HELGOI_~NDER MEERESUNTERSUCHUNGEN Helgol~inder M e e r e s u n t e r s . 37, 493-504 (1984)
Alterations in patterns of e x c r e t i o n and other m e t a b o l i c
functions in d e v e l o p i n g fish e m b r y o s e x p o s e d to
b e n z o ( a ) p y r e n e
R. M. K o c a n & M. L.
L a n d o l t
University of Washington, School of Fisheries, W H - I O; Seattle, Washington 98195, USA
ABSTRACT: S t e e l h e a d trout
(Salmo gairdneri)
e m b r y o s w e r e u s e d as a m o d e l to study some of the physiological and b i o c h e m i c a l c h a n g e s w h i c h occur following exposure to s u b l e t h a l levels of benzo(a)pyrene B(a)P, a k n o w n e n v i r o n m e n t a l contaminant. Embryos w e r e e x p o s e d to B(a)P on day 1, 15, or 25 post fertilization, c o r r e s p o n d i n g to the pre-blastula, e y e d stage a n d late o r g a n o g e n e s i s stage of d e v e l o p m e n t . T h e s e times w e r e c h o s e n to d e t e r m i n e w h e t h e r t h e a g e of the embryo at the time of exposure i n f l u e n c e d their r e s p o n s e to this m o d e l compound. The c h a n g e s w h i c h w e r e o b s e r v e d w e r e related to the a g e of the e m b r y o at the time of exposure but not to the concentration of the B(a)P to w h i c h they w e r e exposed. It was found that the l o n g e r the e m b r y o d e v e l o p e d , the more p e r m e a b l e the e g g s b e c a m e , taking up twice to 10 times as m u c h B(a)P on day 25 as t h e y did on day 1. Embryos e x p o s e d later in d e v e l o p m e n t w e r e also able to excrete the metabolites of B(a)P through the e g g m e m b r a n e 10 times more rapidly t h a n their counterparts w h i c h w e r e e x p o s e d early during d e v e l o p m e n t . Following hatching, the larvae from those groups e x p o s e d later in d e v e l o p - m e n t c o n t a i n e d 2 to 5 times as m u c h B(a)P b o u n d to or dissolved in the tissues t h a n did those e x p o s e d on day 1. H a t c h i n g time was also m o d i f i e d by the times of exposure, with those e x p o s e d on days 15 and 25 h a t c h i n g later a n d over a l o n g e r p e r i o d of time t h a n e i t h e r the u n t r e a t e d or DMSO controls. Although physical abnormalities w e r e rare, there a p p e a r e d to b e a consistent increase in the level of t e r a t o g e n e s i s in those groups w h i c h w e r e e x p o s e d early d u r i n g d e v e l o p m e n t .I N T R O D U C T I O N
O u r i n a b i l i t y to d o c u m e n t a c c u r a t e l y t h e c a u s e of p o p u l a t i o n d e c l i n e s i n a q u a t i c s p e c i e s is l a r g e l y t h e r e s u l t of t h e i n a c c e s s a b i l i t y of m o r i b u n d i n d i v i d u a l s . M a s s i v e , s u d d e n m o r t a l i t y is a n e x c e p t i o n i n w h i c h w e c a n f r e q u e n t l y d e t e r m i n e b o t h t h e c a u s e of t h e d i e - o f f a s w e l l a s t h e s o u r c e of t h e c a u s a t i v e a g e n t ( s ) . M o r e f r e q u e n t l y , h o w e v e r , a s p e c i e s g r a d u a l l y d e c l i n e s i n a b u n d a n c e a n d f i n a l l y d i s a p p e a r s e n t i r e l y f r o m a n a r e a i n w h i c h it o n c e f l o u r i s h e d . T h e r e is u s u a l l y n o s h o r t a g e of s p e c u l a t i o n t o e x p l a i n t h e s e d e c l i n e s , s u c h a s o v e r e x p l o i t a t i o n , h a b i t a t d e g r a d a t i o n , c o m p e t i t i o n w i t h i n t r o d u c e d s p e c i e s a n d r e c e n t l y e v e n t h e E1 Nifio. H a r d s c i e n t i f i c e v i d e n c e h o w e v e r , is u s u a l l y l a c k i n g . T h e r e a s o n f o r t h i s is t h a t t h e c a u s e of s u c h i n s i d i o u s m o r t a l i t y is g r e a t l y s e p a r a t e d i n t i m e f r o m t h e f i n a l r e c o g n i z a b l e e f f e c t - t h e l o s s of a s p e c i e s f r o m its h a b i t a t .
494 R . M . K o c a n & M. L. L a n d o l t
e x h i b i t c h r o m o s o m a l d a m a g e (Kocan et al., 1982), as w e l l as d e v e l o p i n g m u t a t i o n s (Kocan et al., 1981) m u c h as m a m m a l i a n cells do. T h e c h r o m o s o m e d a m a g e s e e n i n vitro has also b e e n v e r i f i e d i n vivo i n o u r l a b o r a t o r y b y Liguori (Ph.d. Thesis, 1984) as well as b e i n g d e s c r i b e d u n d e r n a t u r a l c o n d i t i o n s (Longwell & H u g h e s , 1980). K l i g e r m a n & Bloom (1976) a n d K l i g e r m a n (1979) d e m o n s t r a t e d c h r o m o s o m a l d a m a g e of a different t y p e (SCE) a n d i n a d i f f e r e n t species, i n d i c a t i n g t h a t such s u b l e t h a l D N A d a m a g e is not
u n c o m m o n .
In addition to physical defects observed in D N A , w e have also seen changes in the capacity of trout embryos and fry to excrete xenobiotics, such as polycyclic aromatic
hydrocarbons, following embryonic exposure. Biochemical alterations in e n z y m e func- tion have also been reported by several investigators working with embryonic fish of
different species (Binder & Stegeman, 1980, 1983).
All of these observations suggest that sublethal exposure of fish embryos to toxic
substances m a y result in their inability to successfully compete with normal individuals
at s o m e later period in life due to persistent sublethal d a m a g e which occurred earlier. In order to clarify s o m e of the causes and relationships of these early biochemical altera- tions, w e e x a m i n e d the dynamics of e m b r y o development and s o m e of the critical
processes associated with early life, after the embryos were exposed to a k n o w n toxic substance. T h e concentrations of toxin and methods of exposure were chosen to insure
experimental uniformity and consistent uptake by the eggs, and in no w a y were
intended to duplicate natural exposures.
MATERIALS AND M E T H O D S
E x p e r i m e n t a l e m b r y o s
S t e e l h e a d trout
(Salmo gairdneri)
e m b r y o s w e r e o b t a i n e d from a s i n g l e f e m a l e w h i c h h a d r e t u r n e d to the U n i v e r s i t y of W a s h i n g t o n e x p e r i m e n t a l h a t c h e r y to spawn. F o l l o w i n g f e r t i l i z a t i o n b y a s i n g l e m a l e s t e e l h e a d , the e g g s w e r e m a i n t a i n e d i n lots of 100 i n n e t e g g c h a m b e r s s u s p e n d e d i n a f l o w - t h r o u g h s y s t e m of d e c h l o r i n a t e d city w a t e r at 10 °C. T e m p e r a t u r e , w a t e r flow a n d c h l o r i n e w e r e m o n i t o r e d c o n t i n u o u s l y .T e s t c o m p o u n d
14C-benzo(a)pyrene (14C-B(a)P) (Sp. Act. 58.5 m C i / m M o l - A m e r s h a m ) was d i l u t e d w i t h u n l a b e l l e d B(a)P to m a k e a stock s o l u t i o n of 10btg/~tl i n s p e c t r o p h o t o m e t r i c grade d i m e t h y l s u l p h o x i d e (DMSO). T h i s s o l u t i o n w a s u s e d to m a k e f i n a l e x p o s u r e c o n c e n t r a - tions of 2~tg a n d 20btg/ml of w a t e r (0.5% DMSO). T h e f i n a l activity of 14C-B(a)P was 0.23 t~Ci/tlg.
D e v e l o p i n g fish e m b r y o s e x p o s e d to b e n z o ( a ) p y r e n e 495
300. W h e n c o m p a r i s o n s b e t w e e n e g g s or l a r v a e c o n t a i n i n g d i f f e r e n t a m o u n t s of p i g m e n t w e r e b e i n g m a d e , q u e n c h i n g for each system w a s t a k e n into account, thus e l i m i n a t i n g a n y p o s s i b i l i t y of error w h e n c o m p a r i n g different types of tissue.
E x p o s u r e s
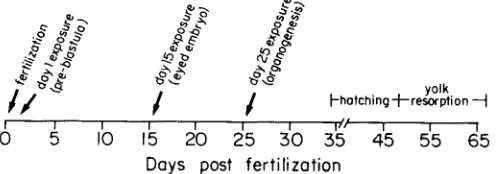
T h r e e g r o u p s of 400 eggs each w e r e e x p o s e d to u n l a b e l l e d B(a)P (Sigma, G o l d Lable) o n d a y 1, d a y 15, a n d d a y 25 post fertilization. Controls c o n s i s t e d of 600 u n t r e a t e d a n d 200 DI~SO t r e a t e d eggs. T h e t h r e e e x p o s u r e p e r i o d s (early, m i d a n d late d e v e l o p - ment) c o r r e s p o n d to p r e - b l a s t u l a (day 1), e y e d e m b r y o (day 15), a n d late o r g a n o g e n e s i s (day 25), a n d serve to d e m o n s t r a t e w h e t h e r the s t a g e of d e v e l o p m e n t of the e m b r y o i n f l u e n c e s the o u t c o m e of e x p o s u r e to B(a)P (Fig. 1). A p a r a l l e l set of 50 e g g s w a s exposed to 14C-B(a)P i n order to d e t e r m i n e the rate of u p t a k e of t h e test c o m p o u n d at each exposure period.
1
1
~-hotching Jf- resor Pti on --i yolki// i ~ i
55
45
55
65
Days post fertilization
Fig. 1. Trout embryo development and benzo(a)pyrene exposure schedule. Following fertilization, eggs were allowed to water harden for 24 h, after which they were exposed to B(a)P in water (0.5 % DMSO) for 24 h. On days 15 and 25 post fertilization, two additional groups of eggs were similarly exposed. Eggs were sampled following each exposure period and again just prior to hatching. After hatching, sac fry (larva and yolk) were also collected for analysis and for collection of excreted
water soluble metabolites of B(a)P
H a t c h i n g
T h e effect of B(a)P o n h a t c h i n g w a s d e t e r m i n e d b y r e c o r d i n g t h e n u m b e r of e g g s w h i c h h a t c h e d each d a y u n t i l all e g g s w e r e h a t c h e d . Each of the t r e a t m e n t g r o u p s a n d controls c o n s i s t e d of four r e p l i c a t e s of 100 e g g s each. T h e four t r e a t m e n t g r o u p s were: (1) u n t r e a t e d controls, (2) DMSO t r e a t e d controls, (3) e g g s e x p o s e d o n e d a y post fertilization (pre-blastula), (4) e g g s e x p o s e d 15 d a y s post f e r t i l i z a t i o n (eyed stage), a n d (5) e g g s e x p o s e d 25 days post fertilization (late o r g a n o g e n e s i s ) .
M e t a b o l i s m / e x c r e t i o n
496 R . M . K o c a n & M. L. L a n d o l t
at h a t c h i n g ( p e r i v i t e l l i n e fluid) a n d the a m o u n t p r e s e n t i n the larva a n d yolk after h a t c h i n g . By d i s s e c t i n g t h e l a r v a free from t h e y o l k w e c o u l d also d e t e r m i n e the a m o u n t p r e s e n t i n the y o l k a n d l a r v a s e p a r a t e l y .
A s e c o n d m e t h o d of d e t e r m i n i n g t h e a m o u n t of l a b e l l e d test c o m p o u n d excreted b y the d e v e l o p i n g e g g s w a s to p l a c e i n d i v i d u a l e g g s i n s m a l l vials a n d cover t h e m with a s m a l l a m o u n t of sterile water. By r e m o v i n g t h e e n t i r e v o l u m e of w a t e r t h r e e t i m e s p e r w e e k a n d r e p l a c i n g it w i t h fresh sterile water, we c o u l d d e t e r m i n e the a m o u n t of l a b e l l e d c o m p o u n d lost into the w a t e r e v e r y t h r e e days. T h e a c c u m u l a t e d counts t h r o u g h o u t i n c u b a t i o n w o u l d t h e n r e p r e s e n t t h e total a m o u n t of l a b e l l e d B(a)P lost from the t i m e of e x p o s u r e u n t i l h a t c h i n g . In this w a y it w a s also p o s s i b l e to m e a s u r e the a m o u n t of r a d i o a c t i v i t y r e l e a s e d w i t h t h e p e r i v i t e l l i n e fluids at t h e t i m e of h a t c h i n g .
This s a m e t e c h n i q u e of i n c u b a t i n g i n a closed s y s t e m w a s also u s e d to collect the e x c r e t e d m e t a b o l i t e s of sac fry d u r i n g t h e p e r i o d of yolk resorption. D u r i n g yolk r e s o r p t i o n fry w e r e k e p t i n a closed s y s t e m w h e r e the w a t e r w a s collected three to four t i m e s p e r w e e k a n d r e p l a c e d w i t h fresh sterile water. This w a t e r c o n t a i n e d all of the e x c r e t e d m a t e r i a l from t h e d e v e l o p i n g l a r v a e a n d w a s u s e d to d e t e r m i n e if a n y differ- e n c e s i n m e t a b o l i c c a p a b i l i t y h a d r e s u l t e d from e x p o s u r e of the eggs at different d e v e l o p m e n t a l periods.
D u r i n g the c o l l e c t i o n period, w a t e r from e a c h t r e a t m e n t g r o u p w a s p l a c e d into a d a r k bottle, g a s s e d w i t h n i t r o g e n a n d frozen at -- 20 °C u n t i l it w a s extracted. Extractions c o n s i s t e d of a s i n g l e h e x a n e e x t r a c t i o n (4 h e x a n e : 1 water) followed b y c e n t r i f u g a t i o n at 2000 r p m for 5 m i n to e n s u r e that the two l a y e r s w e r e fully separated. F o l l o w i n g the h e x a n e e x t r a c t i o n t h e w a t e r l a y e r w a s a g a i n e x t r a c t e d twice with e t h y l a c e t a t e (4:1). The h e x a n e e x t r a c t i o n w a s i n t e n d e d to r e m o v e a n y n o n p o l a r m a t e r i a l w h i c h m i g h t h a v e b e e n p r e s e n t a n d t h e e t h y l a c e t a t e to r e m o v e a n y s l i g h t l y p o l a r m e t a b o l i t e s (Phase I reactants).
A n a l i q u o t of e a c h w a t e r s a m p l e w a s also i n c u b a t e d w i t h [3-glucuronidase (1,000 u n i t s / m l ) a n d s u l p h a t a s e (45 u n i t s / m l ) for 60 m i n to d e c o n j u g a t e a n y s u l p h a t e or g l u c u r o n i c acid c o n j u g a t e s (Phase II reactants) w h i c h m i g h t b e present. In this w a y w e w e r e a b l e to d e t e r m i n e the r e l a t i v e a m o u n t of s o l u b l e u n c o n j u g a t e d a n d c o n j u g a t e d m e t a b o l i t e s e x c r e t e d b y the v a r i o u s t r e a t m e n t groups.
T e r a t o g e n e s i s
From t h e t i m e of h a t c h i n g u n t i l the y o l k w a s fully resorbed, the four g r o u p s of sac fry w e r e e x a m i n e d t h r e e t i m e s p e r w e e k u n t i l t h e y w e r e 60 days old. D u r i n g this period all a n i m a l s w h i c h a p p e a r e d to b e p h y s i c a l l y or b e h a v i o u r a l l y a b n o r m a l w e r e r e m o v e d a n d fixed i n 10 % f o r m a l i n for future classification.
RESULTS
U p t a k e of B ( a ) P
D e v e l o p i n g fish e m b r y o s e x p o s e d to b e n z o ( a ) p y r e n e
Table 1. Concentration of 14C-B(a)P in trout eggs following 24 h exposure
497
Exposure concen- N Exposure day (post-fertilization)
tration (~g/ml)
1 15 25
2 10 24 ± 2" 54 ± 10 59 ± 10
20 10 83 ± 11 729 ± 123 664 ± 87
* Values represent nanograrns (ng) of B(a)P for entire egg (i.e. yolk, embryo, chorion and perivitelline fluid ± S.D.)
i m m e d i a t e l y after t h e 24-h e x p o s u r e p e r i o d . T h e s e v a l u e s w e r e u s e d to c a l c u l a t e t h e p e r c e n t loss a n d r e t e n t i o n p r e s e n t e d l a t e r in this s e c t i o n u n d e r " E x c r e t i o n " . It is e v i d e n t that the n e w l y f e r t i l i z e d e g g is not as p e r m e a b l e to B(a)P in t h e form w e u s e d for exposure, as are t h e m o r e d e v e l o p e d e g g s .
T h e a c t u a l m e a s u r e d c o n c e n t r a t i o n s of B(a)P in t h e w a t e r at t h e start of e x p o s u r e w e r e 1.6-1.8 ~tg/ml for t h e 2 ~tg/ml e x p o s u r e water, a n d 16.1-19.8 ~ g / m l for t h e 20 ~tg/ml e x p o s u r e water. F o l l o w i n g t h e 24-h e x p o s u r e , t h e s e c o n c e n t r a t i o n s w e r e r e d u c e d to 1.4-1.6~tg/ml a n d 12.8-15.1~tg/ml: a 1 2 % a n d 2 0 % loss r e s p e c t i v e l y .
E x c r e t i o n
A distinct d i f f e r e n c e w a s o b s e r v e d in t h e r a t e of e x c r e t i o n of w a t e r s o l u b l e m e t a b o - lites of B(a)P from e m b r y o s e x p o s e d e a r l y vs t h o s e e x p o s e d l a t e in d e v e l o p m e n t (Fig. 2). T h o s e e x p o s e d p r i o r to b l a s t u l a f o r m a t i o n e x c r e t e o n l y 2 to 3 % of t h e total a m o u n t of B(a)P w h i c h w a s in t h e e g g f o l l o w i n g e x p o s u r e , w h i l e t h o s e e x p o s e d l a t e in i n c u b a t i o n lost as m u c h as 20 % of t h e total a m o u n t o r i g i n a l l y t a k e n u p d u r i n g e x p o s u r e . T h e g r o u p e x p o s e d 15 days f o l l o w i n g f e r t i l i z a t i o n (eyed stage), e x h i b i t e d an e x c r e t i o n rate m i d w a y b e t w e e n the e a r l y a n d l a t e e x p o s u r e groups. T h e loss of l a b e l l e d B(a)P w h i c h o c c u r r e d in the d a y 1 e x p o s u r e g r o u p took p l a c e d u r i n g t h e e n t i r e i n c u b a t i o n p e r i o d ( 3 0 + days), w h i l e t h o s e e x p o s e d l a t e in d e v e l o p m e n t h a d o n l y 10 to 15 d a y s to e x c r e t e t h e B(a)P p r i o r to h a t c h i n g . C o n s i d e r i n g this, t h e p e r d a y loss of B(a)P w a s e v e n m o r e s p e c t a c u l a r in t h e l a t e e x p o s u r e group. T h i s e x p e r i m e n t w a s r e p e a t e d w i t h t h r e e s e p a r a t e e g g lots f r o m t h r e e d i f f e r e n t f e m a l e s , a n d t h e results w e r e e s s e n t i a l l y i d e n t i c a l e a c h time. L i k e w i s e , the loss of s o l u b l e B(a)P from t h e e g g w a s t h e s a m e w h e t h e r m e a s u r e d d i r e c t l y by s a m p l i n g the i n c u b a t i o n w a t e r or by m e a s u r i n g t h e d i f f e r e n c e in r a d i o a c t i v i t y i n s i d e the e g g from t h e t i m e of e x p o s u r e u n t i l j u s t p r i o r to h a t c h i n g . In a s u b s e q u e n t e x p e r i m e n t a s i n g l e b a t c h of e g g s w a s split into t w o e q u a l lots w i t h o n l y o n e b e i n g f e r t i l i z e d . Both g r o u p s w e r e e x p o s e d to 14C-B(a)P a n d a l l o w e d to d e v e l o p for 20 days, d u r i n g w h i c h e g g s w e r e s a m p l e d t w i c e p e r w e e k . At t h e e n d of t h e 20 d a y s t h e r a d i a t i o n l e v e l s in t h e e g g s from b o t h g r o u p s w a s c o m p a r e d a n d f o u n d to b e i d e n t i c a l . T h i s i n d i c a t e d t h a t t h e s m a l l a m o u n t of c o m p o u n d lost from t h e e g g s e x p o s e d e a r l y in d e v e l o p m e n t w a s not t h e r e s u l t of e m b r y o m e t a b o l i s m a n d e x c r e t i o n , s i n c e e v e n e g g s w h i c h c o n t a i n e d no e m b r y o lost t h e s a m e s m a l l a m o u n t d u r i n g t h e s a m e p e r i o d (i.e. < 2 %).
498 R . M . K o c a n & M, L. L a n d o l t
DAY 1 EXPOSURE ~ p e r i v i t e i l i n e
fluid
15%
(pre-blostulo) ~ soc
f r y ~
excreted
2
%
DAY 15 EXPOSURE [
.p.erivitelline fluid 13%
DAY 15 EXPOSURE [ i f / / ~ " ~ @ . . ~ e x c r e t e d 9 % (eyed embryo)
perivitetline fluid 21%
~ e x c r e t e d 18%
DAY 25 EXPOSURE ~ x ' x ~ ~ i , ,~,,,....,...
(orgon0genesis)
Fig, 2. Distribution of benzo(a)pyrene in trout eggs from exposure to hatching, B(a)P was identified in three components of the egg/embryo complex: (1) excretions out of the egg during embryo development up to the time of hatching; (2) losses along with perivitelline fluid at the time of hatching; (3) a component bound to or dissolved in the embryo/yolk, Embryos exposed later in
development excreted considerably more of their B(a)P content than did those exposed early. Conversely, the amount present in the embryo/yolk was about 25 % greater in the early exposure group than in the late exposure group. Embryos exposed midway through development exhibited
intermediate values in all three categories
m e n t a g a i n r e l e a s e d m o r e l a b e l l e d c o m p o u n d at the t i m e of h a t c h i n g t h a n did those e x p o s e d early,
Sac fry c o n t i n u e d to e x c r e t e l a b e l l e d s o l u b l e c o m p o u n d s into the w a t e r t h r o u g h o u t yolk resorption. O n c e t h e yolk w a s t o t a l l y r e s o r b e d a n d t h e a b d o m i n a l s u t u r e l i n e closed (30 to 45 days post h a t c h i n g ) , n o l a b e l could b e d e t e c t e d i n the w a t e r i n w h i c h the fry w e r e kept.
H a t c h i n g
D e v e l o p i n g f i s h e m b r y o s e x p o s e d t o b e n z o ( a ) p y r e n e 499
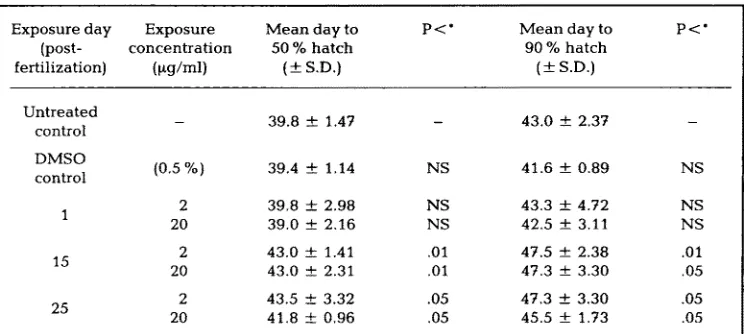
Table 2. M e a n h a t c h i n g day of s t e e l h e a d trout e g g s t r e a t e d with b e n z o ( a ) p y r e n e on days 1, 15 a n d 25 post fertilization
Exposure day Exposure M e a n day to P <: * M e a n day to
(post- concentration 50 % hatch 90 % hatch
fertilization) (ptg/ml) ( ± S,D.) ( ± S.D.)
p < *
Untreated control DMSO control
15
25
39.8 ± 1,47 - 43.0 ± 2.37
(0.5 %) 39.4 -- 1.14 NS 41.6 + 0.89 NS
2 39.8 ± 2.98 NS 43.3 ± 4.72 NS
20 39.0 ± 2.16 NS 42.5 ± 3.11 NS
2 43.0 ± 1.41 .01 47.5 ± 2.38 .01
20 43.0 ± 2.31 .01 47.3 ± 3.30 .05
2 43.5 ± 3.32 .05 47.3 ± 3.30 .05
20 41.8 ± 0.96 .05 45.5 ± 1.73 .05
* ANOVA: 100 e g g s / r e p l i c a t e ; controls = 6 replicates; t r e a t e d = 4 replicates; DMSO = 2 repli- cates; NS = not significant (P >0.25)
n o t e d a s a r e s u l t of e x p o s u r e to B(a)P, e v e n t h o u g h s o m e e g g s w e r e e x p o s e d to c o n c e n t r a t i o n s a s h i g h a s 20 btg/ml, a c o n c e n t r a t i o n a b o u t 1000 t i m e s g r e a t e r t h a n w o u l d n a t u r a l l y b e e n c o u n t e r e d . B o t h B(a)P c o n c e n t r a t i o n s p r o d u c e d t h e s a m e h a t c h i n g p a t t e r n i n t h e i r r e s p e c t i v e t r e a t m e n t g r o u p .
Y o l k a n d l a r v a l t i s s u e
C o m p a r i s o n of t h e a m o u n t of l a b e l l e d c o m p o u n d p r e s e n t i n y o l k a n d l a r v a l t i s s u e a t t h e t i m e of h a t c h i n g s h o w e d t h a t t h e y o l k c o n t a i n e d t h e m a j o r i t y of t h e o r i g i n a l e g g c o n t e n t of l a b e l l e d B(a)P. T a b l e 3 s u m m a r i z e s t h e p e r c e n t d i s t r i b u t i o n of r a d i o a c t i v i t y
Table 3. Concentration of B(a)P in embryo a n d yolk i m m e d i a t e l y post h a t c h i n g Exposure
concentration (~g/ml)
N Exposure day (post-fertilization)
1 15 25
2 Embryo 5 1.3 _±_ 0.2" (6%)** 3,7 ± 1.4 (9%) 5.7 + 3 . 0 ( 1 6 % ) Yolk 5 19 _ 2,1 (94%) 38 +_ 9 . 9 ( 9 1 % ) 31 ± 2 . 5 ( 8 4 % )
20 Embryo 5 5.0 ± 1.3(10%) 18 ± 3.2 (9%) 26 -- 4 . 5 ( 2 2 % )
Yolk 5 44 ± 3.9 (90%) 183 ± 33 (91%) 93 - 30 (78%)
* Values r e p r e s e n t n a n o g r a m s (ng) of B(a)P b o u n d to or dissolved in e m b r y o n i c tissues or yolk ( ± S,D,}
* * Values in ( ) r e p r e s e n t % of B(a)P in larvae or yolk relative to the total a m o u n t of B(a)P in
500 R . M . Kocan & M. L. L a n d o l t
l e v e l s d e t e c t e d i n yolk a n d larval tissue at the t i m e of h a t c h i n g . It is a p p a r e n t from these d a t a that those e m b r y o s w h i c h w e r e e x p o s e d e a r l i e r d u r i n g i n c u b a t i o n a n d excreted less B(a)P t h a n did those e x p o s e d later, r e t a i n e d the extra l a b e l l e d m a t e r i a l i n the yolk (90 to 94%) as c o m p a r e d to those e x p o s e d o n d a y 25 w h i c h r e t a i n e d m u c h less (78 to 84%). From this it is a p p a r e n t that the l a r v a e w h i c h w e r e e x p o s e d early a c c u m u l a t e more B(a)P i n t h e i r tissues at t h e t i m e of h a t c h i n g t h a n those e x p o s e d later i n d e v e l o p m e n t . By the t i m e the yolk h a d b e e n fully r e s o r b e d the fry from e a c h e x p o s u r e group c o n t a i n e d the s a m e l e v e l of b o u n d 14C, r e g a r d l e s s of the t i m e of t h e i r exposure.
M e t a b o l i s m / e x c r e t i o n
T a b l e 4 s u m m a r i z e s t h e r e s u l t s of o r g a n i c e x t r a c t i o n s of w a t e r s i n w h i c h sac fry from the v a r i o u s e x p o s u r e g r o u p s w e r e h e l d u n t i l t h e y h a d fully r e s o r b e d their yolk material. It is a p p a r e n t that b o t h s l i g h t l y p o l a r a n d c o n j u g a t e d forms of B(a)P m e t a b o l i t e s were e x c r e t e d into the water. T h e l a r g e a m o u n t of l a b e l l e d m a t e r i a l w h i c h r e m a i n e d i n the
Table 4. Percent water soluble 14C-B(a)P excreted by sac fry
Extraction procedure* Exposure day (post-fertilization)
1 15 25
Hexane (4 : 1) 90 93 97
Ethylacetate (4 : 1) 71 63 55
Ethylacetate (~-Glucuronidase/sulphatase) 52 43 36
* Extractions done on water collected daily from 10 sac fry from each treatment group over a 5-day period
w a t e r f o l l o w i n g o r g a n i c e x t r a c t i o n a n d ~ - g l u c u r o n i d a s e / s u l p h a t a s e d e c o n j u g a t i o n m a y r e p r e s e n t g l u t a t h i o n e c o n j u g a t e s of B(a)P m e t a b o l i t e s .
T h e r e a p p e a r s to b e a r e l a t i o n s h i p b e t w e e n the t i m e of e x p o s u r e d u r i n g d e v e l o p - m e n t a n d t h e a m o u n t of s o l u b l e m e t a b o l i t e s s u b s e q u e n t l y e x c r e t e d b y the larva d u r i n g yolk resorption, w i t h those e x p o s e d later e x c r e t i n g l a r g e r q u a n t i t i e s of s o l u b l e a n d c o n j u g a t e d m a t e r i a l . By the t i m e the yolk is fully resorbed, however, all of these differences d i s a p p e a r . Both e x p o s u r e g r o u p s (2 ~g a n d 20 ~g) e x h i b i t e d the s a m e p a t t e r n s of solubility.
T e r a t o g e n e s i s
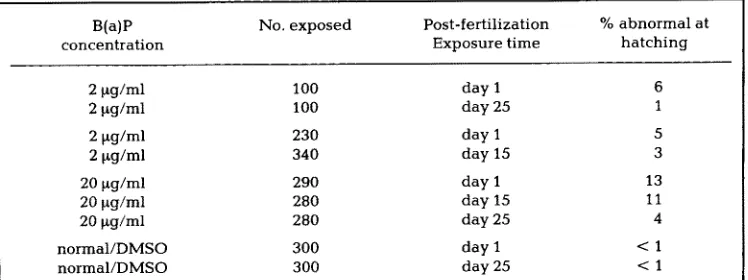
A m o n g the control fish ( u n e x p o s e d a n d D M S O treated), less t h a n 1% of the 600 a n i m a l s e x h i b i t e d a n y type of p h y s i c a l defect b y the t i m e t h e y h a d totally r e s o r b e d their yolk. T h e t r e a t e d groups, however, all s h o w e d i n c r e a s e d p h y s i c a l defects b y this time w i t h the m a j o r i t y a p p e a r i n g i n those a n i m a l s t r e a t e d early d u r i n g their d e v e l o p m e n t . T a b l e 5 s u m m a r i z e s the data from the t e r a t o g e n e s i s e v a l u a t i o n .
D e v e l o p i n g fish e m b r y o s e x p o s e d to b e n z o ( a ) p y r e n e 501
Table 5. Teratogenic effects on fish embryos following exposure to benzo(a)pyrene.
B(a)P No. exposed Post-fertilization % abnormal at
concentration Exposure time hatching
2 ~g/ml 100 day 1 6
2 ~g/ml 100 day 25 1
2 ~tg/ml 230 day 1 5
2 ~g/ml 340 day 15 3
20 ~tg/ml 290 day 1 13
20 ~g/ml 280 day 15 I 1
20 ~g/ml 280 day 25 4
normal/DMSO 300 day I < 1
normal/DMSO 300 day 25 < 1
Day 1 = pre-blastula; day 15 ~ eyed stage (1st eye pigment); day 25 = late organogenesis; day 35 = hatching
D I S C U S S I O N
U n l i k e i n f e c t i o u s d i s e a s e s , g e n e t i c a n d m e t a b o l i c d i s e a s e s a r e u s u a l l y d e t e c t a b l e o n l y w h e n c o m p a r a t i v e m e a s u r e m e n t s c a n b e m a d e from a s i n g l e i n d i v i d u a l , a n d a w e l l e s t a b l i s h e d n o r m a l b a s e l i n e for t h o s e m e a s u r e m e n t s is a v a i l a b l e for c o m p a r i s o n .
T h e data w e h a v e p r e s e n t e d h e r e d e m o n s t r a t e that d r a m a t i c n o n l e t h a l d e v i a t i o n s from the n o r m c a n o c c u r in e m b r y o s f o l l o w i n g e x p o s u r e to a k n o w n e n v i r o n m e n t a l c o n t a m i n a n t . S i n c e no s i g n i f i c a n t m o r t a l i t y o c c u r r e d in t h e test a n i m a l s , a n d p r e c i s e m e a s u r e m e n t s w e r e n e e d e d on l a r g e n u m b e r s of i n d i v i d u a l s w i t h s i m i l a r g e n e t i c b a c k g r o u n d , it is u n l i k e l y t h a t t h e s e s a m e c h a n g e s w o u l d h a v e b e e n d e t e c t a b l e u n d e r n a t u r a l conditions. It is not c l e a r w h a t effect e a c h of t h e a l t e r a t i o n s w e o b s e r v e d w o u l d h a v e on the s u r v i v a l a n d r e p r o d u c t i v e p o t e n t i a l of t h e e x p o s e d p o p u l a t i o n . B a s e d on w h a t is k n o w n of s o m e of t h e e n z y m e s y s t e m s w h i c h w e r e a l t e r e d , t h e r e p r e s u m a b l y c o u l d b e d r a m a t i c effects at s o m e l a t e r p e r i o d in t h e o r g a n i s m ' s d e v e l o p m e n t .
502 R . M . K o c a n & M. L. L a n d o l t
S t e g e m a n , 1983), t h e r e b y i n c r e a s i n g t h e c a p a c i t y of the o r g a n i s m to excrete the foreign c o m p o u n d . T h o s e e m b r y o s w h i c h w e r e e x p o s e d as a p r e - b l a s t u l a w o u l d e n c o u n t e r B(a)P o n l y v i a t h e yolk, w h i c h is b e i n g resorbed, a n d p o s s i b l y n o t r e s p o n d t h e s a m e as those w h i c h r e c e i v e d total b o d y e x p o s u r e . A n o t h e r p o s s i b i l i t y is that different r e s p o n s e s result from e a r l y a n d late exposures, t h u s c a u s i n g e a c h to r e s p o n d d i f f e r e n t l y t h a n w o u l d a n u n e x p o s e d e m b r y o . W h i c h e v e r the case, if t h e s e differences i n m e t a b o l i c p r o c e s s i n g a n d e x c r e t i o n of B(a)P p e r s i s t i n t o later life, t h e y c o u l d alter the o r g a n i s m ' s a b i l i t y to deal w i t h s u b s e q u e n t e x p o s u r e to s i m i l a r c o m p o u n d s a n d its a b i l i t y to process e n d o g e n o u s c o m p o u n d s such as steroids.
As w e h a v e p o i n t e d out, the a g e of a n e m b r y o at the t i m e of e x p o s u r e to a x e n o b i o t i c i n f l u e n c e s t h e u l t i m a t e c a p a c i t y of the l a t e r l a r v a to e x c r e t e that c o m p o u n d d u r i n g yolk resorption. T h o s e e x p o s e d e a r l y excrete t h e c o m p o u n d m o r e slowly t h a n those e x p o s e d l a t e r i n d e v e l o p m e n t . W h e t h e r r a p i d e l i m i n a t i o n of x e n o b i o t i c s is d e s i r a b l e or not d e p e n d s on w h e t h e r the r e d u c t i o n i n b o d y b u r d e n of the c h e m i c a l o u t w e i g h s the rapid f o r m a t i o n of a c t i v a t e d i n t e r m e d i a t e c o m p o u n d s w h i c h h a v e the capacity to b i n d to p r o t e i n a n d n u c l e i c acid a n d t h e r e b y to alter e n z y m e function, DNA stability, a n d i n i t i a t e n e o p l a s i a .
S i n n h u b e r & W a l e s (1974) a n d W a l e s et al. (1978) o b s e r v e d that the e m b r y o ' s age at t h e t i m e of e x p o s u r e to a f l a t o x i n i n f l u e n c e d t h e d e v e l o p m e n t of t u m o r s l a t e r i n life. This c o u l d b e e x p l a i n e d b y different rates of m e t a b o l i s m a n d e x c r e t i o n b y the different ages of t h e e m b r y o s at t h e t i m e of exposure.
It is c l e a r that d i f f e r e n c e s i n b o d y b u r d e n , excretory rate a n d the form of the c o n j u g a t e d c o m p o u n d s e x t e n d s b e y o n d h a t c h i n g a n d t h r o u g h yolk resorption. It has yet to b e d e m o n s t r a t e d , however, w h e t h e r t h e s e differences persist w h e n the o r g a n i s m is e x p o s e d to s i m i l a r c o m p o u n d s as it matures, or w h e t h e r t h e s e e n z y m a t i c a l t e r a t i o n s u l t i m a t e l y affect the i n d i v i d u a l t h r o u g h b e h a v i o u r a l m o d i f i c a t i o n or h o r m o n a l i m b a - l a n c e s (Lorz et al., 1979).
T h e c h a n g e s w e o b s e r v e d i n h a t c h i n g d y n a m i c s m a y l i k e w i s e b e r e l a t e d to altera- tions i n e n z y m e s y s t e m s w h i c h p a r t i a l l y control h a t c h i n g . It h a s b e e n w e l l e s t a b l i s h e d (Blaxter, 1969) that h a t c h i n g e n z y m e s are r e s p o n s i b l e for the o n s e t of h a t c h i n g a n d e g g m e m b r a n e d i s r u p t i o n . It is also k n o w n that a v a r i e t y of e n v i r o n m e n t a l factors, i n c l u d i n g p o l l u t i o n , c a n alter t h e s e e n z y m e s a n d affect the h a t c h i n g d y n a m i c s of a species. Therefore, if p o l y c y c l i c a r o m a t i c s such as B(a)P c a n alter h a t c h i n g g l a n d e n z y m e s , it c o u l d e x p l a i n the c h a n g e s w e o b s e r v e d .
D e v e l o p i n g f i s h e m b r y o s e x p o s e d to b e n z o ( a ) p y r e n e 503 B(a)P o f t e n s u f f e r e d f r o m d e v e l o p m e n t a l a b n o r m a l i t i e s a s d i d t h o s e e m b r y o s p r o d u c e d b y f e m a l e s w h i c h h a d b e e n e x p o s e d to B(a)P p r i o r t o e g g l a y i n g .
A l t h o u g h i n f e c t i o u s d i s e a s e s a r e a s i g n i f i c a n t f a c t o r i n f l u e n c i n g t h e h e a l t h a n d s u r v i v a l of a q u a t i c o r g a n i s m s , t h e r e is a m o r e i n s i d i o u s f o r m of d i s e a s e w h i c h r e s u l t s f r o m e x p o s u r e t o s u b l e t h a l t o x i c s u b s t a n c e s i n t h e e n v i r o n m e n t . T h o s e o r g a n i s m s w h i c h a r e e x p o s e d , a n y t i m e f r o m f e r t i l i z a t i o n to s e x u a l m a t u r i t y , r u n t h e r i s k of b e c o m i n g g e n e t i c a l l y d a m a g e d , a n d t h e r e b y b e i n g l e s s fit t o r e s p o n d t o o t h e r f o r m s of e n v i r o n m e n - t a l i n s u l t s , a n d m o r e s e r i o u s l y , h a v i n g a r e d u c e d c a p a c i t y t o p r o d u c e n o r m a l h e a l t h y o f f s p r i n g . E f f e c t s s u c h a s i n d u c t i o n a n d / o r s u p p r e s s i o n of e x i s t i n g e n z y m e s y s t e m s d u r i n g e m b r y o n i c d e v e l o p m e n t a l s o p o s e t h e t h r e a t of c o m p r o m i s i n g a n i n d i v i d u a l ' s a b i l i t y to c o p e w i t h l a t e r e x p o s u r e t o e n v i r o n m e n t a l c o n t a m i n a n t s .
Acknowledgements: This work w a s s u p p o r t e d in part by grants from the NIEHS (ES-02190-02), EPA
(R-810057-1), NOAA (OMPA-NA 80 RAD 00053) a n d NOAA (MESA-04-78-B01-13). The authors also a c k n o w l e d g e the assistance of K. Sabo, V. Liguori a n d L. Ginno.
L I T E R A T U R E C I T E D
Bend, J. R. & James, M. O., 1978. Xenobiotic m e t a b o l i s m in m a r i n e a n d f r e s h w a t e r species. In: Biochemical and biophysical p e r s p e c t i v e s in m a r i n e biology. Ed. by D. C. Malins & J. R. Sargent. Acad. Press, N e w York, 4, 128-132.
Binder, R. L. & Stegeman, J. J., 1980. Induction of aryl h y d r o c a r b o n hydroxylase activity in e m b r y o s of an estuarine fish. - Biochem. Pharmac. 29, 949-951.
Binder, R. L. & Stegeman, J. J., 1983. Basal levels a n d induction of h e p a t i c aryl h y d r o c a r b o n hydroxylase activity d u r i n g the e m b r y o n i c p e r i o d of d e v e l o p m e n t in brook trout. - Biochem. Pharmac. 32, 1324-1327.
Blaxter, J. H. S., 1969. Development: Eggs a n d larvae. In: Fish physiology. Ed. by W. S. Hoar & D. J. Randall. Acad. Press, N e w York, 3, 178-252.
Hose, J. E., H a n n a h , J. B., Landolt, M. L., Miller, B. S., Felton, S. P. & Iwaoka, W. T., 1981. U p t a k e of benzo(a)pyrene by g o n a d a l tissue of flatfish (pleuronectidae) a n d its effects on s u b s e q u e n t e g g development. - J. Toxicol. environ. Hlth 7, 991-1000.
Hose, J. E,, Hannah, J. B., DiJulio, D., Landolt, M. L., Miller, B. S., Iwaoka, W. T. & Felton, S. P., 1982. Effects of benzo(a)pyrene on early d e v e l o p m e n t of flatfish. - Archs environ. Contain. Toxicol. 11, 167-171.
Iwaoka, W. T., Landolt, M. L., Pierson, K. B., Felton, S. P. & Abolins, A., 1979. Studies on aryl hydrocarbon hydroxylase, polycyclic h y d r o c a r b o n c o n t e n t a n d e p i d e r m a l tumors of flatfish. In: Animals as monitors of e n v i r o n m e n t a l pollutants. Ed. by S. W. Nielsen, G. Migaki & D. G. Scarpelli. Natn. Acad. Sci., Washington, 85-93.
Kligerman, A. D., 1979. Induction of sister c h r o m a t i d e x c h a n g e s in the central m u d m i n n o w following in vivo exposure to m u t a g e n i c agents. - Mutat. Res. 64, 205-217.
Kligerman, A. D. & Bloom, S. E., 1976. Sister chromatid differentiation a n d e x c h a n g e s in adult m u d m i n n o w s (Umbra limi} after in vivo e x p o s u r e to 5-bromodeoxyuridine. - C h r o m o s o m a 56,
101-109.
Kocan, R. M., Landolt, M. L., Bond, J. & Benditt, E, P., 1981. In vitro effect of some m u t a g e n s / carcinogens on cultured fish cells. - A r c h s environ. Contain. ToxicoL 10, 663-671.
Kocan, R. M., Landolt, M. L & Sabo, K. M., 1982. A n a p h a s e aberrations: A m e a s u r e of genotoxicity in m u t a g e n - t r e a t e d fish cells. - Environ. M u t a g e n . 4, 181-189.
Laale, H. W., 1981. Teratology a n d early fish d e v e l o p m e n t . - Am. Zool. 21, 517-533.
5 0 4 R . M . K o c a n & M . L. L a n d o r
s e l e c t e d h e r b i c i d e s o n s m o l t i n g of coho s a l m o n . U.S. E n v i r o n m e n t a l Protection A g e n c y , Corval- lis, O r e g o n (PrepubL Rep.).
Lucier, G. W., 1981. D e v e l o p m e n t a l a s p e c t s of d r u g c o n j u g a t i o n . In: D e v e l o p m e n t a l toxicology. Ed. by C. A. K i m m e l l & J. B u e l k e - S a m , R a v e n Press, N e w York, 101-114.
Nelson, J. S., 1977. E v i d e n c e of g e n e t i c b a s i s for a b s e n c e of t h e p e l v i c s k e l e t o n in brook stickleback, Culaea inconstans, a n d n o t e s on t h e g e o g r a p h i c a l d i s t r i b u t i o n a n d origin of t h e loss. - J. Fish. Res. Bd Can. 34, 1314-1320.
P e d e r s o n , M. G., H e r s h b e r g e r , W. K., Z a c h a r i a h , P. K. & J u c h a u , M. R., 1976. H e p a t i c b i o t r a n s f o r m a - tion of e n v i r o n m e n t a l x e n o b i o t i c s in six s t r a i n s of r a i n b o w trout (Salmo gairdneri). - J. Fish. Res. Bd Can. 33, 666-675.
Schr6der, J. H., 1969. I n h e r i t a n c e of r a d i a t i o n - i n d u c e d s p i n a l c u r v a t u r e s in t h e g u p p y , Lebistes r e t i c u l a t u s . - Can. J. G e n e t . Cytol. 11, 937-947.
Schr6der, J. H., 1973. T e l e o s t s as a tool in m u t a t i o n r e s e a r c h . In: G e n e t i c s a n d m u t a g e n e s i s of fish. Ed, b y J. H. Schr6der. Springer, Berlin, 91-99.
S i n n h u b e r , R. O. & W a l e s , J. H. 1974. A f l a t o x i n B 1 h e p a t o c a r c i n o g e n i c i t y in r a i n b o w trout embryos.
- F e d n Proc. F e d n Am. Socs exp. Biol. 33, 247.
S t e g e m a n , J. J., 1981, P o l y n u c l e a r a r o m a t i c h y d r o c a r b o n s a n d their m e t a b o l i s m in the m a r i n e e n v i r o n m e n t . In: Polycyclic h y d r o c a r b o n s a n d cancer. Ed. b y H. V. G e l b o i n & P. O, P. Ts'o. Acad. Press, N e w York, 3, 1-60.