Estrogen receptor-

signaling modulates epithelial barrier function
Mirjam Looijer-van Langen,2Naomi Hotte,1Levinus A. Dieleman,1Eric Albert,1Chris Mulder,2 and Karen L. Madsen1
1Division of Gastroenterology, Department of Medicine, University of Alberta, Edmonton, Alberta, Canada; and2Department of Gastroenterology and Hepatology, VU University Medical Center, Amsterdam, The Netherlands
Submitted 14 June 2010; accepted in final form 7 January 2011 Looijer-van Langen M, Hotte N, Dieleman LA, Albert E, Mulder
C, Madsen KL.Estrogen receptor-signaling modulates epithelial
bar-rier function.Am J Physiol Gastrointest Liver Physiol300: G621–G626, 2011. First published January 20, 2011; doi:10.1152/ajpgi.00274.2010.— Impaired epithelial barrier function and estrogens are recognized as factors influencing inflammatory bowel disease (IBD) pathology and disease course. Estrogen receptor- (ER) is the most abundant estrogen receptor in the colon and a complete absence of ER expression is associated with disrupted tight-junction formation and abnormal colonic architecture. The aim of this study was to determine whether ER signaling has a role in the maintenance of epithelial permeability in the colon. ERmRNA levels and colonic permeabil-ity were assessed in IL-10-deficient mice and HLA-B27 rats by RT-PCR and Ussing chambers. ERexpression and monolayer resis-tance were measured in HT-29 and T84 colonic epithelial monolayers by RT-PCR and electric cell-substrate impedance sensing. The effect of 17-estradiol and an estrogen agonist [diarylpropionitrile (DPN)] and antagonist (ICI 182780) on epithelial resistance in T84 cells was measured. Expression of ER and proinflammatory cytokines was investigated in colonic biopsies from IBD patients. Levels of ER mRNA were decreased, whereas colonic permeability was increased, in IL-10-deficient mice and HLA-B27 transgenic rats prior to the onset of colitis. T84 cells demonstrated higher resistance and in-creased levels of ER mRNA compared with HT-29 cells. 17 -estradiol and DPN induced increased epithelial resistance in T84 cells, whereas an ER blocker prevented the increased resistance. De-creased ERmRNA levels were observed in colonic biopsies from IBD patients. This study suggests a potential role for ERsignaling in the modulation of epithelial permeability and demonstrates reduced ERmRNA in animal models of colitis and colon of patients with inflammatory bowel disease.
estrogen receptor-; intestinal permeability; inflammation; inflamma-tory bowel disease
INFLAMMATORY BOWEL DISEASE (IBD) is a group of chronic,
inflammatory diseases affecting the gastrointestinal tract. Two main disease phenotypes are Crohn’s disease and ulcerative colitis. IBD is regarded as a complex disease with a multifac-torial pathophysiological basis, but the exact pathogenesis is still unclear. Genetic susceptibility, immune dysregulation driven by a response to commensal flora, triggering environ-mental factors, and impaired epithelial barrier function are recognized as influencing IBD pathology (19). Additionally, intestinal permeability has been shown to be increased in Crohn’s disease patients (27). This phenomenon is also ob-served in healthy first-degree relatives of patients, suggesting that this defect can occur independently of inflammation (16, 20, 28, 33). Furthermore, an increase in intestinal permeability has been shown to predict relapse in patients with Crohn’s
disease (3). However, whether increased gut permeability is a causative event or merely an effect of the inflammatory milieu is still unknown.
Contraceptive use, pregnancy, and hormone replacement therapy have been reported to influence the risk of developing IBD or influencing disease course (5, 8, 18). However, the role of estrogen or gender in IBD pathology remains inconclusive. Estrogen mediates its action via estrogen receptors and is involved in modulating many biological processes, including immune responses and gastrointestinal physiology (9). Two estrogen receptors have been identified: estrogen receptor-␣ (ER␣) and estrogen receptor-(ER), both of which have their own distinct physiological function and distribution (9, 22). The results of ER␣action are well known and involve classical estrogen effects, such as development of secondary sex char-acteristics and regulation of the menstrual cycle in females and sperm maturation in males (9). ERwas discovered in 1996 and its biological effects have not been completely unrav-eled (22). Whereas ER plays a minor role in mediating action in classical estrogen target tissues, a dominant role appears to exist for ERin the brain, cardiovascular system, and colon where it is expressed primarily on epithelial cells (6, 15, 21, 36).
Research has demonstrated a role for ERin animal models, in that anti-inflammatory effects of estrogen or ERagonists have been described in models of chronic intestinal inflamma-tion (14). In ERknockout mice an increase in epithelial cell proliferation, decrease of apoptosis, and decrease in cellular adhesion molecules was found. These mice also had dis-rupted tight-junction formation and abnormal colonic archi-tecture (35).
Based on these findings, we hypothesized a role for ERin the maintenance of epithelial permeability in the colon. In the present study, we demonstrate reduced ERmRNA expression and increased gut permeability to precede the onset of colitis in two animal models. Furthermore, we observed decreased ER mRNA levels in colonic biopsies from IBD patients. Finally, in vitro experiments demonstrated a clear association between ER signaling and epithelial barrier function. These data support our hypothesis that ERsignaling has a potential role in maintaining epithelial barrier function.
MATERIALS AND METHODS
Animal models of inflammation.For this study, two chronic models of intestinal inflammation were examined. HLA-B27 transgenic (TG) rats spontaneously develop colitis and systemic inflammation in a specific pathogen-free (SPF) environment (14). TG rats and their healthy nontransgenic littermates (NTG) were maintained under SPF conditions with free access to food (LabDiet rodent diet 5001) and water. Rats were maintained at 23°C with a 12:12-h light-dark cycle. A second model of inflammation studied was the interleukin-10 (IL-10⫺/⫺) gene-deficient mouse bred on the 129 Sv/Ev background.
Address for reprint requests and other correspondence: K. L. Madsen, Univ. of Alberta, 7-142 Katz Group Center, Edmonton, AB, Canada T6G 2E1 (e-mail: [email protected]).
First published January 20, 2011; doi:10.1152/ajpgi.00274.2010.
by 10.220.33.6 on April 13, 2017
http://ajpgi.physiology.org/
These mice develop a patchy inflammation primarily in the colon, beginning at 6 – 8 wk of age (26). IL-10⫺/⫺and wild-type 129 Sv/Ev controls (wild type) were housed under SPF conditions and fed a standard diet consisting of 9% fat rodent blocks. All experiments were approved by the University of Alberta Health Sciences Animal Policy and Welfare Committee.
Measurements in animal models. At death, small intestinal and colonic tissue were removed, and total RNA was isolated in Trizol (Invitrogen, Burlington, ON, Canada) using the RNA easy kit (Qia-gen, Mississauga, ON, Canada) in accordance with the manufacturer’s instructions. RNA concentrations and purity were determined using the nd-1000 spectrophotometer (NanoDrop Technologies Wilming-ton, DE). For reverse transcription, mastermix was produced contain-ing 1l random primers, 2l DNTP mix, 9.5l RNAse free H2O, 4l 5⫻first-strand buffer, 0.5l RNAse out, 2l 0.1M DTT, 1l Superscript II Reverse Transcriptase (Invitrogen), and 1 g RNA. Real-time PCR analysis was performed using the HT-7900 sequence detector (Applied Biosystems, Foster City, CA) with GAPDH used as the endogenous control. The mouse, rat, and human ER target primers were purchased as ready-to-use inventoried gene expression assays from TaqMan (Applied Biosystems, Streetville, ON, Canada). Analysis was performed using the Sequence Detection Software
version 2.3, which is supplied with the Applied Biosystems HT-7900 instrument.
Intestinal permeability measurement.At death, segments of colon and small intestine were removed and mounted in lucite Ussing chambers for assessment of intestinal permeability. Serosal and mu-cosal surfaces were exposed to 10 ml Ringers buffer (in mmol/l: 115 NaCl, 8 KCl, 1.25 CaCl2, 1.2 MgCl2, 2 KH2PO4, 225 NaHCO3; pH 7.35). The buffers were maintained at 37°C by a heated water jacket and circulated by CO2. Fructose (10 mmol/l) was added to both sides. For measurement of basal mannitol fluxes, 1 mM of mannitol with 10
Ci [H3-mannitol] was added to the mucosal side. The spontaneous transepithelial potential difference (PD) was determined, and the tissue was clamped at zero voltage by continuously introducing an appropriate short-circuit current (Isc) with an automatic voltage clamp (DVC 1000 World Precision Instruments, New Haven, CT), except for 5–10 s every 10 min when PD was measured by removing the voltage clamp. Tissue ion conductance (G) was calculated from PD andIscaccording to Ohm’s Law (7).Iscis expressed asA/cm2, and G as mS/cm2.
Measurements in colonic epithelial cell lines.HT-29 and T84 cells were purchased from the American Type Culture Collection (Manas-sas, VA) and grown in RPMI 1640 media and DMEM/F12 media, respectively, with 5% and 10% heat-inactivated FBS. All experiments
Fig. 1. Colonic permeability and estrogen receptor-(ER) expression levels in IL-10⫺/⫺mice.A: mannitol fluxes were measured in Ussing chambers and
were significantly increased in IL-10⫺/⫺mice compared with wild-type
con-trols.B: levels of mRNA for ERare expressed as fold change compared with wild-type small intestine. Decreased ER expression was observed in the colon of IL-10⫺/⫺ mice compared with wild-type littermates. Values are
means ⫾ SE for 6 – 8 mice in each group. *P ⬍ 0.01 compared with age-matched wild-type males. **P⬍0.01 compared with age-matched wild-type females.
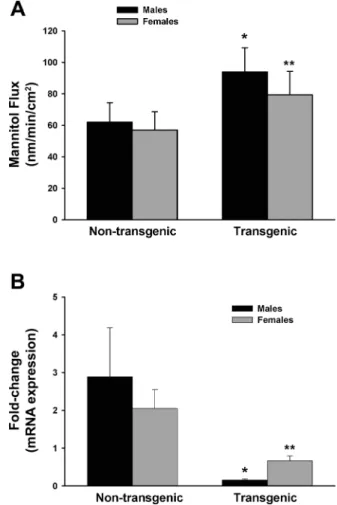
Fig. 2. Colonic permeability and ER expression levels in HLA-B27 rats.
A: mannitol fluxes were measured in Ussing chambers and were significantly increased in transgenic HLA-B27 rats compared with nontransgenic litter-mates.B: levels of mRNA for ERare expressed as fold change compared with nontransgenic small intestine. Decreased ERexpression was observed in the colon of HLA-B27 rats compared with nontransgenic littermates. Values are means⫾ SE for 6 – 8 mice in each group. *P ⬍0.05 compared with age-matched nontransgenic males. **P⬍0.05 compared with age-matched nontransgenic females.
by 10.220.33.6 on April 13, 2017
http://ajpgi.physiology.org/
were carried out in optiMEM (Invitrogen, Burlington, ON, Canada) without FBS supplementation to remove endogenous estrogen. For measurement of ER␣and ERmRNA levels, confluent monolayers were trypsinized and collected in Trizol. RNA isolation and RT-PCR were performed as described above. For measurements of epithelial barrier function, impedance and resistance were measured using the electric cell substrate impedance sensing (ECIS) 1600R (Applied Biophysics, Troy, NY). ECIS plates with 10 electrodes (8W10E) were incubated with 400l RPMI medium per well for at least 3 h at 37°C, after which the electrodes were stabilized following the manufactur-ers=instructions. T84 cells were grown until confluence. Cells were then trypsinized and resuspended with 1⫻106cells in 400l media seeded per well. After cells were confluent, monolayers were treated with either 17-estradiol (1–1,000 nM), diarylpropionitrile (DPN; 1–1,000 nM) or the estrogen receptor blocker ICI 182,780 (Sigma, Oakville, ON, Canada). Changes in resistance were measured for up to 15 h. The acquired data were analyzed for changes in resistance using the ECIS software.
Assessment of human biopsies. Biopsies were obtained from healthy individuals (n⫽13) undergoing screening colonoscopy and from patients with documented Crohn’s disease (n⫽18) or ulcerative colitis (n ⫽ 15) either currently in remission or suffering from a relapse in their disease. All protocols and procedures were approved by the University of Alberta Health Research Ethics board (no. 7113). All subjects provided written consent for tissue analysis. In all cases, biopsies were taken from macroscopically noninvolved tissue in the transverse colon, snap-frozen in liquid nitrogen, and stored at⫺80°C until further processing. RNA isolation and RT-PCR were performed for measurement of ER, TNF-␣, and IFN␥mRNA levels. Relative expression data for each cytokine are shown as normalized to the average ⌬CT of the control group of patients for the respective cytokine (31). The control group was normalized against itself but does not appear as “1” due to patient variability (24).
Statistics.Data are presented as means⫾SE. Data were tested for normality of distribution, and analyses were performed using the statistical software SigmaStat (Jandel, San Rafael, CA). Comparisons between multiple groups were carried out using one-way ANOVA with a post hoc test of significance between individual groups. Differences were considered significant whenPwas⬍0.05.
RESULTS
Intestinal ER mRNA levels and permeability in IL-10⫺/⫺
mice. We demonstrated IL-10⫺/⫺ mice have increased gut
permeability compared with wild-type mice both prior to and following the onset of colitis (25). In this study, we confirmed the increased colonic permeability in IL-10⫺/⫺ mice prior to the onset of colitis. As seen in Fig. 1A, both male and female IL-10⫺/⫺ mice had increased colonic permeability compared with wild-type mice as shown by increased mannitol flux in Ussing chambers. There was no histological inflammation seen in the colon of these mice (data not shown). We then measured the mRNA levels of ER to determine whether alterations in permeability were associated with ERexpression. As seen in Fig. 1B, reduced ER mRNA was seen in the colon of
IL-10⫺/⫺mice compared with wild-type littermates. Wild-type females showed a higher level of ER expression than wild-type males; however, no such differences were seen in IL-10⫺/⫺mice. ERwas not detectable in the small intestine in
either wild-type or IL-10⫺/⫺mice (data not shown).
Intestinal ERmRNA levels and permeability in HLA-B27 rats.To determine whether the reduction in ERwas specific to the IL-10⫺/⫺ mouse, a second model of chronic intestinal
inflammation was examined. HLA-B27 TG mice spontane-ously develop colitis beginning at 8 –12 wk of age (14). Similar to what was seen in the IL-10⫺/⫺mouse, ERexpression was significantly reduced in the colon at 6 wk of age, again in the Table 1. Colonic electrical measurements in 6-wk-old HLA-B27 transgenic (TG) and non-TG (NTG) rats
Isc,uA/cm2 Conductance, mS/cm2
Male Female Male Female
NTG,n⫽5 TG,n⫽5 NTG,n⫽6 TG,n⫽5 NTG,n⫽5 TG,n⫽6 NTG,n⫽6 TG,n⫽6
8.0⫾2.6 13.4⫾2.9 8.5⫾1.0 10.5⫾2.0 28.4⫾4.6 20.7⫾2.6 27.1⫾4.7 20.8⫾5.3 Values are means⫾SE.Isc, short-circuit current.
Fig. 3. Stimulation of ERresults in increased epithelial monolayer resistance as measured using electric cell-substrate impedance sensing (ECIS).A: treat-ment of T84 cells with 17-estradiol increased resistance over time in a dose-dependent fashion. ICI 182780 blocked the effects of 17-estradiol (100 nM). B: treatment of T84 cells with diarylpropionitrile (DPN) increased resistance over time in a dose-dependent fashion. Values are shown as % increases in monolayer resistance relative to the beginning resistance measure-ment in each well. Experimeasure-ments were repeated 3 times with duplicate mea-surements. *P⬍0.05 compared with control.
by 10.220.33.6 on April 13, 2017
http://ajpgi.physiology.org/
absence of any histological inflammation, compared with NTG littermates (Fig. 2B). There was no difference in ER expres-sion between males and females. Levels of ERin the small intestine were barely detectable (data not shown). To deter-mine whether HLA-B27 TG rats also demonstrated alterations in intestinal permeability, colonic tissue was studied in Ussing chambers. At 6 wk of age, mannitol flux was significantly increased in the colon of both male and female HLA-B27 TG rats compared with NTG littermates (Fig. 2A). There were no significant differences inIscor conductance (Table 1).
Stimulation of ER increases epithelial resistance.To fur-ther examine the relationship between ER signaling and epithelial permeability, we utilized two human colonic cell lines that have been shown to express ERbut not ER␣(2, 6). In agreement with previously reported findings (2), we found that neither cell line had any detectable expression of ER␣ (data not shown). The levels of ERmRNA were significantly higher in T84 (5.4 ⫾ 0.9-fold change gene expression) com-pared with HT-29 (1.1⫾ 0.3-fold change) cells, and this was associated with increased epithelial resistance in T84 (2,491⫾ 56 ohms/cm2) compared with HT-29 (824 ⫾ 13 ohms/cm2). Treatment of T84 cells with the ER agonists, 17-estradiol (1–1,000 nM) (Fig. 3A) or DPN (1–1,000 nM) (Fig. 3B) resulted in enhanced monolayer resistance in a dose-dependent fashion. Furthermore, treatment of T84 epithelial cells with the estrogen receptor blocker ICI 182,780 blocked the effects of 17-estradiol (Fig. 3A). These data support a role for ER signaling in modulating epithelial resistance.
ERmRNA levels in human biopsies.Several studies have demonstrated IBD patients to have increased intestinal perme-ability (28, 33). To determine whether ER expression was altered in IBD patients, colonic biopsies were obtained from healthy individuals undergoing screening colonoscopy (n ⫽
13) and ERmRNA levels were measured and compared with levels in Crohn’s disease patients in remission (n ⫽ 13) or relapse (n⫽5) and ulcerative colitis patients in remission (n⫽
9) or relapse (n⫽6). Characteristics of each group are listed in Table 2. All biopsies were taken from grossly noninflamed tissue of the transverse colon. Similar to our observations in the animal models of colitis, a reduction in ERmRNA levels was observed in colonic biopsies from the Crohn’s disease patients in relapse, but interestingly, not the patients in remission. In contrast, in the ulcerative colitis patients, both the patients in remission and the patients suffering from relapse demonstrated reduced levels of ERmRNA (Fig. 4). There were no apparent differences in ERmRNA levels between males and females in this small sample size. To determine whether the reduction in ERlevels was related to enhanced levels of proinflamma-tory cytokines, TNF-␣ and IFN␥ expression were measured. As seen in Table 3, there was no apparent relationship between levels of these cytokines and expression of ER. In fact,
patients with Crohn’s disease actually had reduced levels of IFN␥ compared with controls.
DISCUSSION
In this study, increased colonic permeability and decreased levels of ERmRNA were found in colonic tissue prior to the onset of inflammation in two rodent models of chronic intes-tinal inflammation. This reduction in ER mRNA expression was also seen in grossly noninflamed colonic biopsies from ulcerative colitis patients and Crohn’s disease patients who were suffering from a relapse. Furthermore, this study showed that treatment of T84 cells with either 17-estradiol or the ER agonist, DPN, resulted in a dose-dependent increase in mono-layer resistance.
A role for ERin maintenance of colonic homeostasis and barrier function has been suggested by the findings of Wada-Hiraike et al. (36) in their study utilizing ER⫺/⫺ mice. Our findings support a possible role for ERsignaling in modulat-ing colonic barrier function, in that reduced levels of ERwere found in conjunction with increased colonic permeability in both the IL-10⫺/⫺mouse and the HLA-B27 rat. Unfortunately, protein expression of ERwas not able to be measured in these studies due to problems with specificity with existing antibod-ies (32), which is a limitation of these studantibod-ies. Proinflammatory cytokines have been shown to downregulate estrogen receptors in epithelial cells (23, 34); thus the reduction in ERmRNA levels prior to the onset of colitis in these rodent models could have been related to increased levels of proinflammatory cy-tokines that are present prior to the onset of inflammation in these models (30). However, decreased levels of ER expres-Table 2. Patients demographic profile
Group
Gender
Age (mean and range) Disease Status Biopsy Site
Female Male
Control,n⫽13 7 6 53 (18–88) N/A Transverse colon
Crohn’s disease,n⫽18 8 10 42 (24–62) Relapse,n⫽5; remission,n⫽13 Transverse colon Ulcerative colitis,n⫽15 10 5 39 (18–64) Remission,n⫽9; relapse,n⫽6 Transverse colon
NA, not applicable.
Fig. 4. ERmRNA levels in colonic biopsies from healthy individuals and patients with Crohn’s disease (CD) or ulcerative colitis (UC). CD patients in relapse and UC patients had significantly reduced levels of ERexpression compared with healthy controls. All biopsies were taken from noninflamed regions of the transverse colon. Rem, remission. Values are means⫾SD. *P⬍0.05 compared with controls.
by 10.220.33.6 on April 13, 2017
http://ajpgi.physiology.org/
sion were also observed in colonic biopsies from IBD patients in grossly uninflamed regions of the intestine, and this decrease did not correlate with increased levels of either TNF-␣or IFN␥ in individual patients. Thus it is unlikely that the reduction in ER mRNA expression in IBD patients was due to increased TNF-␣or IFN␥ levels.
The finding that patients with Crohn’s disease in remission had normal levels of ERmRNA expression, while patients in relapse had reduced levels is particularly interesting in view of the findings that increased intestinal permeability is predictive of subsequent relapse (3). However, as with the animal studies, protein levels of ER could not be determined; thus future studies are required to determine the exact role of ER signal-ing in patients with IBD. The underlysignal-ing cause for this de-crease in ERexpression in IBD patients cannot be determined from this study, but may be either genetically determined, or alternatively, related to the nutritional status of these patients. IBD patients commonly exhibit reduced plasma zinc levels, which can directly influence expression and sensitivity of estrogen receptors (10). Zinc deficiency also has numerous effects on the immune system that ultimately result in in-creased levels of oxidative stress and proinflammatory cyto-kine release, both of which can have a role in the breakdown of gut barrier function (29). Future studies are necessary to determine the underlying mechanism responsible for these observed decreased levels of ER, and also whether alterations in ER expression are linked with changes in intestinal per-meability in IBD patients.
Beneficial effects of estrogen and ERagonists in different animal models of chronic colitis have been reported (13–15, 17). Harnish et al. (14) and Harris (15) reported improved histological scores, decreased inflammatory cell infiltration, and improvement of chronic diarrhea by estradiol treatment or administration of ER agonist ERB-041. Although intestinal permeability was not measured in these studies, we previously showed that treatment aimed at correcting the barrier defect in the IL-10⫺/⫺mouse is beneficial in both preventing the onset or and attenuating existing colitis, (4, 11). These findings support a role for gut permeability in modulating colonic inflammation in this animal model.
Previous studies demonstrated variable levels of ER ex-pression and no ER␣ expression in the T84 and HT-29 cell lines (2, 6). We confirmed that the T84 cell line used in these studies expressed significantly higher levels of ER mRNA compared with HT-29 cells, and that neither cell line had ER␣ expression. This increased level of ERmRNA was associated with a significantly higher resistance in T84 cells compared with HT-29 cells. Exposure of cell monolayers to either 17 -estradiol or the ER agonist DPN increased resistance, strongly supporting a role for ER signaling in modulating epithelial permeability. Supporting this hypothesis are the data
showing that blocking of estrogen receptors with ICI 182,780 prevented the increase in the presence of 17-estradiol. Inter-estingly, this increased resistance in response to 17-estradiol is in contrast to effects in endothelial cells or vaginal-cervical epithelial cells, which both exhibit a decreased resistance when exposed to estrogen (1, 12). However, these cells primarily express ER␣, suggesting that ER␣ and ER signaling may have opposing effects on epithelial permeability.
To the best of our knowledge this is the first study to recognize an altered expression of ERmRNA in the colon of experimental models of colitis and samples from patients with inflammatory bowel disease. Interestingly, this decrease in ER expression is present in grossly noninflamed colonic tissue in IBD patients. In conclusion, a better understanding of the role of ERand its modulating effect on intestinal perme-ability might help in the understanding of the complexity of inflammatory bowel diseases, and lead to novel therapeutic options.
ACKNOWLEDGMENTS
The authors thank Matt Emberg for technical assistance. GRANTS
These studies were supported by the Crohn’s and Colitis Foundation of Canada, Canadian Institutes for Health Research, and the Alberta Heritage Foundation for Medical Research (now Alberta Innovates–Health Solutions). DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s). REFERENCES
1. Aberdeen GW, Wiegand SJ, Bonagura TW II, Pepe GJ, Albrecht ED. Vascular endothelial growth factor mediates the estrogen-induced break-down of tight junctions between and increase in proliferation of microves-sel endothelial cells in the baboon endometrium. Endocrinology 149: 6076 –6083, 2008.
2. Arai N, Strom A, Rafter JJ, Gustafsson JA.Estrogen receptormRNA in colon cancer cells: growth effects of estrogen and genistein.Biochem Biophys Res Commun270: 425–431, 2000.
3. Arnott ID, Kingstone K, Ghosh S. Abnormal intestinal permeability predicts relapse in inactive Crohn’s disease.Scand J Gastroenterol35: 1163–1169, 2000.
4. Arrieta M, Doyle J, Madsen K, Meddings J.Reducing small intestinal permeability attenuates colitis in the IL-10 gene deficient mouse.Gut58: 41–48, 2009.
5. Boyko EJ, Theis MK, Vaughan TL, Nicol-Blades B.Increased risk of inflammatory bowel disease associated with oral contraceptive use.Am J Epidemiol140: 268 –278, 1994.
6. Campbell-Thompson M, Lynch IJ, Bhardwaj B.Expression of estrogen receptor (ER) subtypes and ERisoforms in colon cancer.Cancer Res61: 632–640, 2001.
7. Clarkson T.Measurement of short-circuit current and ion transport across the ileum.Am J Physiol206: 658 –668, 1964.
8. Cornish JA, Tan E, Simillis C, Clark SK, Teare J, Tekkis PP.The risk of oral contraceptives in the etiology of inflammatory bowel disease: a meta-analysis.Am J Gastroenterol103: 2394 –2400, 2008.
Table 3. Relative quantitation of TNF-␣and IFN␥in mucosal biopsies
Group Disease Status TNF-␣ IFN␥
Control,n⫽13 N/A 1.44⫾1.08 (0.3–3.4) 1.83⫾2.15 (0.4–7.3)
Crohn’s disease,n⫽18 Remission,n⫽13; 1.22⫾0.87 (0.3–-2.5) 0.61⫾0.41* (0.3–2.4) relapse,n⫽5 1.34⫾0.92 (0.2–2.4) 0.74⫾0.76 (0.2–1.8) Ulcerative colitis,n⫽15 Remission,n⫽9; 1.80⫾2.2 (0.1–6.9) 2.49⫾2.84 (0.1–7.6) relapse,n⫽6 1.39⫾1.75 (0.3–3.4) 4.20⫾5.45 (0.9–10.5) Values are means⫾SE. Numbers in parentheses are range. *P⬍0.05 compared with control.
by 10.220.33.6 on April 13, 2017
http://ajpgi.physiology.org/
9. Dahlman-Wright K, Cavailles V, Fuqua SA, Jordan VC, Katzenellen-bogen JA, Korach KS, Maggi A, Muramatsu M, Parker MG, Gustafs-son JA.International Union of Pharmacology. LXIV. Estrogen receptors.
Pharmacol Rev58: 773–781, 2006.
10. El-Tawil AM.Oestrogens and Crohn’s disease: the missed link. Andro-logia40: 141–145, 2008.
11. Ewaschuk J, Diaz H, Meddings L, Diederichs B, Dmytrash A, Backer J, Looijer-van Langen M, Madsen KL.Secreted bioactive factors from
Bifidobacterium infantis enhance epithelial cell barrier function. Am J Physiol Gastrointest Liver Physiol295: G1025–G1034, 2008.
12. Gorodeski GI.Estrogen decrease in tight junctional resistance involves matrix-metalloproteinase-7-mediated remodeling of occludin. Endocrinol-ogy148: 218 –231, 2007.
13. Gunal O, Oktar BK, Ozcinar E, Sungur M, Arbak S, Yegen B. Estradiol treatment ameliorates acetic acid-induced gastric and colonic injuries in rats.Inflammation27: 351–359, 2003.
14. Harnish DC, Albert LM, Leathurby Y, Eckert AM, Ciarletta A, Kasaian M, Keith JC Jr.Beneficial effects of estrogen treatment in the HLA-B27 transgenic rat model of inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol286: G118 –G125, 2004.
15. Harris HA.Estrogen receptor-: recent lessons from in vivo studies.Mol Endocrinol21: 1–13, 2007.
16. Hollander D, Vadheim CM, Brettholz E, Petersen GM, Delahunty T, Rotter JI. Increased intestinal permeability in patients with Crohn’s disease and their relatives. A possible etiologic factor.Ann Intern Med
105: 883–885, 1986.
17. Houdeau E, Moriez R, Leveque M, Salvador-Cartier C, Waget A, Leng L, Bueno L, Bucala R, Fioramonti J.Sex steroid regulation of macrophage migration inhibitory factor in normal and inflamed colon in the female rat.Gastroenterology132: 982–993, 2007.
18. Kane SV, Reddy D.Hormonal replacement therapy after menopause is protective of disease activity in women with inflammatory bowel disease.
Am J Gastroenterol103: 1193–1196, 2008.
19. Kaser A, Zeissig S, Blumberg RS.Inflammatory bowel disease.Annu Rev Immunol28: 573–621, 2010.
20. Katz KD, Hollander D, Vadheim CM, McElree C, Delahunty T, Dadufalza VD, Krugliak P, Rotter JI.Intestinal permeability in patients with Crohn’s disease and their healthy relatives. Gastroenterology97: 927–931, 1989.
21. Konstantinopoulos PA, Kominea A, Vandoros G, Sykiotis GP, Andri-copoulos P, Varakis I, Sotiropoulou-Bonikou G, Papavassiliou AG. Oestrogen receptor-(ER) is abundantly expressed in normal colonic mucosa, but declines in colon adenocarcinoma paralleling the tumour’s dedifferentiation.Eur J Cancer39: 1251–1258, 2003.
22. Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novel receptor expressed in rat prostate and ovary.Proc Natl Acad Sci USA93: 5925–5930, 1996.
23. Lee SH, Nam HS.TNF-␣-induced down-regulation of estrogen recep-tor-␣in MCF-7 breast cancer cells.Mol Cells26: 285–290, 2008. 24. Livak KJ, Schmittgen TD. Analysis of relative gene expression data
using real-time quantitative PCR and the 2⫺⌬⌬CTmethod.Methods25:
402–408, 2001.
25. Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, Yachimec C, Doyle J, Jewell L, De Simone C.Probiotic bacteria enhance murine and human intestinal epithelial barrier function.Gastroenterology121: 580 – 591, 2001.
26. Madsen KL, Doyle JS, Jewell LD, Tavernini MM, Fedorak RN. Lactobacillus species prevents colitis in interleukin 10 gene-deficient mice.Gastroenterology116: 1107–1114, 1999.
27. Meddings JB, Sutherland LR, May GR. Intestinal permeability in patients with Crohn’s disease.Gut35: 1675–1676, 1994.
28. Munkholm P, Langholz E, Hollander D, Thornberg K, Orholm M, Katz KD, Binder V. Intestinal permeability in patients with Crohn’s disease and ulcerative colitis and their first degree relatives. Gut 35: 68 –72, 1994.
29. Prasad AS.Zinc in human health: effect of zinc on immune cells.Mol Med14: 353–357, 2008.
30. Rennick DM, Fort MM. Lessons from genetically engineered animal models. XII. IL-10-deficient IL-10(⫺/⫺) mice and intestinal inflamma-tion.Am J Physiol Gastrointest Liver Physiol278: G829 –G833, 2000. 31. Schmittgen TD, Livak KJ.Analyzing real-time PCR data by the
com-parative CTmethod.Nat Protoc3: 1101–1108, 2008.
32. Snyder MA, Smejkalova T, Forlano PM, Woolley CS.Multiple ER antisera label in ER knockout and null mouse tissues. J Neurosci Methods188: 226 –234, 2010.
33. Soderholm JD, Olaison G, Peterson KH, Franzen LE, Lindmark T, Wiren M, Tagesson C, Sjodahl R.Augmented increase in tight junction permeability by luminal stimuli in the non-inflamed ileum of Crohn’s disease.Gut50: 307–313, 2002.
34. Stallmach A, Giese T, Schmidt C, Ludwig B, Mueller-Molaian I, Meuer SC. Cytokine/chemokine transcript profiles reflect mucosal in-flammation in Crohn’s disease.Int J Colorectal Dis19: 308 –315, 2004. 35. Wada-Hiraike O, Imamov O, Hiraike H, Hultenby K, Schwend T, Omoto Y, Warner M, Gustafsson JA.Role of estrogen receptor beta in colonic epithelium.Proc Natl Acad Sci USA103: 2959 –2964, 2006. 36. Wada-Hiraike O, Warner M, Gustafsson JA. New developments in
oestrogen signalling in colonic epithelium.Biochem Soc Trans34: 1114 – 1116, 2006.
by 10.220.33.6 on April 13, 2017
http://ajpgi.physiology.org/