aston-Jones, 2004;
Her-man et al., 2003;Mameli, Bellone, Brown, & Lüscher,
2011;van Huijstee & Mansvelder, 2015). Since
Long-Term Potentiation (LTP)/ Long-Long-Term Depression (LTD) induction can be considered as signs of memory forma-tion process at the synaptic level, one can also suggest that addiction memory formation might occur through the induction of LTP/LTD.
It has been shown that opioids induce LTP in hippo-campal synapses by two primary mechanisms: 1. En-hancing glutamate release through disinhibitory mecha-nism (Capogna et al., 1993;Caudle & Chavkin, 1990;
Cohen, Doze, & Madison, 1992;Rezai, Kieffer, Roux,
& Massotte, 2013); and 2. By manipulating postsyn-aptic NMDARs current (Chen & Huang, 1992; Chen & Marine, 1991; Kow, Commons, Ogawa, & Pfaff,
2002; van Huijstee & Mansvelder, 2015). Opioids
modify NMDARs current by increasing conduction of NMDARs and inhibiting Mg2+ from blocking NMDA receptors (Chen & Huang, 1992;Chen & Marine, 1991; Herman et al., 2003).
Consequently, these changes in synaptic transmis-sion result in more calcium flux into the postsynaptic
neuron (Kow et al., 2002;Przewlocki et al., 1999). En-hancement of postsynaptic calcium can activate CAM-KII mechanisms (Herman et al., 2003; Trujillo, 2002). Phosphorylation of CaMKII leads to LTP induction and can be seen as molecular memory formation (Kauer &
Malenka, 2007; Lisman et al., 2012;Sanhueza & Lis
-man, 2013). Furthermore, enhancement of postsynaptic
calcium can lead to phosphorylation of AMPARs (Cas -tellani, Quinlan, Bersani, Cooper, & Shouval, 2005; Castellani, Quinlan, Cooper, & Shouval, 2001;Kauer &
Malenka, 2007).
Also, it has been shown that VGCCs are involved in opioids dependency (Bongianni, Carla, Moroni, &
Pellegrini‐Giampietro, 1986; Mishra et al., 2017). To
investigate the mechanism of a false LTP induced by opioids, we developed a computational model of a postsynaptic neuron in CA1 region. In this model, we considered all important aspects mentioned from these neurophysiological studies. This model will be de-scribed in the next part (Figure 1).
2.2. Computational modeling approach
To analysis opioid-induced synaptic plasticity in CA1 region of the hippocampus, the spine of a CA1 pyrami-dal neuron is modeled by Equation (1):
(1) post post (Vpost V ) Rpostrest m syn
dV
I dt
τ
= − − +This Equation denotes a minimal model to produce Ex-citatory Postsynaptic Potentials (EPSPs) based on Tso-dyks and Markram’s passive membrane model (Tsodyks & Markram, 1997), where τpost shows time constant of neuron membrane, Vrest
post denotes neuron’s membrane potential at rest, and Rm denotes actual resistance of spin. The synaptic current denoted by Isyn can be described by:
(2)
I
syn= −
(I
AMPA+
I
NMDA)
Here, IAMPA and INMDA are AMPAR and NMDAR cur
-rents, respectively. Model of AMPAR current is based on Destexhe’s model (Destexhe, Mainen, & Sejnowski, 1998):
(3)
I
AMPA=
g
AMPAm
AMPA(V
post−
V
AMPA)
Where VAMPA refers to reversal potential of the recep-tor, Vpostis the membrane potential, and mAMPA denotes gating variable of AMPAR. Gating variable of AMPAR can be represented by the following Equation (Destexhe, Mainen, & Sejnowski, 1998):
(4) AMPA g (1 m )
AMPA pre AMPA AMPA AMPA
dm m
278
Here opening and closing rate of the receptor are shown by αampaand βampa, respectively, gpre shows glu-tamate concentration, gAMPA described in Equation (3) denotes AMPAR’s channel conductance that can be enhanced due to CaMKII phosphorylation process by using the following Equation:
(5) 0(1 (CaMKII P1 )/k )
1 half half
AMPA AMPA
g g
e− −
= +
+
Where gAMPA0=0.4 nS is the initial value for AMPAR’s conductance, Phalf=40 µM and khalf=0.4 µM are constants which enables AMPAR conductance to increase through CaMKII phosphorylation process. Model of the NMDAR current can described by the following Equation (Destex -he, Mainen, & Sejnowski, 1998;Moradi et al., 2013):
(6)
I
NMDA=
g
NMDAm
NMDAMg
(V
post−
V
NMDA)
Where Mg shows Mg2+ blocking, represented by:
(7) 2 1 1 1
0 0 1.2 1.2
15.58 0.1
1/ (1 [Mg ] (k 0.1 ) exp( z( 0.1 ) FV R T ))
1 ( ) 1 ( ) post
Mg Op Op δ + − − − = + + − + + +
2 1 1 1
0 0 1.2 1.2
15.58 0.1
1/ (1 [Mg ] (k 0.1 ) exp( z( 0.1 ) FV R T ))
1 ( ) 1 ( ) post
Mg Op Op δ + − − − = + + − + + +
Moreover, NMDAR gating variable is described by:
(8) NMDA �g �1
(
)
�NMDA pre NMDA NMDA NMDA
dm m m
dt =α − −β
Where VNMDA shows the reversal potential of NMDAR, opening and closing rate of receptor are shown by αNMDAand βNMDA respectively, Op refers to opioids con-centration in the synapse, and gpre shows glutamate con-centration. NMDA channels conductance (gNMDA) is rep-resented by the following Equation:
(9)
1.2
0.15
( 0.1 )
1 ( )
NMDA VI VD
g g g
Op
= + +
+
, where gVI and gVD are voltage-independent and volt-age-dependent conductance. The latter conductance is described by:
(10)
,
, 0
(g g ) /
g (V V )
VD
VD VD g
VD post dg dt k
τ
∞ ∞ = − = −Here, gVD,∞ is the final value of gVD, τg denotes time con-stant, gVD,∞ , and Vpost has a linear relation with constant of K. Other parameters and their values have been listed in the Appendix.
Calcium can enter into the neuron through AMPARs, NMDARs, and VGCCs. Also, calcium pumps are
re-sponsible for transferring the calcium out of the neu-ron. So, calcium concentration inside the neuron can be represented by the following Equation (Tewari &
Majumdar, 2012b:
(11)
( )
(
AMPA NMDA R)
post pump
ca spine
I I I
f c s
z FV η +γ + = − − (c ) 1 post post dc f
dt = +θ
(
)
2t endo endo post b K K c θ= +
Here, Cpost shows postsynaptic calcium concentration, IR denotes voltage-gated calcium channels activation, and Spump refers to pumped calcium. Here, η=0.012 and
y=0.06 demonstrate the amount of calcium that can enter into the neuron through AMPARs and NMDARs. Zca=2 denotes calcium valence, F=96487C/mo l is the faraday’s constant, and Vspine=0.9048µM3 is the volume for dendrite
spin. Furthermore, bt=200µM shows total endogenous buf-fer concentration and Kendo=10µM denotes endogenous buffer calcium affinity. Pumped calcium is represented by the following Equation (Tewari & Majumdar, 2012b):
(12)
(
rest)
pump s post post
s
=
k c
−
c
Here, Ks=100/s is the maximum efflux rate of calcium pump, Crest
post=100nM is the rest value for postsynaptic cal -cium concentration, and VGCCs activation is represented by the following Equation (Tewari & Majumdar, 2012b):
(13)
I
R=
g B
R(N, P )(V
open post−
V )
R, where gR=15ps is the conductance of calcium channel, B(N, Popen) denotes a random variable with a binomial dis-tribution which shows the number of opened channels, and VR=27.4mv denotes the reversal potential.
Phosphorylation of CaMKII due to calcium concen-tration can be described by the following Equation
(Zhabotinsky, 2000):
(14)
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
Basic and Clinical
279
July, August 2018, Volume 9, Number 4
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
0 1 2
1 3 1 1 3 1 2 1 3 2 2 1 3 2 2 2 3 3
3 4
2 2 3 3 2 3 3 4 2 3 3 4 2 4 3 5
5 6
2 4 3 5 2 5 3 6 2 5 3 6 2 6
, 2 , 2 1.8 3
1.8 3 2.3 4 , 2.3 4 2.7 5
2.7 5 2.8 6 , 2.8 6 2.7 7
dP v v P dP v v P v P v P dP v P v P v P v P
dt dt dt
dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P v P
dt dt
= − + = − − + = − − +
= − − + = − − +
= − − + = − − + 3 7
7 8
2 6 3 7 2 7 3 8 2 7 3 8 2 8 3 9
9 10
2 8 3 9 2 9 3 10 2 9 3 10
2 3
2
3 4 0 3 4 0 0
2.7 7 2.3 8 , 2.3 8 1.8 9
1.8 9 10 , 10
([ ] / )
(e e ), (e e ) v
1 ([C
p CaN H
p p p p p p PKA
v P dP v P v P v P v P dP v P v P v P v P
dt dt
dP v P v P v P v P dP v P v P
dt dt
de k Ie k dI k Ie k I v Ca K I
dt dt
+
= − − + = − − +
= − − + = −
= − + − = − + − + −
+ 2 3
2 a ] / K )+ H
, where Pi shows the concentration of the i-fold phos-phorylated CaMKII, ep refers to the PP1 concentration which is not bounded to l1P and may demonstrate ac-tive protein phosphatase, the total concentration of PP1 is shown by ep0=0.1µM, free l1P is denoted by I, and free l1 concentration is shown by I0=0.1 µM. Also K3=1/ µMs
and K3=10-3/s are association and dissociation rate
con-stant of PP1-l1P complex, respectively. vcaN=2/s refers to the rate of l1P dephosphorylation due to calcineurin (CaN), VPKA=0.45µM/sis the phosphorylation rate of l1 due to the protein kinase A (PKA), and KH2=0.7µM is the calcium activation Hill constant of CaN.
Phosphorylation (V1), auto-phosphorylation (V2), and dephosphorylation (V3) rates can be represented by the following equations:
(15) 2 8
1 1 0
1 2 4 2
1
10 ([Ca ] / K ) (1 ([Ca ] / K ) )
H
H
k P
v = + ++
2 3 10 1 p M i k e v K iP = +
∑
2 4 1 12 2 4
1
([Ca ] / K ) 1 ([Ca ] / K ) H
H k
v = ++ +
, where k1=0.5/s is the l1 dependent regulation rate of PP1 and KH1=4 µM is the Hill constant of CaMKII for calcium activation. KM=20 µM and are K2=10/s the Michaelis and catalytic constants, respectively. Finally, phosphorylated CaMKII can be presented by:
(16) 10
1
.
ii
Ph CaMKII
P
=
=
∑
In the next part, simulation results are presented.
3. Results
The mathematical modeling approach simulation re-sults are presented in this section. Simulations were implemented in Matlab 2014a. To solve differential equations, the forward Euler method with constant step size of 0.05 ms is used. Synaptic glutamate is assumed as a periodic signal with a frequency of 5 Hz which each pulse has a duration of 4 ms. The amplitude of this signal is 0.2 mM and the simulation time is 10 s. In all Figures, 1 second of the simulation is depicted to clarify dynam-ics of the signals.
3.1. Normal and pathological conditions
One of the features of opioid-induced synaptic plastic-ity is the induction of LTP where normal LTP cannot occur (van Huijstee & Mansvelder, 2015). Therefore, stimulation signal (which is synaptic glutamate) is con-sidered as a pulse train with a frequency of 5 Hz and amplitude of 0.2 mM. Choosing these values are based on experimental observations used by Tewari and Majum-dar’s computational models (Tewari & Majumdar, 2012b; Tewari & Majumdar, 2012a).
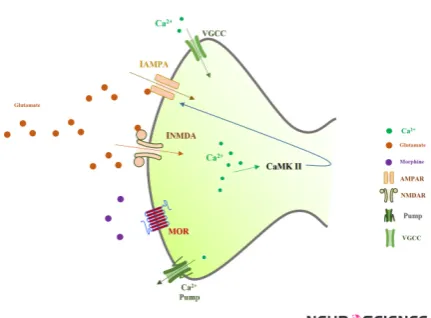
Figure 1. Spine of a CA1 pyramidal neuron which is
mod-eled in this work
As it is shown, synaptic glutamate activates AMPARs and NMDARs. By activation of these receptors, calcium enters into the neuron through VGCCs, NMDARs, and AMPARs. Activation of calcium pumps decreases calcium concentra-tion in the neuron. Elevaconcentra-tion of calcium may lead to ac-tivation of CaMKII mechanism which enhances AMPAR conductance.
280
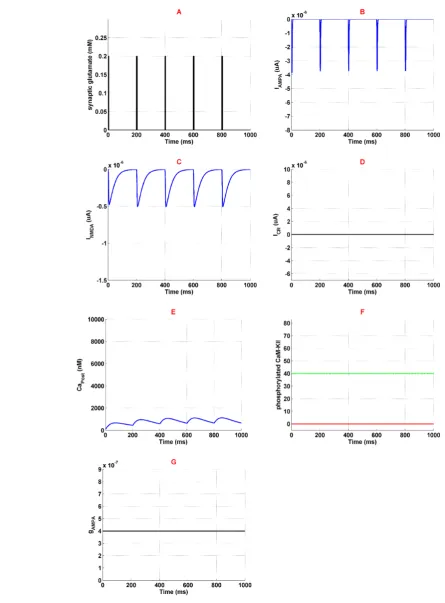
Figure 2. Activation of the neural components in normal condition
Panel A shows stimulation signal which is synaptic glutamate. Panel B and panel C demonstrate AMPAR and NMDAR medi-ated currents. Panel D is the current medimedi-ated by VGCC. Panel E shows calcium concentration variation in the neuron. Panel F shows phosphorylated CaMKII in redline, and green line is the threshold beyond which AMPAR can phosphorylate. Panel G demonstrates AMPAR's conductance which is constant. Simulation time is 1 second in all panels.
Basic and Clinical
281
July, August 2018, Volume 9, Number 4
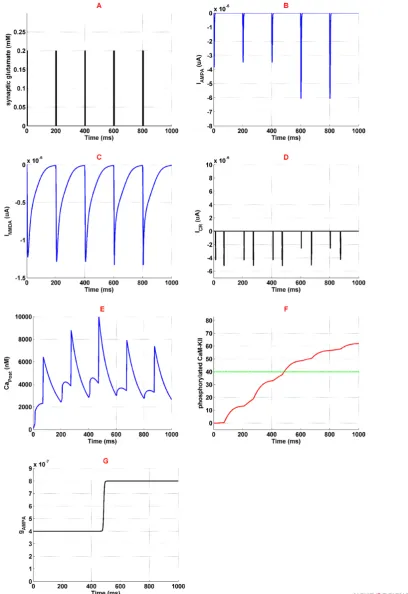
Figure 3. Activation of different neural components in the presence of 1 µM opioid
Panel A shows glutamate concentration which is similar to the normal condition. Panel B is the current mediated by AMPAR. Panel C shows current mediated by NMDAR. Panel D is the VGCC's current. Panel F shows phosphorylated CaMKII in red-line and threshold for AMPAR phosphorylation in green red-line. Finally, panel G is the AMPAR conductance. Simulation time is 1 second for all panels.
282
Applying synaptic glutamate (Figure 2a) leads to AM-PARs activation (Figure 2b) which depolarize membrane potential. Depolarized membrane allows NMDARs ac-tivation by removing Mg2+ blocking (Figure 2c). Since high-voltage calcium channels are considered in the mod-el, calcium channels activation is 0 which is described in
Figure 2d. Ions of Ca2+ can enter into the neuron through AMPARs and NMDARs activation. So, Ca2
concentra-tion enhances in the neuron which is shown in Figure 2e. Entered Ca2+ can engage in CaMKII phosphorylation cess, however, due to lack of enough calcium, this pro-cess is not observed here (Figure 2f). Based on simulation results, none of the AMPARs can phosphorylate (Figure 2g). Generally, this simulation can describe the normal condition which there is not any opioid’s effect in the syn-apse. In this simulation, LTP does not occur in the synapse due to lack of phosphorylated CaMKII and AMPARs.
Applying 1 μM of opioid increases NMDARs activation
through inhibition of Mg2+ blocking and enhancement of
receptors conductance. NMDAR mediated current enhances
in this situation (Figure 3c). Thus, more calcium enters into
the neuron (Figure 3e) which triggers CaMKII phosphory
-lation process (Figure 3f). Furthermore, more activation of
NMDARs results in VGCCs activation. Figure 3d shows calcium current mediated by VGCCs. Exceeding phosphory
-lated CaMKII from a predetermined threshold can phos
-phorylate AMPARs. In fact, AMPAR conductance increases which is shown in Figure 3g. Thus more current is mediated
by AMPARs (Figure 3b) that can work as a positive feedback
for enhancement of calcium concentration in the neuron.
3.2. Comparing different factors on pathological LTP induction
Simulation results show that injection of opioid can induce LTP when normal LTP does not occur. Now we want to assess which components of the model are re-sponsible in inducing such a pathological LTP. Based on these simulations we can suggest a way to prevent this type of pathological LTP.
Figure 4. Normalized values for phosphorylated CaMKII
and phosphorylated AMPAR in five different simulations
The plain black bar is the phosphorylated CaMKII, and the green bar with black patterns is the phosphorylated AM-PAR. Horizontal axes with “normal” label shows simulation in normal condition, “Opioid” label shows opioid injected condition, “CK=0” stands for the simulation in which opioid effect on NMDAR's conductance is assumed as 0, “CMg=0” describes the state that opioid effect on Mg2+ blocking is
as-sumed as 0 and “Icr=0” refers to the case that current medi-ated by VGCC is assumed as 0.
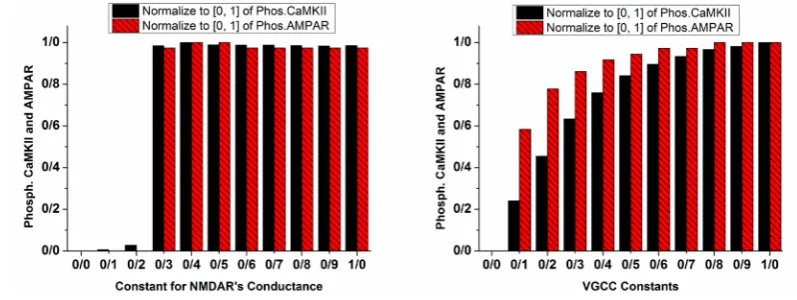
Figure 5. Phosphorylated CaMKII and AMPAR with variable constants for NMDAR's conductance (left panel) and VGCC (right panel)
Normalized values for phosphorylated CaMKII are in plain black bars, and normalized values of phosphorylated AMPAR
are in patterned red bars. Left panel demonstrates the conditions in which NMDAR's conductance coefficient varies with 0.1
increments. The right panel shows the conditions in which current mediated by VGCC is multiplied by the variable constant with 0.1 increments.
Basic and Clinical
283
July, August 2018, Volume 9, Number 4
Further simulations were performed to analyze differ-ent parameters involved in inducing pathological LTP. Normalized value for phosphorylated CaMKII and AM-PARs are calculated in each simulation. The values are calculated by using the following Equation:
(17) Min
Normalized
Max Min Y Y Y
Y Y
− =
−
Where YNormalized refers to the normalized value, Y is the parameter value, also YMax and YMin denote maximum and minimum values of the parameter, respectively. This normalization approach results in normalized values within [0-1] interval.
Opioid-induced LTP were analyzed with 3 different as
-sumptions: 1. Opioid does not modify NMDARs con-ductance; 2. Opioid does not have any infulence on Mg2+ blocking; and 3. VGCCs mediated current is 0. Simula-tion results according to the assumpSimula-tions are shown in
Figure 4. In normal condition, phosphorylated CaMKII
and AMPAR is 0 which specifies there is not any step foot of LTP in the synapse. Presence of opioid leads to phosphorylation of CaMKII and AMPAR, and normal-ized values are in the high state. This simulation demon-strates the presence of LTP in the synapse.
It can be seen that LTP does not occur in the synapse by neglecting opioid effect on NMDAR’s conductance (CK=0 label in Figure 4). Although, neglecting opioid’s effect on Mg2+ blocking (CMg=0 label in Figure 4) is not a critical parameter, knocking out VGCC’s activa-tion leads to attenuaactiva-tion of the LTP (Icr=0 label in Figure
4). These results show that enhancement of NMDAR’s conductance besides VGCC’s activation is a fundamen-tal parameter in inducing pathological LTP. Therefore in the next part, constants for VGCC’s activation and NM-DAR’s conductance are changed for further simulations.
The left panel of Figure 5 shows the normalized value for phosphorylated CaMKII and AMPAR with a vari-able coefficient of NMDAR’s conductance. It can be seen that when the coefficient is 0.2, there is not any phosphorylation; however, when it is 0.3, CaMKII and AMPAR phosphorylate. It means that attenuation of LTP by manipulating NMDAR’s conductance is im-possible. We can whether completely block LTP or not, and there is nothing in between. However, according to the simulation results shown in right panel of Figure 5, increment in VGCC mediated current strengthens LTP. As a matter of fact, increasing channels activation con-stant intensifies LTP.
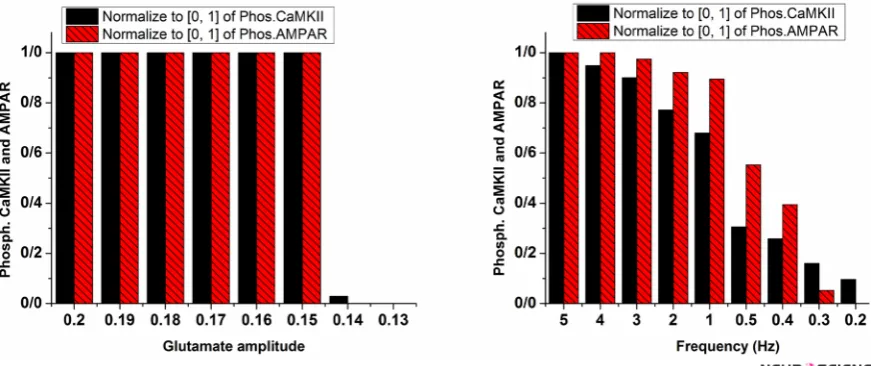
Besides opioid’s direct effect on postsynaptic neurons, change in frequency and amplitude of stimulation sig-nal which are considered as synaptic glutamate can al-ter pathological LTP. The left panel of Figure 6 shows the variation of glutamate on the CaMKII and AMPAR phosphorylation process. This simulation shows that re-ducing glutamate amplitude by 0.01 increments from 0.2 does not lead to inhibition of LTP until glutamate ampli-tude reaches 0.14. At this value, phosphorylated CaMKII and AMPARs decline rapidly, and LTP does not occur. However, a gradual decrease in stimulation frequency leads to gradual descend in LTP induction shown in the right panel of Figure 6.
Figure 6. Phosphorylated CaMKII and AMPAR with variable amplitude (left panel) and frequency of glutamate (Right panel) Normalized values for phosphorylated CaMKII are in plain black bars, and normalized values of phosphorylated AMPARs are in patterned red bars. In the left panel, glutamate amplitude decreased from 0.2 to 0.13 mM. In the right panel, frequency of glutamate pulse train decreased from 5 to 0.1 Hz.
284
4. Discussion
Our simulation results support the hypothesis that opioids consumption can lead to pathological memory formation in hippocampal neurons through induction of LTP. To explain this issue, we developed a computational model in which CaMKII and AMPAR phosphorylation processes are considered as foot prints of LTP induction.
Experimental observations (Chen & Huang, 1992; Chen & Marine, 1991;Kow et al., 2002;Mao, 1999) indicate the opioids effects on NMDARs. Enhancement of NM-DAR mediated current is a byproduct of opioids presence in synapses. Our simulation results showed that enhance-ment of NMDAR’s conductance is more efficient in induc-ing LTP compared to inhibition of Mg2+ blocking through opioids. In other words, despite the inhibitory effect of Mg2+ blocking on NMDAR mediated current, we ob-served that the main reason of LTP induction is enhance-ment of NMDA receptors conductance. Furthermore, our results suggest that prevention of pathological LTP can be achieved by blocking NMDAR’s conductance. NMDAR’s conductance acts like a none-or-all switch in inducing LTP. Therefore, it is impossible to attenuate pathological LTP by decreasing NMDAR’s conductance. However, we may completely block it using this approach.
Bongianni et al. (1986) and Mishra et al. (2017), through their experimental observations, suggested that blocking VGCCs leads to preventing dependency on opioids. In this research, we obtained the same result but in a different way, i.e., we showed the role of VGCCs in forming pathological LTP. Our simulation results demonstrate that by inhibiting VGCCs, pathological LTP is attenuated. Also, we may infer that by decreasing VGCC activation, pathological LTP is gradually lessened. One may be able to apply this finding in cases where one intends or prefers to attenuate pathological LTP rather than blocking it.
On the other hand, disinhibitory effect of opioid in-creases glutamate concentration in the synaptic cleft. Therefore, it is reasonable to assume that changes in the amplitude and the frequency of the stimulation signal in the postsynaptic neuron are due to the presence of the opioid. Simulation results demonstrate that increasing the frequency and the amplitude of the stimulation sig-nal results in LTP induction. There are different experi-mental observations that support these simulation results
(Akaishi et al., 2000;Capogna et al., 1993; Caudle &
Chavkin, 1990;Cohen et al., 1992;Kauer & Malenka,
2007;Rezai et al., 2013).
These observations indicate that opioid inhibits the GA-BAergic signal that leads to enhancement of glutamate concentration in the synapse. Furthermore, our simu-lation results suggest that reducing synaptic glutamate can prevent pathological LTP induction. Amplitude and frequency of glutamate release are parameters that will help in preventing or attenuating LTP. Since the source of glutamate is mainly from presynaptic neurons, inhibi-tion of presynaptic pyramidal neurons may help prevent pathological LTP induction. On the other hand, one of the fundamental functions of the glial cells is reuptake of the synaptic glutamate. Therefore, we also suggest con-sidering the role of glial cells in preventing pathological memory formation. This goal can be achieved through stimulating glutamate transporters. Using this approach, synaptic glutamate can be reduced, and this can result in reducing stimulation of the postsynaptic receptors.
In this paper, we introduced a computational model for postsynaptic neurons in CA1 region of the hippocampus to examine LTP induction due to opioids. The model is consisted of a spine which produces EPSPs. NMDAR, AMPAR and VGCC’s mediated currents are also con-sidered in the model. Calcium ions flow into the neuron through these channels and receptors. Elevation of cal-cium concentration leads to CaMKII and AMPAR phos-phorylation. We assumed that CaMKII and AMPAR phosphorylation are foot prints of LTP induction.
Simulation results of the presented model indicate that presence of opioids can lead to LTP induction in a situ-ation that normal LTP does not occur. This finding sup-ports the idea that opioids may form a memory that is directly linked to drug consumption. Therefore, one of the primary triggers of relapse in withdrawal time is this pathological memory. Furthermore, we observed that, by decreasing VGCC’s activation, NMDAR’s activation, and glutamate concentration in the synapse, pathological LTP reduces and it is even possible to prevent the patho-logical LTP.
Ethical Considerations
Compliance with ethical guidelines
Since the research was a simulation one, the authors did not need to get any ethical approvals.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Basic and Clinical
285
July, August 2018, Volume 9, Number 4
Conflict of interest
The authors declared no conflict of interest.
References
Akaishi, T., Saito, H., Ito, Y., Ishige, K., & Ikegaya, Y. (2000). Morphine augments excitatory synaptic transmission in the dentate gyrus through GABAergic disinhibition. Neuroscience Research, 38(4), 357-63. [DOI:10.1016/S0168-0102(00)00177-2]
Amiri, M., Bahrami, F., & Janahmadi, M. (2012). On the role of astrocytes in epilepsy: A functional modeling approach. Neuroscience Research, 72(2), 172-80. [DOI:10.1016/j.neue-res.2011.11.006] [PMID]
Amiri, M., Montaseri, G., & Bahrami, F. (2011). On the role of astrocytes in synchronization of two coupled neurons: A mathematical perspective. Biological Cybernetics, 105(2),
153-66. [DOI:10.1007/s00422-011-0455-5] [PMID]
Bao, G., Kang, L., Li, H., Li, Y., Pu, L., Xia, P., et al. (2007). Mor-phine and heroin differentially modulate in vivo hippocam-pal LTP in opiate-dependent rat. Neuropsychopharmacology, 32(8), 1738-49. [DOI:10.1038/sj.npp.1301308] [PMID]
Bongianni, F., Carla, V., Moroni, F., & Pellegrini‐Giampietro, D. (1986). Calcium channel inhibitors suppress the morphine‐ withdrawal syndrome in rats. British Journal of Pharmacol-ogy, 88(3), 561-7. [DOI:10.1111/j.1476-5381.1986.tb10236.x] [PMID] [PMCID]
Borjkhani, M., Mahdavi, A., & Bahrami, F. (2014). A mathemati-cal model for neuron astrocytes interactions in hippocampus during addiction. Paper presented at the Iranian Conference on Bio-medical Engineering (ICBME), Tehran, Iran, 26-28 November 2014. [DOI:10.1109/ICBME.2014.7043940]
Capogna, M., Gähwiler, B. H., & Thompson, S. M. (1993). Mech-anism of mu-opioid receptor-mediated presynaptic inhibi-tion in the rat hippocampus in vitro. The Journal of Physiology, 470(1), 539-58.[DOI:10.1113/jphysiol.1993.sp019874]
Castellani, G. C., Quinlan, E. M., Bersani, F., Cooper, L. N., & Shouval, H. Z. (2005). A model of bidirectional synaptic plasticity: From signaling network to channel conductance. Learning & Memory, 12(4), 423-32. [DOI:10.1101/lm.80705] [PMID] [PMCID]
Castellani, G. C., Quinlan, E. M., Cooper, L. N., & Shouval, H. Z. (2001). A biophysical model of bidirectional synap-tic plassynap-ticity: dependence on AMPA and NMDA receptors. Proceedings of the National Academy of Sciences, 98(22), 12772-7.
[DOI:10.1073/pnas.201404598] [PMID] [PMCID]
Caudle, R., & Chavkin, C. (1990). Mu opioid receptor activation reduces inhibitory postsynaptic potentials in hippocampal CA3 pyramidal cells of rat and guinea pig. Journal of Pharma-cology and Experimental Therapeutics, 252(3), 1361-9. [PMID]
Chen, L., & Huang, L. Y. M. (1992). Protein kinase C reduces Mg2+ block of NMDA-receptor channels as a mechanism of modu-lation. Nature, 356(6369), 521-3. [DOI:10.1038/356521a0]
Chen, L., & Marine, L. Y. M. H. (1991). Sustained potentiation of NMDA receptor-mediated glutamate responses through
acti-vation of protein kinase C by a μ opioid. Neuron, 7(2), 319-26.
[DOI:10.1016/0896-6273(91)90270-A]
Cohen, G. A., Doze, V. A., & Madison, D. V. (1992). Opi-oid inhibition of GABA release from presynaptic termi-nals of rat hippocampal interneurons. Neuron, 9(2), 325-35.
[DOI:10.1016/0896-6273(92)90171-9]
Contet, C., Kieffer, B. L., & Befort, K. (2004). Mu opioid receptor: A gateway to drug addiction. Current Opinion in Neurobiology, 14(3), 370-8. [DOI:10.1016/j.conb.2004.05.005] [PMID]
Destexhe, Z. F. M. A., & Sejnowski, T. J. (1998). Kinetic models of synaptic transmission. In C. Koch, & I. Segev (Eds.), Methods in Neuronal Modeling (pp. 1-25). Cambridge: MIT Press.
Dong, Y., & Nestler, E. J. (2014). The neural rejuvenation hy-pothesis of cocaine addiction. Trends in Pharmacological Sci-ences, 35(8), 374-83. [DOI:10.1016/j.tips.2014.05.005] [PMID] [PMCID]
Ga, B. (1980). Excitatory action of opioid peptides and opiates on cultured hippocampal pyramidal cells. Brain Research, 194(1), 193-203. [DOI:10.1016/0006-8993(80)91328-1]
Harris, G., Wimmer, M., Byrne, R., & Aston-Jones, G. (2004). Glutamate-associated plasticity in the ventral tegmental area is necessary for conditioning environmental stimuli with morphine. Neuroscience, 129(3), 841-7. [DOI:10.1016/j.neuroo-science.2004.09.018] [PMID]
Heidari, M. H., Amini, A., Bahrami, Z., Shahriari, A., Movafag, A., & Heidari, R. (2013). Effect of chronic morphine consump-tion on synaptic plasticity of rat’s hippocampus: A transmis-sion electron microscopy study. Neurology Research Internation-al, 2013, 290414. [DOI:10.1155/2013/290414] [PMID] [PMCID]
Herman, B. H., Frankenheim, J., Litten, R. Z., Sheridan, P. H., Weight, F. F., & Zukin, S. R. (2003). Glutamate and addiction. New York: Humana Press.
Hosseinmardi, N., Azimi, L., Fathollahi, Y., Javan, M., & Naghdi, N. (2011). In vivo sodium salicylate causes tolerance to acute morphine exposure and alters the ability of high frequency stimulation to induce long-term potentiation in hippocampus area CA1. European Journal of Pharmacology, 670(2), 487-94.
[DOI:10.1016/j.ejphar.2011.09.008] [PMID]
Hosseinmardi, N., Fathollahi, Y., Naghdi, N., & Javan, M. (2009). Theta pulse stimulation: a natural stimulus pattern can trigger long-term depression but fails to reverse long-term potentia-tion in morphine withdrawn hippocampus area CA1. Brain Re-search, 1296, 1-14. [DOI:10.1016/j.brainres.2009.08.020] [PMID]
Kauer, J. A., & Malenka, R. C. (2007). Synaptic plastic-ity and addiction. Nature Reviews Neuroscience, 8(11), 844-58.
[DOI:10.1038/nrn2234] [PMID]
Kelley, A. E. (2004). Memory and addiction: Shared neural circuitry and molecular mechanisms. Neuron, 44(1), 161-79.
[DOI:10.1016/j.neuron.2004.09.016] [PMID]
Kow, L. M., Commons, K., Ogawa, S., & Pfaff, D. (2002). Po-tentiation of the excitatory action of NMDA in
ventrolat-eral periaqueductal gray by the μ-opioid receptor agonist,
DAMGO. Brain Research, 935(1), 87-102. [DOI:10.1016/S0006-8993(02)02532-5]
286
Li, J., Tang, J., Ma, J., Du, M., Wang, R., & Wu, Y. (2016).
Dy-namic transition of neuronal firing induced by abnormal
astrocytic glutamate oscillation. Scientific Reports, 6(1), 32343.
[DOI:10.1038/srep32343]
Lisman, J., Yasuda, R., & Raghavachari, S. (2012). Mechanisms of CaMKII action in long-term potentiation. Nature Reviews Neurosci-ence, 13(3), 169-82. [DOI:10.1038/nrn3192] [PMID] [PMCID]
Lüscher, C., & Malenka, R. C. (2011). Drug-evoked synaptic plasticity in addiction: From molecular changes to circuit remodeling. Neuron, 69(4), 650-63. [DOI:10.1016/j.neuu-ron.2011.01.017] [PMID] [PMCID]
Mameli, M., Bellone, C., Brown, M. T., & Lüscher, C. (2011). Co-caine inverts rules for synaptic plasticity of glutamate trans-mission in the ventral tegmental area. Nature Neuroscience, 14(4), 414-6. [DOI:10.1038/nn.2763] [PMID]
Mao, J. (1999). NMDA and opioid receptors: their interactions in antinociception, tolerance and neuroplasticity. Brain Research Reviews, 30(3), 289-304. [DOI:10.1016/S0165-0173(99)00020-X]
Mishra, P. R., Barik, M., & Ray, S. B. (2017). Effect of nimodipine on morphine-related withdrawal syndrome in rat model: An observational study. Journal of Pediatric Neurosciences, 12(1), 7.
[DOI:10.4103/1817-1745.205652] [PMID] [PMCID]
Moradi, K., Moradi, K., Ganjkhani, M., Hajihasani, M., Gharibza-deh, S., & Kaka, G. (2013). A fast model of voltage-dependent NMDA receptors. Journal of Computational Neuroscience, 34(3), 521-31. [DOI:10.1007/s10827-012-0434-4] [PMID]
Nestler, E. J. (2001). Molecular basis of long-term plasticity un-derlying addiction. Nature Reviews Neuroscience, 2(2), 119-28.
[DOI:10.1038/35053570] [PMID]
Nestler, E. J. (2013). Cellular basis of memory for addiction. Dia-logues in Clinical Neuroscience, 15(4), 431. [PMID] [PMCID]
Peters, J., & De Vries, T. J. (2012). Glutamate mechanisms un-derlying opiate memories. Cold Spring Harbor Perspectives in Medicine, 2(9), a012088. [DOI:10.1101/cshperspect.a012088] [PMID] [PMCID]
Przewlocki, R., Parsons, K. L., Sweeney, D. D., Trotter, C., Net-zeband, J. G., Siggins, G. R., et al. (1999). Opioid enhancement of calcium oscillations and burst events involving NMDA receptors and L-type calcium channels in cultured hip-pocampal neurons. The Journal of Neuroscience, 19(22), 9705-15.
[DOI:10.1523/JNEUROSCI.19-22-09705.1999] [PMID]
Rezai, X., Kieffer, B. L., Roux, M. J., & Massotte, D. (2013). Delta opioid receptors regulate temporoammonic-activated feed-forward inhibition to the mouse CA1 hippocampus. PloS One, 8(11), e79081. [DOI:10.1371/journal.pone.0079081] [PMID] [PMCID]
Sanhueza, M., & Lisman, J. (2013). The CaMKII/NMDAR complex as a molecular memory. Molecular Brain, 6(1), 10.
[DOI:10.1186/1756-6606-6-10] [PMID] [PMCID]
Tewari, S., & Majumdar, K. (2012a). A mathematical model for astrocytes mediated LTP at single hippocampal syn-apses. Journal of Computational Neuroscience, 33(2), 341-70.
[DOI:10.1007/s10827-012-0389-5] [PMID]
Tewari, S. G., & Majumdar, K. K. (2012b). A mathematical mod-el of the tripartite synapse: Astrocyte-induced synaptic plas-ticity. Journal of Biological Physics, 38(3), 465-96. [DOI:10.1007/ s10867-012-9267-7] [PMID] [PMCID]
Trujillo, K. A. (2002). The neurobiology of opiate tolerance, de-pendence and sensitization: Mechanisms of NMDA receptor-dependent synaptic plasticity. Neurotoxicity Research, 4(4), 373-91. [DOI:10.1080/10298420290023954] [PMID]
Tsodyks, M. V., & Markram, H. (1997). The neural code between neocortical pyramidal neurons depends on neurotransmitter release probability. Proceedings of the national academy of sci-ences, 94(2), 719-23. [DOI:10.1073/pnas.94.2.719]
van Huijstee, A. N., & Mansvelder, H. D. (2015). Glutamatergic synaptic plasticity in the mesocorticolimbic system in addic-tion. Frontiers in Cellular Neuroscience, 8, 466. [DOI:10.3389/ fncel.2014.00466]
Volman, V., Bazhenov, M., & Sejnowski, T. J. (2012). Compu-tational models of neuron-astrocyte interaction in epilepsy. Frontiers in Computational Neuroscience, 6, 58. [DOI:10.3389/ fncom.2012.00058] [PMID] [PMCID]
Williams, J. T., Christie, M. J., & Manzoni, O. (2001). Cellular and synaptic adaptations mediating opioid dependence. Physiological Reviews, 81(1), 299-343. [DOI:10.1152/physs-rev.2001.81.1.299] [PMID]
Wolf, M. E. (2003). LTP may trigger addiction. Molecular Inter-ventions, 3(5), 248. [DOI:10.1124/mi.3.5.248] [PMID]
Zhabotinsky, A. M. (2000). Bistability in the Calmodulin-de-pendent protein kinase-phosphatase system. Biophysical Jour-nal, 79(5), 2211-21. [DOI:10.1016/S0006-3495(00)76469-1]
Basic and Clinical
287
July, August 2018, Volume 9, Number 4
Appendix. Numerical values of the model parameters
Variable Description Value (Units) Source
Rm Actual resistance of the spine-head 0.79*105 MΩ (Tsodyks & Markram, 1997)
Vrest
post Postsynaptic resting membrane potential -70 mv (Tsodyks & Markram, 1997) τpost Postsynaptic membrane time constant 50 ms (Tsodyks & Markram, 1997)
VAMPA AMPAR reversal potential 0 mv (Destexhe et al., 1998)
αAMPA AMPAR forward rate constant 1.1 µMs-1 (Destexhe et al., 1998)
βAMPA AMPAR backward rate constant 190s-1 (Destexhe et al., 1998)
VNMDA NMDAR reversal potential 0 mv (Destexhe et al., 1998)
F Faraday’s constant 96487 Cmol-1 (Moradi et al., 2013)
R Real gas constant 8.314 J/K (Moradi et al., 2013)
T Absolute temperature 293.15 K (Moradi et al., 2013)
Z Valence of Mg2+ 2 (Moradi et al., 2013)
δ Relative electrical distance of the binding site of Mgoutside of the membrane 2+ from the 0.8 (Moradi et al., 2013)
K0 IC50 at 0 mV 4.1 µM (Moradi et al., 2013)
[Mg2+]
0 Concentration of Mg2+ n the extracellular fluid 1 µM (Moradi et al., 2013) αNMDA NMDAR forward rate constant *107.24 M-1 s-1 (Destexhe et al., 1998)
βNMDA NMDAR backward rate constant 6.6 s-1 (Destexhe et al., 1998)
K Steepness of speed of voltage-dependent gating of channel 0.007 (Moradi et al., 2013)
V0 The Vm at which gVD,∞ is equal to 0 -100 mv (Moradi et al., 2013)
τg Time constant which determines the speed of voltage-dependent gating of channel 0.05 ms (Moradi et al., 2013)