Annotation: What electrical brain activity tells
us about brain function that other techniques
cannot tell us – a child psychiatric perspective
Tobias Banaschewski
1and Daniel Brandeis
21Child and Adolescent Psychiatry, University of Go¨ttingen, Germany;2Child and Adolescent Psychiatry, and Centre
for Integrative Human Physiology, University of Zu¨ rich, Switzerland
Background: Monitoring brain processes in real time requires genuine subsecond resolution to follow the typical timing and frequency of neural events. Non-invasive recordings of electric (EEG/ERP) and magnetic (MEG) fields provide this time resolution. They directly measure neural activations associated with a wide variety of brain states and processes, even during sleep or in infants. Mapping and source estimation can localise these time-varying activation patterns inside the brain.Methods:Recent EEG/ ERP research on brain functions in the domains of attention and executive functioning, perception, memory, language, emotion and motor processing in ADHD, autism, childhood-onset schizophrenia, Tourette syndrome, specific language disorder and developmental dyslexia, anxiety, obsessive-compulsive disorder, and depression is reviewed. Results: Over the past two decades, electrophysiology has substantially contributed to the understanding of brain functions during normal development, and psychiatric conditions of children and adolescents. Its time resolution has been important to measure covert processes, and to distinguish cause and effect. Conclusions: In the future, EEG/ERP para-meters will increasingly characterise the interplay of neural states and information processing. They are particularly promising tools for multilevel investigations of etiological pathways and potential predictors of clinical treatment response. Keywords: ADHD, anorexia nervosa, anxiety, autism, childhood-onset schizophrenia, depression, developmental dyslexia, EEG, endophenotypes, ERP, fMRI, neuropsycho-logy, obsessive-compulsive disorder, specific language disorder, tic disorder. Abbreviations: ACC: anterior cingulate gyrus; ADHD: attention deficit hyperactivity disorder; ASD: autism spectrum disorder; BOLD: blood-oxygen-level-dependent; CNV: contingent negative variation; COMT: Catechol-O-methyltransferase; CPT A-X: (cued) continuous performance test A-X (type); CHRM2: cholinergic receptor, muscarinic 2; DRD2: D2 receptor gene; EEG: electroencephalogram; ERN: error-related nega-tivity; ERP: event-related potential; fMRI: functional Magnetic Resonance Imaging; GABRA2: GABA-A receptor gene; LDAEP: loudness-dependent amplitude change of auditory evoked ERP; MDD: major depressive disorder; MEG: magnetoencephalogram; MMN: mismatch negativity; NIRS: Near Infrared Spectroscopy; PET: Positron Emission Tomography; REM: rapid eye movement; RP: readiness potential (or Bereitschaftspotential); SSRI: selective serotonine reuptake inhibitor.
Electrophysiological and magnetencephalographic methods allow non-invasive monitoring of brain processes in real time. Their supreme time resolu-tion captures even fast neural events and high-frequency oscillations, and permits functional brain imaging at millisecond resolution through source analysis of high density maps. As a consequence, they can directly measure the full spatio-temporal dynamics of neural activation (see: methodological section) associated with a wide variety of cognitive processes and their development, and can be used repeatedly without problems in longitudinal studies and even during sleep or in infants (for reviews see: Picton et al., 2000; Picton & Hillyard, 1988; Taylor & Baldeweg, 2002). Over the past two decades, electrophysiology has increasingly been used for investigating the biological substrates of psychiatric disorders.
The aim of this paper is to illustrate the current state of the art of electrophysiological brain imaging (here: EEG and ERP) in child and adolescent psy-chiatry. It will be shown how this research has
con-tributed substantially to our understanding of brain states and functions in normal development, and of deviations in psychiatric conditions occurring in childhood or adolescence.
The paper starts with an introduction to meth-odological issues and contrasts advantages and disadvantages of electroencephalogram (EEG) and event-related potentials (ERP) methodology with those of other neuroimaging techniques and neuro-psychology. Secondly, recent advances of EEG/ERP research in the domains of attention, executive functions, perception, memory, language, emotion, and motor processing are reviewed. This selective review is not comprehensive, but rather aims to illustrate the key issues through exemplary findings in child psychiatry, focusing on ERP research. The selection of studies was based (a) on the presence of findings that contribute to theoretical controversies or clinical treatment issues, (b) on the quality of the samples investigated and the analyses used, and (c) on whether a paper clearly illustrates specific methodological strengths or weaknesses of EEG/
ERP research. We only included those studies with adults which were exemplary and lacked an analo-gous equivalent in child psychiatric findings.
In an outlook, the annotation will then give examples how electrophysiological parameters have been (or may be) used to predict pharmacological treatment response and to identify endophenotypes for multilevel investigations of etiological pathways.
Methodology
The EEG measures brain electrical activity as recorded from electrodes placed on the scalp. It di-rectly reflects neural mass action, and mainly syn-chronised postsynaptic potentials of aligned neurons such as pyramidal cells in cortex. Such spatio-temporal synchronisation of neural networks results in net polarisation of extended brain regions, which may be transient, slow, or oscillatory. The largest EEG potentials are generated by cortical sources which vary in depth and orientation, reflecting the cortical folding. The attenuation of EEG activity with depth is moderate, and far less than for the MEG. The clinical routine of auditory brain stem potential recordings demonstrates that even activity from small and deep subcortical structures is reliably detected in the scalp EEG after averaging (see below). In contrast, action potentials are unlikely to con-tribute to the scalp EEG, because their fields are radially symmetric around the axons and thus can-cel at some distance despite their large amplitude. Although an influential model has related surface polarity of slow ERPs to neural inhibition (positivity, as during the frontocentral NoGo P300) or activation (negativity, as during the pre-central CNV) of nearby cortical regions (Birbaumer, Elbert, & Canavan, 1990), tomographic source models of brain electrical activity (Pascual-Marqui, Michel, & Lehmann, 1994; Strik, Fallgatter, Brandeis, & Pascual-Marqui, 1998) and fMRI correlates of slow ERPs (Hinterberger et al., 2003) indicate a more complex relationship between the local polarity of scalp maps, which strongly
depends on local cortical folding, and the underlying distributed neural networks which may involve a distributed pattern of activations and deactivations. EEG waveforms generally are classified according to their dominant frequency (see Table 1), ampli-tude, shape, and the sites of scalp distribution (topography).
The EEG frequency composition is highly heritable (van Beijsterveldt & van Baal, 2002). It reflects the state of development and wakefulness, along with modulations due to state regulation and arousal. The magnetoencephalogram (MEG) measures the corresponding magnetic fields with sensors around the head. Although electric and magnetic fields are typically closely related, the MEG is insensitive to radial brain sources and skull conductivities, and less sensitive to deep sources, thus (compared to EEG recording) providing a more selective view of the same neural mass activity.
ERP are changes in the ongoing EEG or MEG which are time-locked (i.e., stimulus- or response-locked) and phase-locked (i.e., time-locked and with the same polarities) to perceptual, cognitive, and motor processes. They are typically extracted from the ongoing electroencephalogram by means of sig-nal averaging (see glossary). Averaging not only eliminates the spontaneous background EEG, but also those event-related EEG modulations which are not phase-locked as ‘noise’. Other techniques can also characterise such event-related modulations of specific frequencies (event-related synchronisation or desynchronisation), stimulus-induced phase-locking (i.e., stability of the evoked EEG rhythms independently of amplitude measures), or the power (i.e., the ‘strength’) within defined frequency bands of the spontaneous EEG.
ERP consist of characteristic sequences of com-ponents or ‘microstates’ (i.e., time segments with a stable topographical distribution of brain electrical activity) that span a continuum between early activity primarily determined by the physical char-acteristics of the eliciting stimulus (latency range <250 ms), and later components (latency range
Table 1 Overview of EEG frequency bands, their topography, developmental and functional characteristics, and exemplary findings EEG
Frequency bands Delta (<4 Hz) Theta (4–7 Hz) Alpha (8–12 Hz) Beta (13–30 Hz) Gamma (30–70 Hz)
Topography Posterior Frontal
Developmental characteristics
Predominant during neonatal period & early childhood
Increases until early adolescence
Continues to mature until adulthood
Functional state e.g., sleep, decreased vigilance e.g., relaxation e.g., concentration,
neuronal excitability
e.g., feature binding
Exemplary findings ADHD: increased slow wave
activity
Associated with GABA-A receptor polymorphism Risk for alcohol
dependence associated with increased beta ADHD: EEG subtype with increased beta Cholinergic muscarinic
recep-tor gene related to theta and delta event-related oscillations
>250 ms) dominated by cognitive rather than phy-sical characteristics of the stimuli (Brandeis & Leh-mann, 1986; Picton & Hillyard, 1988). These components are characterised by their topography, polarity and amplitude, by their latency, and their functional significance (see Table 2a and 2b for an overview).
Mapping the scalp topographies of electric poten-tials or current source densities (also called surface Laplacians, acting as spatial high pass filters em-phasising localised components) gives important clues to the intracranial generators of brain electrical activity which determine map topography. Topo-graphically stable micro-states usually coincide with the conventional component peaks. Separate meas-ures of map topography and of map strength (i.e., Global Field Power) provide a simplified description of the map (Brandeis, Vitacco, & Steinhausen, 1994; Lehmann, 1987).
Brain sources of scalp recorded ERP can be estim-ated from EEG or MEG topographies through source modelling. However, the inverse problem (i.e., cal-culating sources from the known scalp distribution) allows for multiple solutions which fit the data equally well. Their validity can be considerably im-proved by allowing not only focal (e.g., dipole based)
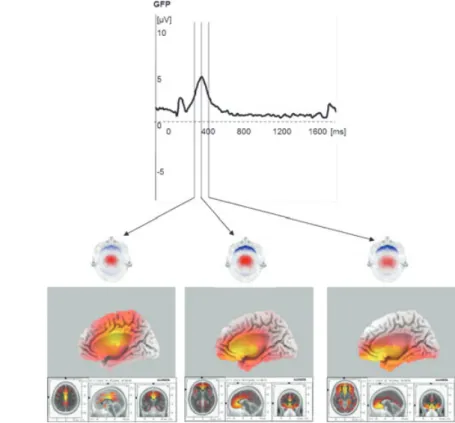
but also distributed (tomographic) solutions, and by taking neurophysiological and anatomical con-straints into account (Michel et al., 2004; Pascual Marqui, 2002; Pascual-Marqui et al., 1999). Ab-solute localisation errors for focal sources are usu-ally around 1 cm and remain well below 2 cm even for deep sources. Due to their complementary sen-sitivities, combined EEG and MEG recordings fur-ther facilitate source localisation (Fuchs et al., 1998). See Figure 1.
EEG/ERP and neuropsychology
Performance data alone can only give indirect clues about covert processing. Different covert mechan-isms leading to similar overt performance may ap-pear indistinguishable, and deviant covert information processing may precede overt performance deficits or even underlie normal performance. In addition, many tasks tap more than one latent dimension of functioning (Banaschewski et al., 2005).
Thus, the construct validity (i.e., do they measure what they mean to measure) of neuropsychological tasks is often uncertain. In contrast, EEG/ERP studies can reveal both psychophysiological precur-sors and correlates of poor performance, and
meas-Figure 1 Time course, scalp distribution, and sources of an ERP. Theactivity(GFP curve, top) peaks at 380 ms with the NoGo P300 to O-not X trials in the CPT O-X following smaller visual evoked potential components. Themaps (middle, red positive, blue negative potentials, scaled to maximum) demonstrate a stable central positivity and increasing prefrontal negativity during the NoGo P3 at 330, 380 and 430 ms. Thesources(bottom, maximal activity in yellow and indicated by cursors in slices) suggest anterior cingulate cortex (ACC) activation progressing towards increasingly anterior brain locations. Grand mean 48-channel ERP data of 12 young adult controls from an ongoing study. Children show a similar but shorter and less prominent NoGo P300, leading to less reliable source localization. Source tomography with standardised sLORETA, assuming SNR 10; Pascual Marqui, 2002
ure covert attention even in the absence of an overt response (Brandeis et al., 1998; van Leeuwen et al., 1998). Moreover, EEG/ERP parameters can index processing of task-irrelevant, distracting or unat-tended stimuli, and may be recorded even during sleep, unconsciousness, or in infants. They may indicate whether task performance reflects the same underlying processes across development and in various populations. Thus, EEG/ERP studies increasingly are being used (a) to illuminate the content validity of neuropsychological tests by revealing the modular architecture of more complex neuropsychological constructs, (b) to test alternative models and (c) to constrain psychological theories.
EEG/ERP and other imaging techniques
Neuroimaging techniques based on cerebral blood flow or metabolism excel at spatial resolution, but lack the genuine and high temporal resolution pro-vided by EEG/ERP to reveal the exact timing, sequencing or oscillation frequency of covert brain processes and their neural substrates. In addition, neuroimaging methods such as NIRS, fMRI or PET reflect delayed metabolic consequences of the neural activity while the latter is measured directly by EEG/ MEG.
The native time resolution of EEG/MEG (vs. fMRI) and the native spatial resolution of fMRI (vs. EEG/ MEG) continue to differ by an order of magnitude. Some authors also caution that they reflect different aspects of neural processing operating at different time scales, which can at best provide converging evidence, and thus require integration through models rather than through direct fusion (Horwitz & Poeppel, 2002). On the other hand, progress in both event-related fMRI (faster temporal sampling and deconvolution) and in EEG/MEG (denser spatial sampling and source tomographies) source imaging has brought together the effective spatio-temporal resolution of these methods. Recent multimodal animal research has established that local EEG and fMRI responses are closely coupled during visual stimulation (Logothetis, Pauls, Augath, Trinath, & Oeltermann, 2001), and multimodal (EEG-fMRI) human studies demonstrate statistical correspon-dence (Brem et al., 2006; Vitacco, Brandeis, Pascual-Marqui, & Martin, 2002). Such multimodal research also highlights the fact that the accurate localisation with fMRI is necessary, but not sufficient to the un-derstanding of brain function unless complemented by the high temporal resolution of EEG/MEG, be-cause initial and re-entrant brain activation in the same brain region occurs within less than 100 ms and cannot be disentangled by the low time resolu-tion of fMRI alone (Noesselt et al., 2002). Recent developmental work combining EEG and fMRI in the same subjects is particularly relevant in this context. These studies illustrate the importance and the sensitivity of EEG-based measures in a
develop-mental context, and provide intriguing evidence for a two-stage model of late maturation. The distribution of networks for visual, motor and even reading functions (as measured by fMRI) seems largely established before adolescence, but fine-tuning of specific neural networks in terms of specialisation and processing speed (as measured by ERPs) and the deactivation of non-specific ‘default’ networks (measured by both ERPs and fMRI) continue beyond adolescence (Brem et al., 2006; Bucher et al., in press; Halder et al., in press).
The temporal and spatial features of the adult EEG are linked through the distinct topographies and sources of the EEG frequency bands, with the most occipital and superior locations for alpha and the most frontal and inferior locations for delta (Michel, Henggeler, & Lehmann, 1992a; Michel, Lehmann, Henggeler, & Brandeis 1992b). How distributed neuronal networks correlate with the EEG frequency bands is becoming increasingly clear through sim-ultaneous EEG-fMRI recordings. This method has provided an impressive verification that EEG fre-quency bands indeed reflect activity of cortical and subcortical networks hypothesised to be involved in regulation of attention and arousal, respectively; higher frequencies are related to localised cortical activation, while lower frequencies are more related to deactivation or inhibition. Increased occipital alpha activity during the resting state has been mapped onto increased thalamic and decreased occipital, temporal, and frontal fMRI activity in adults (Gold-man, Stern, Engel, & Cohen, 2002; Moosmann et al., 2003), and mainly increased frontal (dorsal medial) and posterior (cingulate and precuneus) fMRI activ-ity with increased beta at occipital leads (Laufs et al., 2003).
Importantly, the EEG can characterise functional states directly through absolute calibrated measures of amplitude in the time or frequency domain. Such calibrated measures are not possible with BOLD (blood-oxygen-level-dependent) fMRI, which can only characterise the differences between states, and re-quires calibration with regard to yet another state (Davis, Kwong, Weisskoff, & Rosen, 1998). Absolute calibrated measures of brain activity can also be obtained by other imaging techniques such as PET. However, PET applications in child psychiatry are particularly limited due to the use of radioactive markers, along with the high cost and technical demands.
Brain functions and applications – EEG
Resting state, activationThe EEG changes with age and state of activation. Maturation is accompanied by a reduction of the slower components, whereas drowsiness and cere-bral dysfunction both produce slowing. The typical resting EEG is recorded during relaxed wakefulness
with closed eyes. Its main feature in adults is usually the alpha rhythm, which emerges during early childhood, increasing in frequency and responsive-ness. Clinically, relaxed wakefulness reflects a par-ticularly important state of controlled attention and arousal, and is easily and reliably reproduced in a wide range of patient groups and ages. The corres-ponding EEG has repeatedly been shown to dis-criminate between patients and controls. Long-term stability (5 years) of the individual frequency-power profiles in eight-year-old healthy children (Woerner, Rothenberger, & Lahnert, 1987) indicates that these profiles may be useful for genetic studies on con-trolled attention and arousal.
The EEG can measure brain function not only across a wide range of maturational and develop-mental states, but also during both wakefulness and sleep. As an essential indicator of sleep stages and cycles, the EEG provides unique information regarding altered sleep physiology in psychiatric disorders specific to certain sleep stages such as the disturbance of rapid eye movement (REM) sleep in depression, which is associated with metabolic increases in paralimbic and prefrontal networks (Nofzinger et al., 2004). Sleep structure may also be affected in children with ADHD, with an increased duration of the absolute REM sleep and increased number of sleep cycles (Kirov, Pillar, & Rothenberger, 2004). Since spectral and topographic sleep EEG features in adults also show striking intraindividual stability (Buckelmu¨ ller, Landolt, Stassen, & Acher-mann, 2006; Finelli, AcherAcher-mann, & Borbely, 2001), these measures may help to search for genes under-lying functional aspects of undisturbed human sleep. The typical EEG frequency analyses translate the high time resolution into a high frequency resolution. Numerous studies demonstrate that this frequency resolution is essential to characterise arousal, state regulation, and clinical features. Findings from EEG-based schizophrenia research can illustrate this point. Drug-naı¨ve first-episode adult schizophrenic patients exhibit increased slow as well as reduced medium frequency activity, with both dysfunc-tions localised in a similar frontal network (Pascual-Marqui et al., 1999). Such an increase of slow frontal EEG activity also occurs in active tasks, where it re-sults in increased variability of frontal ERPs, and has been interpreted as noise in prefrontal transmission under genetic control (Winterer et al., 2004). More refined topographic microstate analyses also reveal prominent developmental stages during adolescence (Koenig et al., 2002), and deviant state-transitions in the sub-second time range in adult patients with schizophrenia (Lehmann et al., 2005).
Because EEG frequencies characterise both development and arousal or attention, EEG studies are well suited to characterise developmental atten-tion disorders such as ADHD (attenatten-tion deficit hyperactivity disorder). Extensive EEG research has also related subtypes of ADHD to developmental
models. The spontaneous or resting EEG of ADHD children is characterised by increased slow, and de-creased fast activity (reviewed in Barry, Clarke, & Johnstone, 2003a; Brandeis, 2000). The increased slow activity is most prominent in children with hyperkinetic disorder (HD) or ADHD-combined, but is also found in ADHD children of the inattentive subtype. This slow activity has a frontocentral dis-tribution, although group differences are most prominent over posterior regions and reliably dis-criminate ADHD children from controls. This type of EEG deviation could be compatible with models implicating maturational lag1 or underarousal in
ADHD. However, at least one subtype of ADHD which is characterised by impulsivity and increased frontal beta activity does not fit this developmental lag scheme (Clarke, Barry, McCarthy, & Selikowitz, 2001). Further investigations of these subtypes in terms of their clinical utility would appear promising. The resting EEG abnormalities also show develop-mental continuity, i.e., increased slow posterior activity continues to distinguish adolescents and adults with ADHD from age-matched normal (Bres-nahan, Anderson, & Barry, 1999) and clinical con-trols (Bresnahan & Barry, 2002). They are thus hard to reconcile fully with developmental lag models.
In conclusion, EEG provides easy access to reli-able measures of normal and deviant arousal and state regulation across a wide age range including infancy, where other functional brain mapping methods are of limited use. EEG studies also clarify that clinical deficits often affect multiple EEG fre-quency bands, and thereby reflect functional deficits in several distinct networks evident at the same time. Such conclusions cannot be drawn using methods with lower time-frequency resolution. Table 1 pro-vides an overview of EEG frequency bands, their topography, developmental and functional charac-teristics, and exemplary findings discussed above.
Brain functions and applications – ERP
Automatic stimulus processing and attentional orientingA main strength of ERPs is their ability to resolve the split-second time course of information processing. ERPs have been used to determine whether incoming sensory information is selected at initial, early (la-tency range 100–250 ms) or at later (la(la-tency range >250 ms) stages of processing, i.e., before or after
1If markers of developmental psychopathology also
charac-terise an earlier stage of normal development, maturational lag becomes a plausible explanation. However, if such markers are not typical for a normal child of any age, the pattern instead suggests a deviation of development. If a marker of develop-mental psychopathology does not normalise with maturation, a developmental lag hypothesis can no longer explain the finding (Rothenberger, Banaschewski, Siniatchkin, & Hein-rich, 2003).
stimulus properties are fully analyzed. ERPs have demonstrated that the early stages of auditory and visual information processing can be altered by attention (Mangun, Hillyard, & Luck, 1992), even in young healthy children (Taylor & Khan, 2000; Yordanova, Banaschewski, Kolev, Woerner, & Ro-thenberger, 2001).
In ADHD children, already the early information processing stages related to the initial orienting and stimulus evaluation are altered (Brandeis et al., 1998; Jonkman et al., 1997; Kemner et al., 1996; Oades, 1998; Robaey, Breton, Dugas, & Renault, 1992; Steger, Imhof, Steinhausen, & Brandeis, 2000; van Leeuwen et al., 1998; Yordanova et al., 2000).
Brain mapping indicates that children with ADHD exhibit increased early automatic attentional ori-enting (increased N1) before failing to allocate suffi-cient attentional resources in further processing stages (reduced P300, central processing, motor output) (Brandeis et al., 2002a). This finding con-verges with shorter latencies of early auditory ERPs around 100 ms (Oades, 1998; Satterfield, Schell, & Nicholas, 1994), and with a deviant topography of the visual N1 at around 200 ms in a stop task2 in children with ADHD (Brandeis et al., 1998; Pliszka,
Liotti, & Woldorff, 2000); the latter results also indicate that a failure in early orienting and prepar-atory mechanisms can precede and partly determine subsequent processing.
In adult schizophrenia, abnormal early sensory processing is reflected by less refractory reduction (less habituation) of the P50 amplitude of auditory ERP (Myles-Worsley, Ord, Blailes, Ngiralmau, & Freedman, 2004). A recent meta-analysis confirmed that the P50 effect in schizophrenia has a large effect size (Cohen’s d¼1.5), similar to the most robust findings reported in neuroimaging and neuro-psychology in schizophrenia (for a meta-analysis see: Bramon, Rabe-Hesketh, Sham, Murray, & Frangou, 2004). Correspondingly, there is electro-physiological evidence for reduced latent inhibition in adult schizophrenic patients (increased N100 amplitudes to irrelevant stimuli), indicating a deficit in learning to ignore irrelevant stimuli (Kathmann, von Recum, Haag, & Engel, 2000).
Furthermore, ERP studies indicate that adult patients with schizophrenia are profoundly impaired in the ability to recognise complete objects based on fragmentary information, a process termed percep-tual closure, as reflected by a strongly reduced visual P100 (Doniger, Foxe, Murray, Higgins, & Javitt, 2002).
Recordings of auditory evoked potentials found difficulties in anorectic patients modulating auditory
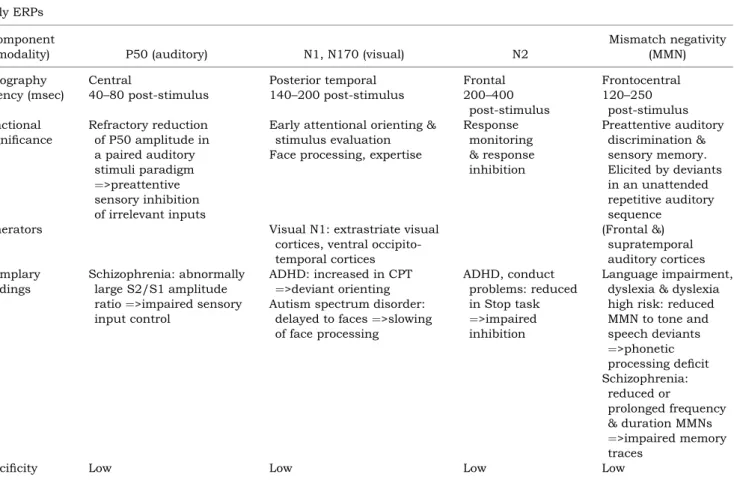
Table 2a Overview of exemplary early ERP components, their topography, developmental latency, functional significance, gener-ators, corresponding exemplary findings and their specificity
Early ERPs Component
(modality) P50 (auditory) N1, N170 (visual) N2
Mismatch negativity (MMN)
Topography Central Posterior temporal Frontal Frontocentral
Latency (msec) 40–80 post-stimulus 140–200 post-stimulus 200–400
post-stimulus 120–250 post-stimulus Functional significance Refractory reduction of P50 amplitude in a paired auditory stimuli paradigm ¼>preattentive sensory inhibition of irrelevant inputs
Early attentional orienting & stimulus evaluation Face processing, expertise
Response monitoring & response inhibition Preattentive auditory discrimination & sensory memory. Elicited by deviants in an unattended repetitive auditory sequence
Generators Visual N1: extrastriate visual
cortices, ventral occipito-temporal cortices (Frontal &) supratemporal auditory cortices Exemplary findings Schizophrenia: abnormally large S2/S1 amplitude
ratio¼>impaired sensory
input control
ADHD: increased in CPT
¼>deviant orienting
Autism spectrum disorder:
delayed to faces¼>slowing
of face processing ADHD, conduct problems: reduced in Stop task ¼>impaired inhibition Language impairment, dyslexia & dyslexia high risk: reduced MMN to tone and speech deviants ¼>phonetic processing deficit Schizophrenia: reduced or prolonged frequency & duration MMNs ¼>impaired memory traces
Specificity Low Low Low Low
2The Stop task is a reaction time task where Stop signals
oc-casionally follow the Go signals and require inhibition of on-going responses.
Tab le 2b Ove rview of exem plary lat e ER P compo nents, the ir topogr aphy, de velopm ental lat ency, func tional sign ificanc e, generators, corresp onding exemp la ry findings and their spe-cifici ty Late ERPs Co mpon ent (mod ality) E rror-rel ated negat ivity (ERN) P300 R eadine ss poten tial (RP ); lateral ised RP (LRP) Co ntinge nt negative vari ation (CN V) N400 Topog raphy Fron toc entral P3a, No go-P3: fron tocent ral; Cu e-P3, oddbal l P3: cent ropari etal Ini tial bilater ally cent rally sym metric, the n lateral ised over motor cortex Ce ntropari etal Laten cy (msec) 100 after error 300–8 00 post-st imulu s RP start s 1000, LRP 200–5 00 prior to mov ement 400 post-stim ulus Functi onal significa nce E rror evalu ation & co nflict mon itoring Sensi tive to mood a nd pe rson ality varia bles Odd ball-P3 , Cue P3: attenti onal allo cation, stim ulus evalu ation & cont ext u pdatin g No go-P30 0 ampl itud e, a nterio risation: respon se control P3a: novel ty orienting Motor prep aration Cogniti ve prep aration , time est imatio n and workin g me mory Sem antic langu a ge processing , cont extual integr ation Generat ors A CC; modul ated by DLP FC & dopami ne No goP3: ACC, DLP FC, parie tal cortic es cue P30 0: poste rior attent ion netwo rks P3a: inf erior parie tal & prefro ntal regi ons; reflects ph asic noradr energic activi ty SM A, motor cortic es SMA, moto r cort ices Ant erior me dial tempor al lobes Exem plary findings A DHD, schi zophr enia: redu ced ER N ¼ >att enuated ACC a ctivity and reduced co ntrol; A DHD: dim inished sensi tivity to inter nal feedb ack, enha nced sensi tivity to ext ernal nega tive feedba ck ADHD , schi zophr enia: redu ced target P300 ampl itude a nd No go-P30 0 ante riori sation pu re ADHD: reduced cue P3 ¼ >att ention al prob lems ADHD + condu ct prob lems: more prono unced de ficits in Nogo P3 ¼ >inhi bitory prob lems Alcoh ol & subst ance abu se: Schi zophr enia: oddbal l P300 topogr aph y with right-sh ift ¼ left hem isphe re attenu ation P3 asso ciated wi th: quant itative tra it loci at chromosome 2, 5, 6, 17 and wit h DRD2, COM T, CHRM 2 polym orphi sms ADHD : LRP attenu a tions; Pati ents wi th simp le tics devian t RP & other motor poten tials ADHD : redu ced CNV Ap hasia: attenu ated/ de layed N400 Speci ficity Lo w No ne (exce pt topogra phy) Lo w Low Lo w
stimuli adequately at the subcortical level during starvation, indicating dysfunctions at a subcortical level (brainstem) and a transitory functional un-coupling of cortical versus subcortical neuronal systems; these deficits persisted even after gaining weight (Miyamoto, Sakuma, Kumagai, Ichikawa, & Koizumi, 1992; Rothenberger, Blanz, & Lehmkuhl, 1991; Rothenberger, Dumais-Huber, Moll, & Woerner 1995a).
In summary, ERP studies demonstrate that fast perceptual and attentional functions that occur within the initial 150 ms of information processing are affected in ADHD, schizophrenia and anorexia nervosa, even though different early processing de-ficits characterise these three disorders. The exam-ples illustrate that the time resolution of EEG/ERPs is essential to clarify how altered activation in sen-sory systems (which is also found in fMRI studies, e.g., for adolescents with ADHD, Rubia, Smith, Brammer, Toone, & Taylor, 2005) reflects a primary information processing problem, and not a second-ary, late consequence of altered cognitive or emo-tional evaluation.
Executive functions, attention, inhibition, motor processes and motivation
Later attentional and executive functions can be measured through the P300 type components fol-lowing targets in numerous tasks. The P300 com-ponents have been linked to attentional allocation, stimulus evaluation and context updating processes of working memory (Polich & Herbst, 2000). Reduced P300 amplitudes are a robust finding in a variety of psychiatric disorders in adults, including schizo-phrenia (Bramon et al., 2004), increased risk for al-cohol and substance abuse (Carlson, Katsanis, Iacono, & Mertz, 1999; Porjesz et al., 2005). In chil-dren similar P300 reductions are present in chilchil-dren at risk for schizophrenia (Schreiber, Stolz-Born, Kornhuber, & Born, 1992), and in ADHD (Brandeis et al., 2002a; reviewed in: Barry, Johnstone, & Clarke, 2003b; Brandeis, 2000; Klorman, 1991), as well as in a number of other disorders. P300 attenuation is usually measured for correctly detec-ted targets only and is therefore not just a correlate of poor performance. While the much-replicated P300 attenuation in schizophrenia is not disorder-specific, the right-shift of the P300 topography due to a left-sided amplitude reduction (found in adult patients) appears rather specific, and correlated with left temporal gray matter loss (McCarley et al., 2002; Salisbury et al., 1998; Strik, Fallgatter, Stoeber, Franzek, & Beckmann, 1996a).
The specific brain functions underlying inatten-tion, impulsivity and impaired response control can be separated with adequate ERP tasks, for example the cued continuous performance test (CPT A-X) which yields different P300 components to cue, tar-get, and NoGo stimuli.
In ADHD children, impaired attention is reflected in a reduction of the P300 to cues which signal that the next stimulus may be a target. This finding (van Leeuwen et al., 1998) was replicated in multicentre studies (Banaschewski et al., 2003; Brandeis et al., 2002a; van Leeuwen et al., 1998). Importantly, these attentional deficits occur without concomitant re-sponses or performance deficits, temporally precede inhibitory or executive control, predict subsequent performance (Banaschewski et al., 2003; van Leeu-wen et al., 1998), and do not depend on age and sex (Brandeis et al., 2002a). These covert attention de-ficits are more pronounced in ADHD children with-out than in those with comorbid externalising behaviour problems, despite a more severe psycho-pathology in the latter group (Albrecht, Banaschew-ski, Brandeis, Heinrich, & Rothenberger, 2005; Banaschewski et al., 2003). Electrophysiological evidence thus supports conclusions from recent family studies that the comorbid condition repre-sents a separate pathological entity as considered in the ICD-10 classification system, rather than a sum of deficits from both pure disorders (Faraone, Biederman, Mennin, Russell, & Tsuang, 1998).
Tomographic source solutions converge to poster-ior cue P300 sources (Brandeis et al., 2002a; Herr-mann & Fallgatter, 2004; van Leeuwen et al., 1998). This finding contrasts with the predominance of frontal deficits in metabolic studies of ADHD, and suggests under-activation of the posterior attention system in ADHD children, and an involvement of central noradrenergic networks as the cue P300 is substantially modulated by this transmitter system. Further deficits of ADHD children during beha-viourally silent waiting and preparation periods are implicated by reduced amplitudes of the contingent negative variation (CNV) component (Dumais-Huber & Rothenberger, 1992; Hennighausen, Schulte-Ko¨rne, Warnke, & Remschmidt, 2000; Perchet, Re-vol, Fourneret, Mauguiere, & Garcia-Larrea, 2001). This component has been related to several brain processes implicated in ADHD, such as selective preparation, time estimation (Elbert, Ulrich, Rock-stroh, & Lutzenberger, 1991; Pouthas, Garnero, Ferrandez, & Renault, 2000) and working memory (McEvoy, Smith, & Gevins, 1998). While specific motor preparation (LRP) is also reduced in children with ADHD (Steger et al., 2001), the core deficits (e.g., attenuated P3) were shown not to be time-locked to motor output processing stages, but rather to increase with central factors such as coordina-tion demands (Steger et al., 2001; Steger et al., 2000).
Inhibitory control deficits as reflected by reduced Nogo P300 are also found in children with ADHD (Brandeis, van Leeuwen, Steger, Imhof, & Stein-hausen, 2002b; Fallgatter et al., 2004; Rubia, Oos-terlaan, Sergeant, Brandeis, & v Leeuwen, 1998). However, these are preceded by state regulation deficits (Brandeis et al., 1998; Pliszka et al., 2000) or
accompanied by executive control deficits, particu-larly at slow event rates (Wiersema, van der Meere, Roeyers, Van Coster, & Baeyens, 2006). All these findings implicate a more general state or response regulation problem in ADHD (Banaschewski et al., 2004). Unlike the covert attentional deficits, re-sponse control deficits are more prominent in chil-dren with comorbid externalising disorders.
Consistent with this suggestion, most ERP studies have found that children with ADHD have a normal enlargement of their N2 component in Nogo as com-pared to Go trials during GoNogo tasks (Ba-naschewski et al., 2004; Fallgatter et al., 2004; Fallgatter, 2001; Overtoom et al., 1998, 2002; Wier-sema et al., 2006); interestingly, this enlargement has specifically been associated with inhibition (Eimer, 1993; Falkenstein, Hoormann, & Hohnsbein, 1999; Jodo & Kayama, 1992; Kok, 1986; Pfefferbaum, Ford, Weller, & Kopell, 1985; Sasaki, Gemba, & Tsujimoto, 1989; Sasaki et al., 1996; Schroger, 1993).
In contrast, the subsequent Nogo-P300 and its an-teriorisation (a topographic measure of how much it is anterior to the go-P300) are reduced in children (Brandeis et al., 2002b; Fallgatter et al., 2004) and adults with ADHD (Fallgatter et al., 2005). This Nogo-P300 seems to reflect more general processes of re-sponse and conflict control beyond inhibition, and seems to be related to activations of the anterior cin-gulate cortex, and additional frontal and parietal regions in adults (e.g., Carter et al., 2000; Falkenstein et al., 1999; Fallgatter et al., 2001, 2004). Similarly, both attentional and inhibitory deficits are seen for ADHD children performing the Stop task, as their reduced activity to Go-signals precedes an attenuated right frontal N2-activity to Stop-signals (Albrecht et al., 2005; Brandeis et al., 1998; Pliszka et al., 2000). Control deficits in psychiatric disorders are also evident from altered ERP activity to performance errors and feedback. The initial error related frontocentral negativity (ERN) peaks 100 ms after motor onset. It has been associated with perform-ance monitoring, conflict inhibition and error pro-cessing (for a review in adults: Falkenstein, Hoormann, Christ, & Hohnsbein, 2000), localised to the anterior cingulate gyrus (ACC), and is modulated by lateral prefrontal cortex and the dopaminergic system (e.g., Carter et al., 2000; Dehaene, Posner, & Tucker, 1994). The development of conflict and error processing is reflected in major differences between children, adolescents and adults regarding the time course of concomitant brain activation (Davies, Se-galowitz, & Gavin, 2004; Hogan, Vargha-Khadem, Kirkham, & Baldeweg, 2005; Rueda, Posner, Roth-bart, & Davis-Stober, 2004).
The ERN amplitude is sensitive to mood and per-sonality variables (Luu, Collins, & Tucker, 2000), especially when correct responses are rewarded and/or incorrect responses are punished (Dikman & Allen, 2000), suggesting that affective and motiva-tional processes may significantly influence
pro-cesses of performance monitoring and conflict processing. Consistent with this, frontocentral negativities with ACC sources are also involved in processing feedback signals during guessing and gambling in adults (Gehring & Willoughby, 2002; Nieuwenhuis, Holroyd, Mol, & Coles, 2004).
In children with ADHD, one study found the ERN during STOP task performance to be markedly re-duced, suggesting a deficit in cognitive control pro-cesses (Liotti, Pliszka, Perez, Kothmann, & Woldorff, 2005). However, another study indicated that chil-dren with ADHD may have a normal ERN, but may suffer from abnormal response strategy adjustments, reflected in deviant ERP activity following the ERN (diminished error positivity), suggesting reduced evaluation of the error (Wiersema, van der Meere, & Roeyers, 2005). Studies recording ERP associated with feedback processing (feedback-related negativ-ity) in children with ADHD found diminished sensit-ivity to internal feedback (internal error signals, predicting the likelihood of an error) during a time production task (van Meel, Heslenfeld, Oosterlaan, Luman, & Sergeant, 2005a), suggesting problems in assigning relative motivational significance to negat-ive outcomes rather than in differentiating between favourable and unfavourable outcomes. In contrast, children with ADHD showed an enhanced sensitivity to external negative feedback during a guessing paradigm despite similar behavioural responses (van Meel, Oosterlaan, Heslenfeld, & Sergeant, 2005b). An enhanced ERN in patients with obsessive-compulsive disorder (OCD) suggests that the clinical features in OCD are related to hyper-functioning error and ac-tion-monitoring processes (Hajcak & Simons 2002; Johannes et al., 2001).
In schizophrenia, ERP studies in adults have documented diminished frontal response control through reduced Nogo anteriorisation (Fallgatter & Muller, 2001) and deviant neural processes involved in response inhibition (Kiehl, Smith, Hare, & Liddle, 2000). Correlations of Nogo-P300 and fMRI data suggest that adult patients with schizophrenia and healthy subjects use different neural structures to inhibit responses (Ford et al., 2004). Auditory P300 amplitude abnormalities in adults have indicated high-level cognitive dysfunctions that do not origin-ate from potential preceding impairments at lower information processing levels (van der Stelt, Frye, Lieberman, & Belger, 2004; van der Stelt, Lieber-man, & Belger, 2005).
Several child psychiatric disorders are also char-acterised by movement problems (e.g., ADHD, tic disorders, autistic stereotypies, developmental co-ordination disorder). Therefore, the neural dynamics of normal and pathological movement in the develop-ing child are of special interest, and may even have clinical potential for training of deviant movement regulation in the light of recent ERP evidence for rapid plasticity of motor activation in adults (Halder et al., 2005).
With ERPs, it is even possible to study the covert process of motor preparation with the readiness po-tential (RP) preceding movement onset. This slowly increasing precentral negativity peaks around the onset of a voluntary movement. Its initial bilateral phase is followed by the lateralised readiness po-tential (LRP), which is contralateral to the movement and considered to reflect the output of the response selection stage (Coles, 1989; Eimer, 1998). A con-sistently negative pre-central RP can only be regis-tered by age ten (Chiarenza, Papakostopoulos, Giordana, & Guareschi-Cazzullo, 1983). Recent developmental work combining ERP with fMRI also indicates that the basic motor network is established around age 9, and that topographic changes of motor ERPs beyond this age may result from increasing deactivation of a non-specific, task-irrelevant default network with age (Halder et al., in press). Since the RP, as a voluntary ERP, is usually missing before a tic (Hallett, 2001) and tics can be seen during all sleep stages (Rothenberger et al., 2001), these motor phenomena seem to be non-voluntary. Hence, electrophysiology is at the moment the only method which may contribute empirical evidence to further clarify the issue of voluntary vs. involuntary aspects of movements. Also, earlier RP studies first sugges-ted that patients with tic disorder may develop frontal premotor compensatory mechanisms to con-trol for their tics (Rothenberger, 1982). This assumption was recently supported indirectly by studies with MRI (Leckman, Vaccarino, Kalanithi, & Rothenberger, 2006) and is in line with the fact that the frontally increased RP normalises after success-ful treatment of the tics with neuroleptics (Rothen-berger, 1990).
ADHD children exhibit a more bilateral RP dis-tribution as well as an attenuation of both LRP and P300. This suggests parallel problems in motor preparation and attention or coordination (Rothen-berger, 1995b; Steger et al., 2000) which could not be measured by other methods.
In conclusion, specific ERP tests allow detailed mapping of the information processing sequence of attentional, inhibitory and premotor brain functions for state regulation and response control. Non-specific deficits of control functions with attenuated ACC activity characterise several disorders including comorbid ADHD + CD and schizophrenia, while deficits of premotor functions and posterior atten-tional networks are prominent in tic disorders and pure ADHD, respectively.
Perception, face processing, emotion
Neural correlates of face recognition in the occipito-temporal brain, including the bilateral fusiform and right superior temporal cortices, specialised for faces, are indexed by the N170 (Henson et al., 2003). This component can be used to assess develop-mental changes of face recognition already during
postnatal development (de Haan & Nelson, 1999; Taylor, Edmonds, McCarthy, & Allison, 2001); its recording does not require verbal responses, and it is more sensitive in assessing facial recognition in infants than the traditional behavioural measure of looking time (de Haan & Nelson, 1997).
ERP studies reveal a slowing of face processing (by 20 ms) in young children with autism spectrum disorder (ASD), as reflected in N170 latency (McPartland, Dawson, Webb, Panagiotides, & Car-ver, 2004). These children’s ERP amplitudes also fail to differentiate familiar from unfamiliar faces, although they distinguish familiar from unfamiliar objects, suggesting that autism is associated with face recognition impairment early in life (Dawson et al., 2002). Furthermore, 3–4-year-old children with ASD show a disordered pattern of neural re-sponses to emotional stimuli (Dawson, Webb, Carver, Panagiotides, & McPartland, 2004).
In contrast, fMRI studies failed to find consistent differences between adults with autism spectrum disorder and normal controls in fusiform gyrus ac-tivation during face processing (Hadjikhani et al., 2004; Hubl et al., 2003).
This example does indicate that structure and overall activation can be basically intact, even though the timing of the processes involved may be impaired; thus, measuring brain functions with a high time resolution can be crucial. Early interven-tions that enhance social attention should result in changes in brain activity that are reflected in ERP to face stimuli, and those children showing the greatest social improvement should exhibit more normal brain activity.
ERP indices confirm the presence of an attentional bias for negative stimuli in anxiety, as high-anxious adults show significantly larger amplitudes in the early CNV during vigilance towards negative stimuli (Carretie, Mercado, Hinojosa, Martin-Loeches, & Sotillo, 2004). In contrast to behavioural data, the ERP data indicate that threat-related faces elicit faster and stronger early ERP responses in high-anxious than in low-high-anxious adults (Bar-Haim, Lamy, & Glickman, 2005).
Anxious children also show higher ERP ampli-tudes than their controls, probably reflecting diffi-culties in the effortful regulation of negative emotion (Lewis & Stieben, 2004; Lewis, Lamm, Segalowitz, Stieben, & Zelazo, 2006). Correspondingly, atypical EEG patterns of frontal brain activation have been found in anxious school children (Baving, Laucht, & Schmidt, 2002). These findings indicate that EEG studies may contribute to the understanding of the development and the neuronal underpinnings of emotion regulation in children.
Sensory memory and language
Auditory memory traces are measured through the MMN, a frontally negative ERP component with a
latency of 120 to 250 ms that is elicited by deviations of sound in a repetitive auditory sequence. It can be considered as an outcome of an automatic ‘compar-ison process’ that contrasts incoming auditory input against the memory traces of the preceding sounds, and it is a valuable tool for investigating very early pre-attentive auditory discrimination and sensory or ‘echoic’ memory even in infants and children (e.g., Cheour, Leppanen, & Kraus, 2000; Na¨a¨ta¨nen, 1990). Auditory ERPs have been used to study mechan-isms of early language acquisition (Baldeweg, Rich-ardson, Watkins, Foale, & Gruzelier, 1999; Leppa¨nen & Lyytinen, 1997). Thus, the development of language-specific memory traces has been de-monstrated by the age of 1 year (Cheour et al., 1998), although considerable topographic ERP differences remain between children and adults (Maurer, Bu-cher, Brem, & Brandeis, 2003b). Auditory ERPs have been applied in the investigation of specific language impairment and related disorders, and for identify-ing children at risk, as they may reveal immaturity or deficits of auditory processing even when beha-vioural thresholds look normal (Bishop & McArthur, 2005).
In developmental dyslexia, ERP studies docu-mented that neurophysiological correlates of word recognition are significantly attenuated (Brandeis et al., 1994; Schulte-Ko¨rne, Deimel, Bartling, & Re-mschmidt, 2004). Abnormalities of the MMN corre-lated with the degree of phonological impairment (Baldeweg et al., 1999; Kraus et al., 1996), pointing to a selective processing deficit at an earlier phonetic level as a possible source of the difficulties in learn-ing to read, while at a later lexical level information seems to be processed normally (Bonte & Blomert, 2004). Several groups studied auditory ERPs to speech sounds in infants with and without a genetic risk for dyslexia. All of them found significant group differences, although the ERP deviance varied with the speech test and the infants’ age. Infants at risk had larger responses to standard sounds (more right-central negativity after 400 ms) by the age of 6 months (Pihko et al., 1999), larger response (more right frontal positivity after 300 ms) to deviants by the age of 2 weeks (Leppa¨nen et al., 1999), and essentially absent mismatch responses to subtle speech contrasts by the age of 2 months (van Leeu-wen et al., 2006). Children at risk for dyslexia con-tinue to show altered mismatch responses to subtle tone and speech deviants when they start learning to read, with a less left lateralised mismatch response to speech stimuli implicating phonological deficits (Maurer, Brem, Bucher, & Brandeis, 2003a). Lon-gitudinal studies confirm that infants’ ERPs to speech sounds also predict (in a statistical sense) their subsequent language development, such as verbal memory at age 5 (Guttorm et al., 2005) or reading skills at age 8 (Molfese, 2000).
Although MMN abnormalities indicative of auditory processing deficits have also been reported
in various other disorders (for reviews: Cheour et al., 2000; Na¨a¨ta¨nen & Escera, 2000), the underlying sensory memory impairments may be specific for a given disorder. Thus, dyslexics exhibit MMN attenuations mainly to subtle speech and frequency deviance (Baldeweg et al., 1999), while schizophrenic patients, or those at high risk for schizophrenia due to microdeletion in the 22q11.2 gene, have reduced MMNs to large frequency and especially duration deviants (adolescents: Baker, Baldeweg, Sivagnanasundaram, Scambler, & Skuse, 2005; adults: Javitt, Grochowski, Shelley, & Ritter, 1998; Michie et al., 2000; Oades, Dittmann-Balcar, Zerbin, & Grzella, 1997), suggesting differ-ent deficits in preattdiffer-entive auditory processing in schizophrenic patients and individuals with dyslexia.
The processing of language at the semantic and syntactic level is indexed by several ERP compon-ents. The N400 is considered to be a sensitive index of semantic processing reflecting neural mechan-isms of semantic integration into context. It has been localised to the anterior medial temporal lobe (McCarthy, Nobre, Bentin, & Spencer, 1995). ERP data indicate that both lexical expectations facil-itating early phonological processing and mechan-isms of semantic priming facilitating integration into semantic context are already present in 19-month-olds (Friedrich & Friederici, 2004). Syntactic viola-tions elicit an early left anterior negativity and a late centroparietal positivity called P600 in 3-year-old children (for a review see: Friederici, 2005).
These findings demonstrate that ERP studies (in contrast to fMRI) are particularly well suited to track very early language development, and to detect language processing deficits ranging from simple automatic sensory memory to high-level syntactic functions.
Brain functions and applications – single trial
analyses
Intraindividual variability – ADHD
Although computationally demanding, single-trial analyses are important to investigate the degree of trial-to-trial variability that may characterise differ-ent clinical syndromes. A single-trial analysis of low frequency ERP components (Heinrich et al., 2001) suggested that children with ADHD allocate more attentional resources for an adequate performance with time-on-task, supporting a sustained attention deficit in ADHD, which is not consistently found using neuropsychological tasks (van der Meere, 1996; van Leeuwen et al., 1998). Moreover, large variability in component amplitudes and latencies between single-trial ERP, suggestive of greater variability in the extent and timing of latent cognitive processing stages, has been found in ADHD (Lazzaro et al., 1997), matching reaction time results
(Drechsler, Brandeis, Foldenyi, Imhof, & Steinhau-sen, 2005; Kuntsi & Stevenson, 2001).
Outlook
EEG/ERP parameter – potential predictors of treatment response?
The intensity (i.e., loudness)-dependent amplitude change of auditory evoked ERP has been suggested as an indicator of the level of central serotonergic neurotransmission in adults (Hegerl, Gallinat, & Juckel, 2001; Strobel et al., 2003). This parameter has potential clinical value since subgroups of pa-tients with a serotonergic dysfunction can be iden-tified and can be treated more specifically. In depressed patients, a significant relationship be-tween strong LDAEP, indicating low serotonergic function, and a favourable response to SSRI has been found. Additionally, there is evidence that the LDAEP is a predictor of favourable response to a preventive lithium treatment in adult patients with affective disorders, and of responsiveness to rebo-xetine treatment in major depression (Hegerl et al., 2001; Linka, Muller, Bender, Sartory, & Gastpar, 2005). In adult schizophrenic patients, a prospective longitudinal study has demonstrated the predictive value of auditory P300-amplitudes on clinical out-come in terms of social functioning (Strik, Fallgatter, Stoeber, Franzek, & Beckmann, 1996b).
Electrophysiological measures also reflect changes in the pattern of brain activations due to training. Short-term attention training had a specific effect on the scalp distribution of the ERPs that resembled the effect of maturation, thus supporting the direction of the behavioural data showing more adult-like per-formance after training (Rueda, Rothbart, Mc-Candliss, Saccomanno, & Posner, 2005). Positive behavioural and specific neurophysiological effects of neurofeedback training of slow cortical potentials has been demonstrated in children with ADHD (Heinrich, Gevensleben, Freisleder, Moll, & Rothen-berger, 2004).
Future studies will have to show whether objective and reliable tools like ERP combined with genetic markers can be used to predict individual patients’ treatment response even better.
EEG/ERP parameter – potential endophenotypes for multilevel investigations of etiological pathways?
Many psychiatric disorders are probably best con-ceptualised as clinically and etiologically hetero-genous conditions with differentiable pathways linking specific putative causes to clinical pheno-types (Banaschewski et al., 2005). The still limited knowledge of etiological and pathogenic mechanisms is a major barrier for the development of selective treatments for specific subgroups of patients. Efforts to clarify the disorders’ molecular genetic basis have
often been hindered by an inability to detect non-clinically penetrant carriers of the predisposing genes and by uncertainties concerning the nature of the non-genetic influences and the extent of locus heterogeneity.
Thus, ideally, molecular genetic studies should not be performed on psychiatric diagnostic categor-ies alone, but also on quantitative neurobiological measures or markers (i.e., intermediate phenotypes, or endophenotypes) of the genetic risk to develop psychiatric disorders which are also associated with the psychopathological symptoms (Castellanos & Tannock; 2002; de Geus, 2002; Gottesman & Shields, 1973; Tsuang & Faraone, 2000). This endophenotype approach is an alternative method for disentangling phenotypic variation that may facilitate the identification and functional charac-terisation of susceptibility genes and other etiologic factors in etiologically complex disorders, because endophenotypes are less complex and more proximal to gene function than either diagnostic categories or neuropsychological measures (Almasy, 2003). This approach has the advantage of providing insight into the underlying pathogenic mechanisms and can help to disentangle the pathways through which genes may exert their influences on neuropsychological parameters and behavioural symptoms.
Quantitative electrophysiological markers of human information processing and psychopathology are thus reliably detected throughout childhood and adolescence, and differentiate between cases, high-risk relatives and controls. Furthermore, they show a higher heritability than neuropsychological vari-ables, suggesting that they may provide potentially useful endophenotypes. Unfortunately, heritability of fMRI measures has still to be determined; a recent study found that prefrontal cortex activation is sen-sitive to genetic vulnerability for ADHD (Durston, Mulder, Casey, Ziermans, & van Engeland, in press). Considerable heritability for EEG band power has been found for both slow and fast activities in both 5-year-old children (averaging .81 for theta and .73 for beta) and adults (averaging .89 for theta and .86 for beta). For ERP, the estimated mean heritability is 60% for P300 amplitude and 51% for P300 latency, depending to some extent on task conditions (van Beijsterveldt & van Baal, 2002). A substantial pro-portion of genetic influences on P300 amplitude may be explained by strong heritability of theta and delta oscillations elicited during cognitive processing of stimuli contributing to P300 (Anokhin et al., 2001; Yordanova & Kolev, 1996). Functionally, P300 ac-tivity in adults has been associated with quantitative trait loci at chromosomes 2, 5, 6 and 17 (Porjesz et al., 2005) and with specific genes involved in dopamine transmission (DRD2 polymorphism, Hill et al., 1998; COMT polymorphism, Gallinat et al., 2003).
The utility of quantitative electrophysiological measures as endophenotypes of disinhibitory
dis-orders, including alcoholism, has recently been illustrated in adults by the Collaborative Study on the Genetics of Alcoholism. The association between GABA-A receptor gene GABRA2 (the gene encoding the alpha2 subunit), EEG beta frequency and the risk for alcohol dependence in adults (Covault, Gel-ernter, Hesselbrock, Nellissery, & Kranzler, 2004; Edenberg et al., 2004) suggests that this gene affects the predisposition to develop alcohol dependence through influences on the level of neural excitability. Furthermore, a cholinergic muscarinic receptor gene on chromosome 7 (CHRM2) was shown to be related to the theta and delta event-related oscillations underlying P300 to target stimuli and also to the clinical diagnoses of alcoholism and depression (Jones et al., 2004; Wang et al., 2004).
For ADHD, such specific pathophysiological pathways have still to be identified. Taken together, ERP findings indicate that children with ADHD suf-fer from a more general regulation problem, includ-ing deficits of attentional orientinclud-ing and response preparation and response monitoring, than just from a response inhibitory deficit causing secondary def-icits in other executive functions as proposed by Barkley (1998). Hence, ERP could differentiate the cognitive-motor problem best, compared to neuro-psychology and other methods. Further, ERP para-digms have differentiated children with ADHD with and without comorbid conduct disorder or tic dis-order (Banaschewski et al., 2003, 2004; Rothenber-ger et al., 2000; Yordanova, Dumais Huber, Rothenberger, & Woerner, 1997; Yordanova, Hein-rich, Kolev, & Rothenberger, in press), indicating that the presence of comorbidity in ADHD may alter brain electrical correlates specifically, without ne-cessarily affecting the level of overt behavioural performance. Meanwhile, the idea of multiple devel-opmental pathways to ADHD has been articulated and developed in several recent theory papers (Banaschewski et al., 2005; Nigg, Blaskey, Stawicki, & Sachek, 2004a; Nigg, Goldsmith, & Sachek, 2004b; Sonuga-Barke, 2003).
Conclusions/summary
Electrophysiological parameters provide a non-invasive method to monitor spatio-temporal activa-tion in the brain during sensory, cognitive, affective, attentional and motor information processing as well as during state regulation in real time. They can reliably be reproduced in a wide range of patient groups and across a wide age range, even in infants, and during both wakefulness and sleep, and they can give indices of covert, pre-attentive or pre-motor processes. This makes them ideal for studies of brain function during normal and deviant child develop-ment. Their excellent time-frequency resolution has proven particularly useful to clarify whether multiple deficits follow and cause each other, or are present
simultaneously, which is not possible with slower metabolic methods. At the same time, the advances in source resolution now allow genuine dynamic neuroimaging with EEG/ERP.
Over the past two decades, the functional signific-ance of many EEG and ERP parameters has become biologically more meaningful and better grounded in neurosciences. EEG/ERP research has contributed substantially to the understanding of brain proces-ses and functions in normal development and their deviations in child and adolescent psychiatric con-ditions. They have helped to clarify whether overt task performance reflects the same underlying pro-cesses throughout development and in various dis-orders (e.g., response inhibition in schizophrenia) and to illuminate the content validity of neuro-psychological tests by revealing the modular archi-tecture of more complex neuropsychological constructs (e.g., inhibition, attention), to test alter-native models of information processing (e.g., all-or-none or discrete stage models3 vs. continuous models) and to constrain psychological theories.
EEG/ERP research on ADHD, indicating the occurrence of a sequence of multiple activation def-icits of posterior and anterior attentional networks within a subsecond range, may serve as an illustra-tion; attentional focusing and orienting deficits, temporally and causally preceding inhibitory or executive control, were found to predict subsequent performance during CPT-AX performance.
However, the issues of specificity of findings associated with certain disorders and the impact of comorbid disorders still needs to be addressed. Few EEG and ERP studies have investigated the impact of comorbid disorders using a full factorial design, to allow separation of effects of comorbid disorders and their interaction (for an exception see: Banaschewski et al., 2003, 2004), and some of the most reliable deficits are not specific for one disorder. In addition, only few EEG- or ERP-based deficits have been rep-licated across a wide age range. Studies on children and adolescents are still largely lacking. Given these limitations, the clinical use of EEG/ERP as a tool for individual diagnoses of child psychiatric disorders is clearly not warranted at this stage.
Because some EEG/ERP measures are substan-tially heritable and developmentally stable, they are particularly promising endophenotype candidates that may disentangle phenotypic variation (e.g., ADHD with vs. without comorbid externalising behaviour) and facilitate the identification and functional characterisation of susceptibility genes and other etiologic factors in etiologically complex disorders. This may help to pave the way for the
3
All-or-none or discrete stage models of information proces-sing assume – in contrast to continuous models – that partially processed information cannot be used by further processing stages, respectively that information processing on a later stage cannot start until a prior stage is completed.
identification of prognostic predictors for (psycho-pathological) developmental processes and the de-sign of selective treatments for specific diagnostic subgroups of patients and families.
Correspondence to
T. Banaschewski, University of Go¨ttingen, Child and Adolescent Psychiatry, von-Siebold-Str. 5, D-37075 Go¨ttingen, Germany; Tel: 0049-551-396727; Fax: 0049-551-398120; Email: [email protected]
References
Albrecht, B., Banaschewski, T., Brandeis, D., Heinrich, H., & Rothenberger, A. (2005). Response inhibition deficits in externalizing child psychiatric disorders: An ERP-study with the Stop-task. Behavioral and Brain Functions,1, 22.
Almasy, L. (2003). Quantitative risk factors as indices of alcoholism susceptibility. Annals of Medicine, 35, 337–343.
Anokhin, A.P., van Baal, G.C., van Beijsterveldt, C.E., de Geus, E.J., Grant, J., & Boomsma, D.I. (2001). Genetic correlation between the P300 event-related brain potential and the EEG power spectrum. Beha-vior Genetics,31, 545–554.
Baker, K., Baldeweg, T., Sivagnanasundaram, S., Scambler, P., & Skuse, D. (2005). COMT VAL108/ 158 MET modifies mismatch negativity and cognitive function in 22q11 deletion syndrome. Biological Psychiatry,58, 23–31.
Baldeweg, T., Richardson, A., Watkins, S., Foale, C., & Gruzelier, J. (1999). Impaired auditory frequency discrimination in dyslexia detected with mismatch evoked potentials.Annals of Neurology,45, 495–503. Banaschewski, T., Brandeis, D., Heinrich, H., Albrecht, B., Brunner, E., & Rothenberger, A. (2003). Associa-tion of ADHD and conduct disorder – brain electrical evidence for the existence of a distinct subtype. Journal of Child Psychology and Psychiatry, 44, 356–376.
Banaschewski, T., Brandeis, D., Heinrich, H., Albrecht, B., Brunner, E., & Rothenberger, A. (2004). Ques-tioning inhibitory control as the specific deficit of ADHD – evidence from brain electrical activity. Jour-nal of Neural Transmission,111, 841–864.
Banaschewski, T., Hollis, C., Oosterlaan, J., Roeyers, H., Rubia, K., Willcutt, E., & Taylor, E, (2005). Towards an understanding of unique and shared pathways in the psychopathophysiology of ADHD. Developmental Science,8, 132–140.
Bar-Haim, Y., Lamy, D., & Glickman, S. (2005). Attentional bias in anxiety: A behavioral and ERP study.Brain Cognition,59, 11–22.
Barkley, R.A. (1998). Attention-deficit hyperactivity disorder.Scientific American,279, 66–71.
Barry, R.J., Clarke, A.R., & Johnstone, S.J. (2003a). A review of electrophysiology in attention-deficit/hyper-activity disorder: I. Qualitative and quantitative electroencephalography. Clinical Neurophysiology, 114, 171–183.
Barry, R.J., Johnstone, S.J., & Clarke, A.R. (2003b). A review of electrophysiology in attention-deficit/hyper-activity disorder: II. Event-related potentials.Clinical Neurophysiology,114, 184–198.
Baving, L., Laucht, M., & Schmidt, M.H. (2002). Frontal brain activation in anxious school children.Journal of Child Psychology and Psychiatry,43, 265–274. Birbaumer, N., Elbert, T., & Canavan, A.G. (1990). Slow
potentials of the cerebral cortex and behaviour. Physiological Reviews,70, 1–41.
Bishop, D.V., & McArthur, G.M. (2005). Individual differences in auditory processing in specific lan-guage impairment: A follow-up study using event-related potentials and behavioural thresholds. Cor-tex,41, 327–341.
Bonte, M.L., & Blomert, L. (2004). Developmental dyslexia: ERP correlates of anomalous phonological processing during spoken word recognition. Brain Research. Cognitive Brain Research,21, 360–376. Bramon, E., Rabe-Hesketh, S., Sham, P., Murray, R.M.,
& Frangou, S. (2004). Meta-analysis of the P300 and P50 waveforms in schizophrenia. Schizophrenia Research,70, 315–329.
Brandeis, D. (2000). Psychophysiologie der hyperkinet-ischen Sto¨rung. In H. Steinhausen (Ed.), Hyperkine-tische Sto¨rungen im Kindes- und Jugendalter (pp. 71–89). Stuttgart: Kohlhammer.
Brandeis, D., Banaschewski, T., Baving, L., Georgiewa, P., Blanz, B., Warnke, A., Steinhausen, H.C., Rothen-berger, A., & Scheuerpflug, P. (2002a). Multicenter P300 brain mapping of impaired attention to cues in hyperkinetic children.Journal of the American Acad-emy of Child and Adolescent Psychiatry,41, 990–998. Brandeis, D., & Lehmann, D. (1986). Event-related potentials of the brain and cognitive processes: Approaches and applications. Neuropsychologia, 24, 151–168.
Brandeis, D., van Leeuwen, T.H., Rubia, K., Vitacco, D., Steger, J., Pascual-Marqui, R.D., & Steinhausen, H.C. (1998). Neuroelectric mapping reveals precursor of stop failures in children with attention deficits. Behavioural Brain Research,94, 111–125.
Brandeis, D., van Leeuwen, T.H., Steger, J., Imhof, K., & Steinhausen, H.C. (2002b). Mapping brain func-tions of ADHD children. In K. Hirata, Y. Koga, K. Nagata, & K. Yamazaki (Eds.), Recent advances in human brain mapping (vol. 1232, pp. 649–654). Amsterdam: Elsevier.
Brandeis, D., Vitacco, D., & Steinhausen, H.C. (1994). Mapping brain electric micro-states in dyslexic chil-dren during reading. Acta Paedopsychiatrica, 56, 239–247.
Brem, S., Bucher, K., Halder, P., Summers, P., Dietrich, T., Martin, E., & Brandeis, D. (2006). Evidence for developmental changes in the visual word processing network beyond adolescence.NeuroImage, 29, 822– 837.
Bresnahan, S.M., Anderson, J.W., & Barry, R.J. (1999). Age-related changes in quantitative EEG in attention-deficit/hyperactivity disorder. Biological Psychiatry, 46, 1690–1697.
Bresnahan, S.M., & Barry, R.J. (2002). Specificity of quantitative EEG analysis in adults with attention deficit hyperactivity disorder. Psychiatry Research, 112, 133–144.