1535-9778/11/$12.00 doi:10.1128/EC.05215-11
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Cell Cycle Regulators Interact with Pathways That Modulate
Microtubule Stability in
Saccharomyces cerevisiae
䌤
Aya Shohat-Tal and Dan Eshel*
Department of Biology, Brooklyn College, The City University of New York, 2900 Bedford Avenue, Brooklyn, New York 11210, and The City University of New York Graduate Center, 365 Fifth Avenue, New York, New York 10016
Received 19 August 2011/Accepted 19 October 2011
The integrity of mitosis is dependent upon strict regulation of microtubule stability and dynamics. Although much information has been accumulated on regulators of the microtubule cytoskeleton, our knowledge of the specific pathways involved is still limited. Here we designed genetic screens to identify regulators of microtu-bule stability that are dispensable in the wild type yet become essential under microtumicrotu-bule-disrupting
condi-tions. We found that the transcriptional cofactor Swi6p and activator Swi4p, as well as the G2/M-specific cyclin
Clb2p, are required in a microtubule-destabilizing environment. Swi6p and Swi4p can combine as a transcriptional complex, called the SBF complex (SBF for Swi4/6 cell cycle box [SCB]-binding factor) that
is functionally homologous to the metazoan DP1/2-E2F complex and that controls the G1/S transition
through the genes it regulates. We show that Swi6p’s contribution to microtubule stability can be either dependent or independent of the SBF complex. The SBF-dependent pathway requires downregulation of SBF complex levels and may thereby reroute the transcriptional program in favor of greater microtubule stability. This pathway can be triggered by overexpression of Fcp1p, a phosphatase in the general
transcription machinery, or by expression of an allele ofSWI6that is associated with reduced
transcrip-tion from SBF-controlled promoters. The SBF-independent pathway is activated by a constitutively nuclear allele of Swi6p. Our results introduce novel roles in microtubule stability for genes whose participation in the process may be masked under normal conditions yet nonetheless acquire a dominant role when microtubule stability is compromised.
The fidelity of chromosome segregation in mitosis is essen-tial for the accurate propagation of genetic information to daughter cells, and it is dependent upon timely, coordinated
changes in the microtubule (MT) cytoskeleton. In
Saccharo-myces cerevisiae, chromosome segregation is achieved in four microtubule-dependent steps. The first step is spindle assem-bly, which involves the migration of duplicated spindle pole bodies (SPBs) to form a bipolar spindle. This process requires the plus-end-directed activity of the kinesin-like motors of the BimC family, Cin8p and Kip1p, that cross-link and slide
an-tiparallel microtubules of the spindle midzone at the G2/M
transition (27, 47). The second step is orientation of the mitotic spindle at the site of cytokinesis, which is mediated by cy-toplasmic MTs and requires their transient, dynamic inter-actions with the cell cortex through dynein-dependent (51) and dynein-independent (50, 4) pathways. The third step is chromosome movement along kinetochore MTs through their depolymerization in anaphase A (46), and the fourth step is complete chromosome separation in the process of anaphase B spindle elongation. This last process is powered by Cin8p and Kip1p that cross-link and push apart polar MTs, by polymerization of the same MTs in the midzone driven by a poleward flux of tubulin subunits, and by a pulling force gen-erated by dynein on cytoplasmic MTs (49).
These changes in the MT architecture are dependent on
tight regulation on different levels. Most directly, motor pro-teins and MT-associated propro-teins (MAPs) influence MT po-lymerization, stability, and dynamics (31, 18, 7, 29, 11), thereby affecting processes the specific MT state facilitates. For
exam-ple, the temperature-sensitive double motorcin8-3 kip1⌬
mu-tant is sensitive to various MT-destabilizing drugs. At elevated
temperatures, spindle assembly at G2/M is compromised (20,
42, 49). Introduction of thetub2-402allele, which
hyperstabi-lizes MTs (28), suppressed the sensitivities both to elevated temperatures and to MT-destabilizing drugs that are associ-ated with this double mutant (39), suggesting destabilized MTs in this background.
Another level of regulation is by upstream cyclin/Cdk complexes whose periodic expression drives specific cell
cy-cle events. Late in G1phase, the Cln3p-Cdc28p protein kinase
complex activates two transcription complexes, the MBF com-plex (MBF for MluI cell cycle box [MCB]-binding factor) and the SBF complex (SBF for Swi4/6 cell cycle box [SCB]-binding factor), and these in turn promote the transcription of a num-ber of genes important for budding and DNA synthesis (10,
26). At G2/M, the Clb2p-Cdc28p complex represses the activity
of SBF, returning the expression of SBF-regulated genes to low levels (1). MT regulation by cyclin/cdk complexes may manifest indirectly: Clb2p-Cdc28p, for example, contributes to the sta-bility or localization of motors posttranslationally (8, 12). It is therefore not surprising that certain Clb/Cdk mutants share
the same phenotype exhibited bycin8-3 kip1⌬double mutants
grown at the elevated temperature, being unable to assemble a bipolar spindle due to a failure to segregate duplicated SPBs (20, 42, 49). These reports reinforce the ties that exist between upstream and downstream MT regulators.
* Corresponding author. Mailing address: Department of Biology, Brooklyn College, The City University of New York, 2900 Bedford Avenue, Brooklyn, NY 11210. Phone: (718) 951-5000, ext. 2015. Fax: (718) 951-4826. E-mail: [email protected].
䌤Published ahead of print on 28 October 2011.
1705
on September 8, 2020 by guest
http://ec.asm.org/
In wild-type cells under normal conditions, phenotypes are often not manifested because other proteins or pathways act
redundantly. A general example isKSS1, a gene whose
de-letion in wild-type cells has no phenotypic consequence on the mating pathway and therefore was originally thought not to be involved in the process. It was later shown that it can fill in for another protein, Fus3p, which is functionally re-dundant with Kss1p, when Fus3p is not present (48). Simi-larly, in this study we aimed to identify proteins that under normal conditions have no apparent role in MT stability, but when the stability of MTs is compromised, their involvement becomes essential.
We previously conducted a genetic screen to identify pro-teins that when overexpressed can correct the temperature sensitivity and the MT destabilization phenotype associated with cin8-3 kip1⌬ cells (54). Here we describe pathways by which Fcp1p, one of the overexpression suppressors, increases MT stability in this background. The underlying mechanisms were elucidated through the identification of proteins that Fcp1p’s effect is dependent upon or that can substitute for it altogether. On the basis of these results, we propose novel pathways that regulate the stability of microtubules.
MATERIALS AND METHODS
Strains and media.Saccharomyces cerevisiae strains used in this study are listed in Table 1. All strains are either S288C derivatives or were backcrossed at least seven times to an S288C background. Strain DEY2169 was constructed by crossing strain MAY2169 with strain YNN282 (Yeast Genetic Stock Center, University of California, Berkeley), followed by tetrad dissection and analysis as previously described (32). Medium preparation and yeast genetic and transfor-mation techniques were essentially as previously described (32). Sensitivity to
thiabendazole (TBZ) was tested on yeast extract-peptone-dextrose (YPD) agar medium to which the desired amount of TBZ (MP Biomedicals Inc.) was added from 10 mg/ml stock in dimethylformamide (DMF). Serial dilution experiments were repeated three times.
Screen for multicopy suppressors.Details of the screen for multicopy sup-pressors were previously described (54). In brief, after transformation ofcin8-3 dyn1⌬cells with the YEp24-based yeast genomic library, colonies that grew at both the permissive temperature (26°C) and the nonpermissive temperature (35°C) upon replica plating were picked. Plasmids were extracted from these colonies, the genomic insert containing the suppressor was sequenced, and the gene responsible for the suppression was isolated. One of these suppressors, the FCP1gene, was subcloned into the XbaI and SmaI sites of the 2m plasmid YEplac112 and was used to transformcin8-3 kip1⌬cells.
Secondary screen with a Tn7insertional library.cin8-3 kip1⌬double mutants carrying the YEplac112-FCP1plasmid were transformed with a NotI-digested Tn7-derived insertional library (41). Gene disruptions were generated by homol-ogous recombination as the truncated genomic fragments replaced their native chromosomal locus. In ourcin8-3 kip1⌬background, such transformations gen-erated triple mutants. Cells containing the transposon were grown at 26°C, replica plated, and incubated at the nonpermissive temperature, 35°C. Since all cells originally grew at the nonpermissive temperature (due toFCP1 overexpres-sion), colonies that ameliorated this effect subsequent to insertional mutagenesis and grew at 26°C but not at 35°C were selected for further analysis.
Identification and confirmation of transformants.To identify genes that ame-liorate the effect ofFCP1overexpression, we isolated genomic DNA from col-onies that lost the ability to grow at the nonpermissive temperature, as described previously (32). Thermal asymmetric interlaced (TAIL) PCR (24) was used to amplify flanking sequences adjacent to the insertions. Degenerate PCR primers were as described previously (24), and specific primary and nested primers were designed on the basis of the inserted Tn7sequences. Sequence data from both ends of the insert were compared against the yeast genome database to deter-mine the identification of the genomic sequences it contains. DNA from trans-formant colonies was subjected to Southern blot analysis in order to exclude multiple Tn7insertions.
Construction of triple mutants.Deletion strains from theSaccharomyces De-letion Project were propagated on YPD plates containing 200g/ml Geneticin TABLE 1. Saccharomyces cerevisiaestrains and plasmids used in this study
Strain or plasmid Genotype Source or reference
Strains
MAY589 MATahis3-⌬200 leu2-3,112 ura3-52 ade2-101 M. A. Hoyt
MAY2169 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 M. A. Hoyt
YNN282 MAT␣his3-⌬200 ura3-52 ade2-101 lys2-801 trp1⌬ YGSCa
DEY2169 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬ This study DEY3001 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬swi6::KanMX4 This study DEY3002 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬swi4::KanMX4 This study DEY3003 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬mbp1::KanMX4 This study DEY3004 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬cln1::KanMX4 This study DEY3005 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬cln2::KanMX4 This study DEY3006 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬cln3::KanMX4 This study DEY3007 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬clb1::KanMX4 This study DEY3008 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬clb2::KanMX4 This study DEY3009 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬clb3::KanMX4 This study DEY3010 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬clb4::KanMX4 This study DEY3011 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬clb5::KanMX4 This study DEY3012 MATacin8-3 kip1::HIS3 his3-⌬200 leu2-3,112 ura3-52 ade2-101 trp1⌬clb6::KanMX4 This study DEY300 MATahis3-⌬200 leu2-3,112 ura3-52 ade2-101 swi6::KanMX4 This study DEY301 MATahis3-⌬200 leu2-3,112 ura3-52 ade2-101 swi4::KanMX4 This study DEY302 MATahis3-⌬200 leu2-3,112 ura3-52 ade2-101 clb2::KanMX4 This study
Plasmids
YEplac112 2mTRP1 New England Biolabs
pRS315 LEU2 CEN ATCC
pMA1208 CIN8 LEU2 CEN M. A. Hoyt
YEplac112-FCP1 FCP12mTRP1 This study
BD1265 SWI6 LEU2 CEN L. Breeden
BD1436 swi6-S160D LEU2 CEN L. Breeden
BD1651 swi6-S160A LEU2 CEN L. Breeden
aYeast Genetic Stock Center, University of California, Berkeley, CA.
on September 8, 2020 by guest
http://ec.asm.org/
(Gibco, Invitrogen). DNA was isolated from these strains as described above, and the genes containing the KanMX4 cassettes were PCR amplified using the primers shown in Table 2. PCR products containing the deletion cassettes were verified by gel electrophoresis and transformed into haploidcin8-3 kip1⌬double mutants carrying the 2m plasmid YEplac112-FCP1. DNA was subsequently purified from Geneticin-resistant triple mutants, and a confirmation primer (a few hundred base pairs upstream or downstream of the deletion) together with one of the original primers (Table 2) was used to confirm the correct integration. Immunoblots.Ten-milliliter log-phase cultures were centrifuged, and pellets were resuspended in 1 ml ice-cold Tris-EDTA (TE) buffer. Total protein lysate was prepared by vortexing cells with glass beads in 50l yeast extraction buffer (0.6% SDS and 10 mM Tris [pH 7.4] with the addition of 1g/ml leupeptin [Sigma], 2g/ml aprotinin [Sigma], 1g/ml pepstatin [Sigma], and 1 mM phen-ylmethylsulfonyl fluoride [PMSF] [Sigma]). Sample buffer (50l) (125 mM Tris-HCl, 20% glycerol, 4.1% SDS, 4%-mercaptoethanol) was then added, the solution was spun, and the supernatant was boiled for 5 min. A 5-l sample was mixed with an equal volume of Laemmli Sample Buffer (Bio-Rad) and loaded on 4 to 12% gradient NuPAGE Novex Bis-Tris minigel (Invitrogen), according to the manufacturer’s instructions. EZ-Run prestained protein marker (Fisher Sci-entific) was loaded as well. Western blotting was performed as described previ-ously (53) using Hybond enhanced chemiluminescence (ECL) nitrocellulose membrane (Amersham Biosciences). Rabbit anti-Swi6 antibody (a gift from L. Breeden) and peroxidase-linked goat anti-rabbit secondary antibodies (GE Healthcare) were both used at a 1:5,000 dilution. Rabbit anti-Swi4 antibody (a gift from B. Andrews) and peroxidase-linked goat anti-rabbit secondary antibod-ies were used at a dilution of 1:5,000 and 1:10,000, respectively. Mouse anti-3-phosphoglycerate kinase (anti-PGK) antibody (Invitrogen) was used as a control to normalize the results. Peroxidase-linked anti-mouse IgG secondary antibody (GE Healthcare) was used at a 1:5,000 dilution. To visualize the results, blots were developed using Amersham’s ECL detection reagents (GE Healthcare) in accordance with the manufacturer’s instructions and detected using Fujifilm Imager LAS-4000. Fluorescence quantification and analysis were done using Fujifilm Multi Gauge software. Western blots were repeated three times.
Statistical analysis.Data were expressed as means⫾standard errors of the means (SEM). Data were analyzed using analysis of variance coupled with Holm-Sidak test for multiple pairwise comparison. Data analysis was performed using SigmaStat (Chicago, IL).Pvalues of⬍0.05 were considered statistically significant.
RESULTS
Fcp1p is a strong suppressor of the temperature and TBZ
sensitivities associated withcin8-3 kip1⌬cells. FCP1 on an
overexpression vector was identified in our original screen (54) as a strong suppressor of the temperature and thiabendazole
(TBZ) sensitivities ofcin8-3 kip1⌬mutants. It restored growth
at the restrictive temperature (35°C) and conferred resistance to the MT-destabilizing drug TBZ to levels exceeding that of
the isogenic wild-type strain (Fig. 1A and B).cin8-3 kip1⌬cells
are also sensitive to the MT-destabilizing drug benomyl and its derivative methyl-benzimidazole-2-yl carbendazim (MBC)
(39) (data not shown). FCP1 overexpression rendered both
double mutant and wild-type cells more resistant to these latter drugs (not shown). TBZ, however, was the preferred drug for this study, as it showed pronounced differences in the growth profiles for various mutations in Swi6p, a protein that is the primary focus of this work.
Fcp1p (transcription factor IIF [TFIIF]-stimulated C-termi-nal domain [CTD] phosphatase) is a phosphatase in the gen-eral transcription machinery. It specifically dephosphorylates Ser2 of the tandem heptapeptide repeat Tyr1-Ser2-Pro3-Thr4-Ser5-Pro6-Ser7 in the C-terminal domain of the largest subunit (Rbp1p) of RNA polymerase II (RNAPII) (36). This con-served heptapeptide repeats 26 times in yeast and up to 52 times in metazoan CTDs (13). Upon transcriptional termina-tion, Fcp1p-mediated CTD dephosphorylation is required to recycle the polymerase at the end of each round of
transcrip-TABLE 2. Primers used to amplify the deletion cassettes from the yeast deletion strains Gene Forward primer used to amplify the deletion cassette Reverse primer used to amplify the deletion cassette Confirmation primer a SWI6 TTGCGACCGATCTTCACT GATTTTGAGCGACGCGAAAT TCATGTTGGATTTTAGGCAGG (rev) SWI4 TGTCACTACAAATTATTG GAAACGAGAAGAAGGATC GTCTGAGAGAACAAAGCCAA (rev) MBP1 ATAGTATCGCTGGCGTTTG TTGGAAGGTGTCACTGTCTTG ATATCACTGGACGTGGTTGTG (forw) CLN3 GAGTCCCAGTCGCAATCTCACATC ACGATAGTAATCGTATTTTAGGTTGTGC GAGGAAACAAGAGATAATTGGTCATC (rev) CLN1 TTTTGCTGACAATACATTTTAACGA CATTATTACTTACGATGGAAAAGCG TTAGAGGATGGGTCGTCAG (rev) CLN2 TCGGTCTCTTCGTTCTTCC GAGCTTAGGATTTTACCACAACTGA TTTTCGCCGGTTGAGTGTATC (rev) CLB1 TTCTAGGCAATTTTGGTTGTATTGT CATGGTTTATTTCAAGGTTTTTGAC AGGACACGATCGAGATTCCC (forw) CLB2 GAATTGATACTTCTGAAAAGCGAAA TACTGATGCTTTTCTTTGTCTACCC GTGCCTTAGGGGGACTCAAACTTAA (forw) CLB3 CAGGAAGAATGAAAACATCTCAGTT ATTTGGTGGCAGTATATGAATGAGT GGTCACTAAAAATGATGGCG (rev) CLB4 GTTTATTCGGAGTTTCTCATGTGTT ATTGTCAAATAAAGATTCGGTCAAA AAGGTACATCTTCCCGCTATG (forw) CLB5 AAGACGCGCCCTTGATGGTACAAA TGCTTTCTTCTCATTTTTCATTTCT GGTAAAGAGCTAGTTCAACTG (rev) CLB6 ACACAGGACTTGGTTGTTTCTACTC ACACAGAAAACCAAAATCTCACTTC TTTGGTGAACTGGATAATCAAGATTC (rev) a rev, reverse; forw, forward.
on September 8, 2020 by guest
http://ec.asm.org/
tion, an action that primes it for the next transcription cycle (16).
In order to identify domains, and therefore a specific
func-tion of Fcp1p that is necessary for the suppression of thecin8-3
kip1⌬phenotype, we assayed the effects of overexpression of
variousfcp1mutant genes on our strain. We separately
gener-ated amino acid substitutions of the first and second conserved
aspartatic acids of the⌿⌿⌿DXDX(T/V)⌿⌿motif (where⌿
is any aromatic amino acid), which are central to catalysis (33). In vivo, mutation of either aspartate abrogates the ability to rescue the lethality associated with a chromosomal deletion of
FCP1, even though the mutant proteins are expressed similar
at levels to that of the wild-type protein (36). We also
gener-ated a deletion of the C-terminal region ofFCP1(amino acids
626 to 732), a region that was implicated in binding the RAP74 subunit of TFIIF and TFIIB, both of which regulate Fcp1p’s activity (37). This deletion abolishes the protein’s interaction with the RAP74 subunit of TFIIF and with TFIIB yet
main-tains normal CTD phosphatase functionin vitro(2).
We found that the integrity of both the phosphatase domain and the C-terminal region was necessary for the suppression of
the temperature sensitivity ofcin8-3 kip1⌬cells when FCP1
was overexpressed (Fig. 1C). These findings of the requirement of Ser2 dephosphorylation by Fcp1p and the C-terminal region of Fcp1p for suppression were further corroborated when we
overexpressedSSU72in the same strain and failed to observe
suppression of the mutant phenotype.SSU72codes for a
phos-phatase specific for Ser5 of the same consensus sequence of the CTD of RNAPII and lacks a functionally distinct C-terminal domain (40).
Deletion ofSWI6andSWI4 ameliorates the effect ofFCP1
overexpression on MT stability in the cin8-3 kip1⌬
back-ground.The results of the mutational analysis of Fcp1p led us
to conclude that the effect of FCP1 overexpression on MT
stability may be indirect, as it requires the normal function of Fcp1p in the context of general transcription. We therefore
hypothesized that FCP1 overexpression, through aberrant
transcription, triggers a putative pathway that results in greater MT stability. To test this possibility, we designed a secondary
screen with a Tn7insertional library (see Materials and
Meth-ods) to search for genes that are needed for the effect ofFCP1
overexpression in the cin8-3 kip1⌬ background to manifest.
Transposon-mediated truncation ofSWI6in thecin8-3 kip1⌬
[FCP1-2m] background restored the sensitivity of the double
mutant to the elevated temperature and exacerbated its sensi-tivity to the microtubule-destabilizing drug TBZ (data not shown). We confirmed these results with a targeted deletion of
SWI6 to verify that it is this specific deletion and not other
unforeseen elements of the screen that is responsible for
ame-liorating the effect ofFCP1overexpression on the temperature
and TBZ sensitivity ofcin8-3 kip1⌬cells (Fig. 2A and B).
Furthermore, to exclude the possibility that aSWI6deletion
independently introduces added temperature sensitivity to the
already compromised strain, we introduced to thecin8-3 kip1⌬
swi6⌬triple mutant cells wild-typeCIN8on a plasmid. A
res-cue of the mutant phenotype (Fig. 2A) suggested that the effect
of Swi6p on MT stability is pertinent specifically to thecin8-3
kip1⌬genetic background.
Figure 2B also demonstrates that the single swi6 deletion
increases the sensitivity of wild-type cells to TBZ to
approxi-mately the same degree as that of the cin8-3 kip1⌬ double
FIG. 1.FCP1overexpression suppresses the temperature sensitiv-ity and thiabendazole (TBZ) sensitivsensitiv-ity associated withcin8-3 kip1⌬
cells. (A and B) Aliquots (5l) of cultures at a density of 2⫻106 cells/ml, followed by three serial 10-fold dilutions, were spotted onto YPD medium and allowed to grow at the permissive (26°C) and non-permissive (35°C) temperature (A) or in the absence or presence of increasing concentrations of TBZ (B). WT, wild type. (C) Overexpres-sion of various mutants ofFCP1abolish the suppression. CD, C-ter-minal domain.
FIG. 2. Deletion ofSWI6 orSWI4 from thecin8-3 kip1⌬ back-ground abrogates the effect ofFCP1overexpression on the tempera-ture sensitivity and TBZ sensitivity of the strain. (A and B) Aliquots (5 l) of cultures at a density of 2⫻106cells/ml, followed by three serial 10-fold dilutions, were spotted onto YPD medium and allowed to grow at the permissive (26°C) and nonpermissive (35°C) temperature (A) or in the presence of 80g/ml TBZ (B).
on September 8, 2020 by guest
http://ec.asm.org/
mutant and thatFCP1 overexpression does not increase the
resistance to TBZ of theswi6single mutant. Thus,FCP1
over-expression may act through Swi6p to stabilize microtubules, as it does not bypass the need for Swi6p under microtubule-destabilizing conditions.
To investigate the role of Swi6p in our system, we assayed genes whose products physically interact with Swi6p for their
ability to phenocopy the effect of aSWI6deletion. Swi6p is a
transcription cofactor that forms heteromeric complexes with the DNA-binding proteins Swi4p and Mbp1p to mediate
tran-scription at the G1/S transition (26). It is the regulatory or
trans-activator subunit of these complexes. SBF (SCB-binding factor) is the complex containing Swi4p and Swi6p that
en-hances transcription of G1 cyclin genes via SCB elements
(Swi4/6 cell cycle box; CACGAAA). MBF (MCB-binding fac-tor) consists of Mbp1p and Swi6p. This complex enhances the transcription of S-phase cyclin genes as well as genes involved in DNA synthesis and repair. It acts via MCB elements (MluI cell cycle box; ACGCGTNA) and leads to the initiation of a complex transcriptional cascade required for coordinated cell cycle progression (10).
Because of the involvement of Swi6p in both these com-plexes, we specifically looked into the potential implication of these complexes in the suppression. To do this, we first
gen-eratedcin8-3 kip1⌬swi4⌬andcin8-3 kip1⌬mbp1⌬triple
mu-tants to see whether these deletions similarly block the effect of
FCP1 overexpression on the temperature sensitivity of our
strains.
As shown in Fig. 2A,cin8-3 kip1⌬swi4⌬cells, but notcin8-3
kip1⌬mbp1⌬cells, lost the ability to grow at the nonpermissive
temperature even in the presence of a multicopy plasmid
car-ryingFCP1. Growth ofcin8-3 kip1⌬swi4⌬cells at the elevated
temperature was restored only upon introduction of a
plasmid-borne wild-type copy ofCIN8, suggesting that aswi4deletion
does not independently introduce sensitivity to the strain. These results suggest that the SBF complex is a likely candi-date through which MT stability is conferred. Cells in which
swi4alone was deleted or cells in which swi4was deleted in
combination withcin8-3 kip1⌬are not sensitive to TBZ;
how-ever, the deletion does exacerbate the sensitivity of the double mutant to other microtubule-destabilizing drugs, such as beno-myl and its derivative MBC (not shown). The mechanisms of action of the three drugs employed in our screens are similar— they all work by sequestering tubulin subunits, preventing po-lymerization (14, 55). Variation in the sensitivities of different strains to different drugs was previously attributed to ATP-binding cassette (ABC) transporters that are known to be responsible for drug resistance in fungi (6, 43, 44).
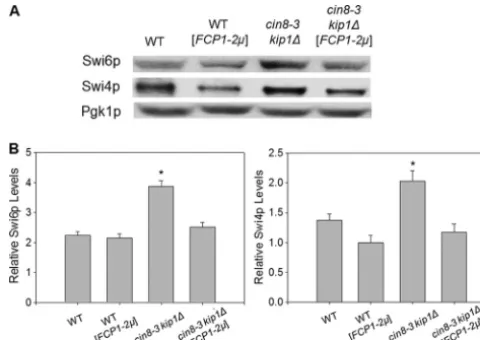
FCP1 overexpression downregulates Swi6p and Swi4p to
similar extents incin8-3 kip1⌬cells.We next performed
West-ern blot analysis to further investigate whether the SBF com-plex has a role in the suppression. We found that in wild-type
cells,FCP1overexpression does not significantly alter the
lev-els of Swi4p and Swi6p (Fig. 3).cin8-3 kip1⌬cells, however,
have elevated levels of these proteins compared to the wild
type. WhenFCP1is overexpressed in the double mutant, the
levels of the two proteins return to roughly wild-type levels. Furthermore, though Swi4p and Swi6p are present in different
amounts in cells, overexpression ofFCP1in thecin8-3 kip1⌬
background has the outcome of downregulating the proteins
similarly (1.61- and 1.55-fold, respectively) relative to double
mutants that do not overexpressFCP1.
These results demonstrate that the levels of Swi6p and Swi4p in the cell are affected when MT stability is compromised, as in the cin8-3 kip1⌬ background. This compromised condition may be sensed, resulting in a change in transcription patterns. More importantly, and in line with Fcp1p’s role in
transcrip-tion, the results suggest that in cin8-3 kip1⌬ cells, FCP1
overexpression leads to a reduction in the levels of the SBF complex specifically, as its components Swi4p and Swi6p are downregulated comparably. Taken together with the results shown in Fig. 2, the data reveal a correlation between SBF complex levels and MT stability and present the possibility that curtailing transcription from SBF promoters supports MT sta-bility.
Expression of the swi6-S160D allele, which results in
re-duced transcription from SBF-regulated promoters, rescues
the TBZ sensitivity ofcin8-3 kip1⌬swi6⌬cells.The results in
Fig. 2 and 3 that point to the SBF complex as an intermediary through which MT stability is reconciled led us to ask whether alteration of transcription from SBF promoters is needed for the suppression of the TBZ sensitivity of our mutants. To
examine this, we obtained two alleles ofSWI6, a constitutively
nuclear (swi6-S160A) allele, whose expression does not reveal
major changes in SBF-driven transcription, and a predominantly
cytoplasmic (swi6-S160D) allele, which was shown to result in a
marked reduction of transcription from SBF-regulated promoters (52). Protein expression from plasmids carrying these alleles was previously shown to be comparable to protein expression from a
plasmid carrying the wild-typeSWI6gene (52).
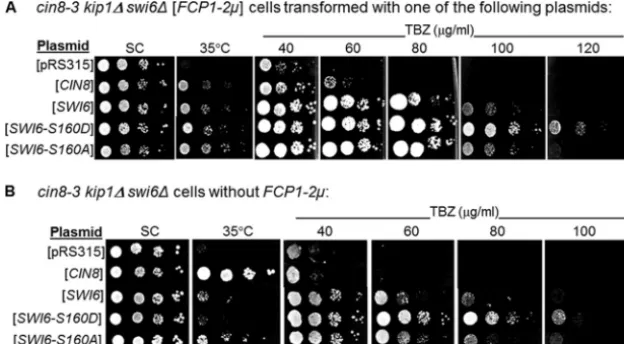
We expressed these alleles in the compromisedcin8-3 kip1⌬
swi6⌬[FCP1-2m] strain. As shown in Fig. 4A, introduction of
a wild-type copy ofSWI6on a centromeric plasmid resulted in
the predicted complementation of the temperature-sensitive
FIG. 3.FCP1overexpression incin8-3 kip1⌬cells correlates with a similar downregulation factor of Swi6p and Swi4p levels relative to cells not overexpressingFCP1.(A) Representative blot of three inde-pendent experiments showing protein expression of Swi6 and Swi4 relative to the Pgk1p control. (B) Densitometry analysis of Swi6 and Swi4 protein levels relative to the Pgk1p control. Data are expressed as means plus standard errors of the means (SEM) (error bars). Values that were significantly different (P⬍0.05) compared to the values for all other groups are indicated by an asterisk.
on September 8, 2020 by guest
http://ec.asm.org/
phenotype. Introduction of other alleles ofSWI6on a plasmid also resulted in complementation. When assayed for growth on
a medium containing TBZ, cin8-3 kip1⌬ swi6⌬ [FCP1-2m]
cells carrying wild-typeSWI6orswi6-S160Aon a plasmid
re-sulted in resistance to the microtubule-destabilizing drug at a
level roughly equivalent to that ofcin8-3 kip1⌬double mutants
overexpressingFCP1 (compare Fig. 4A with Fig. 1B).
Inter-estingly, cells carrying the Swi6-S160D allele demonstrated
even higher resistance to TBZ. This led us to suggest that an
additive effect may be in place when bothFCP1overexpression
and Swi6-S160D are present in thecin8-3 kip1⌬swi6⌬
back-ground, resulting in such high levels of resistance. If this is the
case, then theswi6-S160Dallele may be able to mediate
suf-ficient suppression in the absence ofFCP1overexpression.
To test this hypothesis, triple mutants were cured of the
multicopy plasmid carryingFCP1. Figure 4B reveals that while
plasmids carrying the wild-typeSWI6or itsS160Aversion no
longer supported growth at TBZ concentrations that
compro-mised the double mutant,swi6-S160D, although less resilient at
extremely high TBZ concentrations in the absence ofFCP1
overexpression, is nonetheless able to restore growth in the cells to a level similar to that of the double mutant
overex-pressingFCP1(compare Fig. 4B with Fig. 1B).
These results suggest that the effects of theseswi6alleles on
MT stability is independent ofFCP1overexpression and may
substitute for it to achieve the same suppression. Furthermore, because of its reported reduced transcription from SBF
pro-moters (52), the suppression caused by the S160D allele of
SWI6proves that not only is modulation of the levels of the
proteins that make up the SBF complex needed (as suggested in Fig. 3) but that a functional SBF complex must be in place
to exert its effect on transcription of the G1/S genes it is known
to regulate.
Expression of the constitutively nuclear swi6-S160A allele
causes resistance to elevated temperatures for cin8-3 kip1⌬
swi6⌬cells.In the presence ofFCP1overexpression, all alleles
ofSWI6tested have the capacity to complement the
temper-ature-sensitive phenotype (Fig. 4A). However, in the absence
ofFCP1overexpression, only theswi6-S160Aallele is able to
suppress the temperature sensitivity of our mutants (Fig. 4B). The fact that rescue of the temperature sensitivity in the
absence of FCP1 overexpression requires the S160A allele
while resistance to TBZ requires theS160Dallele ofSWI6is
explained as follows: when damage to microtubules is global (i.e., when both nuclear and cytoplasmic MTs are affected, as is the case when cells are grown in the presence of TBZ),
downregulating G1/S transcription from SBF-controlled
pro-moters through expression of theswi6-S160Dallele can
sup-press the MT defect. On the other hand, when the damage is
limited to spindle MTs, as is the case for cin8-3 kip1⌬ cells
grown at the elevated temperatures (20, 42, 49), the constitu-tive presence of Swi6p in the nucleus can cause some suppres-sion. There may be a dose effect in that Swi6p-S160D, which is mostly cytoplasmic, is not able to achieve the same effect at elevated temperatures when the damage is solely to spindle
MTs. Since expression of theswi6-S160Aallele does not alter
transcription from SBF-regulated promoters (52), this suggests that there may be another role for Swi6p in MT stability that is distinct from its role in the SBF complex.
Deletion of CLB2 abrogates the Fcp1p-mediated
suppres-sion of the temperature and TBZ sensitivities ofcin8-3 kip1⌬
cells. To explore the possibility that regulators of the SBF
complex are also involved inFCP1’s suppression and therefore
relevant to our background, we screened different cyclins for
their ability to mitigate the effect ofFCP1overexpression on
MT stability in thecin8-3 kip1⌬background. The rationale for
looking into cyclins is that Cin8p, Kip1p, Fcp1p, Swi6p, and Swi4p were all previously shown to physically interact with Cdc28p, the catalytic subunit of the main cyclin-dependent kinase (1, 8, 22, 56), which pairs up with different cyclins to regulate cell cycle progression in yeast. Furthermore, strains
containing certain cyclin orCDC28deletions are incapable of
assembling a bipolar spindle, a phenotype shared with our double mutant (20).
As the periodic expression of cyclins serves to limit the window of action of Cdc28p to the proper time in the cell cycle
FIG. 4. Serial dilutions ofcin8-3 kip1⌬swi6⌬cells containing centromeric plasmids indicated on the left. Aliquots (5l) of cultures at a density of 2⫻106cells/ml, followed by three serial 10-fold dilutions, were spotted onto synthetic complete medium (SC) at the permissive temperature (26°C) or at the nonpermissive temperature (35°C) and on YPD medium containing different concentrations of TBZ. (A) Cells that also contained
FCP1on a 2m plasmid; (B) cells withoutFCP1overexpression.
on September 8, 2020 by guest
http://ec.asm.org/
(17), we generated triple mutants, strains with cin8-3 kip1⌬ mutations and mutations of different cyclins in order to test for a stage-specific dependency of the suppression (Fig. 5A). De-letions of different cyclins were introduced individually into cin8-3 kip1⌬cells carryingFCP1on a multicopy plasmid. As
before, we looked for a deletion that abrogatesFCP1’s
sup-pression of the temperature and TBZ sensitivities and for
complementation by a plasmid carrying wild-typeCIN8.CLB2
was the only gene whose deletion satisfied these parameters
(Fig. 5). Incidentally, CLB2 codes for a B-type cyclin that
activates Cdc28p to promote the transition from G2to M phase
(23), which corresponds to the timing ofcin8-3 kip1⌬ arrest
following G1synchronization and release at the restrictive
tem-perature (27, 47, 49). We also tested for the ability of the three
mutations to antagonize FCP1’s suppression (i.e., stabilize
MTs when deleted and whenFCP1overexpression is absent),
but none of the deletions above restored growth withoutFCP1
overexpression (not shown).
Surprisingly, deletion ofCLN3orWHI5, both of which are
known to regulate the SBF complex at the G1/S transition (15),
had no effect on the Fcp1p-mediated rescue of the double mutant phenotype. This corroborates earlier indications that Swi6p has a function in MT stability that is independent of the SBF complex.
DISCUSSION
This study suggests novel roles in MT stability for proteins that were not previously associated with this function. It opens new avenues of research into the exploration of the specific
mechanisms by which upstream cell cycle regulators affect the stability of the microtubule cytoskeleton.
FCP1overexpression and MT stability.FCP1is an essential
gene in both budding and fission yeast and is conserved among eukarya (2). Domain analysis of Fcp1p revealed that both the phosphatase activity and functional targeting to the transcrip-tion machinery are required for the suppression of the
tem-perature sensitivity ofcin8-3 kip1⌬cells (Fig. 1C). These data,
together with the established role of Fcp1p in global gene
transcription, suggest that the effect ofFCP1 overexpression
on MT stability may not be direct (i.e., mediated through physical interaction with components of the cytoskeleton) but instead may be indirect, possibly involving different pathways that act in conjunction or in parallel. Beyond the role of Fcp1p in transcription, it may be a key player in shaping specific phosphorylation patterns in its substrate, the C-terminal do-main of the large subunit of RNAPII, through dephosphory-lation of Ser-2 in the heptad repeats of the CTD. The signifi-cance of the phosphorylation state of the CTD of RNAPII is iterated in the proposed “CTD code” (5), which was suggested to convey information to CTD-binding proteins, some of which recognize particular phosphorylation patterns and trigger spe-cific cellular responses (19).
How can overexpression of FCP1 suppress defects
associ-ated with a compromised microtubule cytoskeleton?
Muta-tions inFCP1increase phosphorylation of Ser2 residues of the
CTD of RNAPII and rapidly shut down mRNA synthesis (30,
37). Conversely,FCP1overexpression inSchizosaccharomyces
pomberesults in increased levels of the hypophosphorylated (IIa) form of the CTD relative to control strains (35). By
overexpressing FCP1 in our strains, we may be mimicking
massive dephosphorylation of the CTD. It was demonstrated inXenopusextracts that upon fertilization, the CTD undergoes fast and massive dephosphorylation that is attributable to the Xenopus orthologue of FCP1 (45). This dephosphorylation contributes to the preparation of the transcriptional machinery for zygotic genome activation, as it is accompanied by a shift between different gene expression programs for the fertilized oocyte. The transition results in the activation of intracellular signals, some of which regulate changes in the microtubule cytoskeleton, including remodeling of the MT architecture that is required for expulsion of half the chromosomes into the polar body and for the establishment of a cytoplasmic micro-tubule network whose responsibility is to promote migration of
the male and female pronuclei (21). Likewise,FCP1
overex-pression in our genetic background may specify a transcrip-tional program that is accompanied by changes in MT stability.
SBF-dependent route to MT stability. The transcriptional
program specified byFCP1overexpression that is relevant for
MT stability converges with Swi6p, Swi4p, and Clb2p, because
deletion of these genes, separately, in thecin8-3 kip1⌬
back-ground abrogates theFCP1-mediated suppression of the
tem-perature sensitivity and the sensitivity to MT-destabilizing drugs that are associated with these cells.
The relevance of Clb2p for MT-dependent processes was
previously characterized. Cells in whichCLB2 has been
de-leted have a diminished capacity to separate the SPBs (20), a
phenotype in common withcin8-3 kip1⌬double mutants grown
at the nonpermissive temperature after release from␣
-factor-induced G1 arrest (27, 47, 49). Additionally, the cyclin/Cdk
FIG. 5. Deletion ofCLB2in thecin8-3 kip1⌬ background abro-gates the effect ofFCP1overexpression. (A) Aliquots (5l) of cultures at a density of 2⫻106cells/ml, followed by three serial 10-fold dilu-tions, were spotted onto synthetic complete (SC) medium at the per-missive (26°C) and nonperper-missive (35°C) temperatures. (B) Introduc-tion of a centromeric plasmid containing a wild-type copy of theCIN8
gene rescues the temperature and TBZ sensitivity associated with
cin8-3 kip1⌬clb2⌬[FCP1-2m] cells. Serial dilutions were performed as described in Materials and Methods.
on September 8, 2020 by guest
http://ec.asm.org/
complex Clb2/Cdc28 was shown to promote SPB separation and spindle assembly via Cin8p and Kip1p. The motors are
phosphorylated directly by the Clb2/Cdc28 complexin vitroand
in vivo, and this phosphorylation plays a role in promoting SPB separation and spindle assembly (8). In another study, it was proposed that Clb2/Cdc28 kinase activity regulates SPB sepa-ration indirectly by preventing the degradation of the motors (12).
Still, there may be paths to MT stability mediated through Clb2p that were not previously considered, paths that become dominant when the MT cytoskeleton is compromised. For ex-ample, the Clb2/Cdc28 complex, by virtue of its interaction with and phosphorylation of Swi4p, was shown to switch off
SBF-dependent transcription at the G2/M phase (1, 38), a time
whencin8-3 kip1⌬cells become arrested at the restrictive
tem-perature (27, 47). In our study, the presence of FCP1 on a
multicopy plasmid correlated with a proportional reduction in the levels of Swi6p and Swi4p relative to cells not
overexpress-ingFCP1 (Fig. 3), suggesting that the SBF complex is being
downregulated, and expression of theswi6-S160Dgene, which
leads to reduced activity from SBF promoters (52), was able to
substitute for FCP1 overexpression in suppressing the TBZ
sensitivity. Because of this similar effect of Clb2p at G2/M, of
FCP1 overexpression, and of constitutive expression of
swi6-S160Don the SBF complex and on MT-dependent processes, it is not unreasonable to propose that when MT stability is compromised, downregulation of the SBF complex alleviates the impairment. This downregulation, in turn, may lead to rerouting of the transcriptional program in favor of greater MT stability.
SBF-independent route to MT stability.Several lines of
ev-idence led us to propose the notion of an SBF-independent route to MT stability. The first is the suppression of the
tem-perature sensitivity ofcin8-3 kip1⌬swi6⌬cells by constitutive
expression of the S160A allele of SWI6 (Fig. 4B), an allele
which does not lead to major changes in cell cycle-specific activation or repression of transcription of SBF-regulated pro-moters (52). The second is the fact that deletion of regulators of the SBF complex did not achieve the same abrogation of
FCP1suppression, as did a deletion ofSWI4orSWI6(Fig. 5A).
Swi6p shuttles in and out of the nucleus in a cell cycle-dependent manner, with nuclear import occurring concomi-tantly with dephosphorylation of Ser-160 in late mitosis and
with nuclear export peaking from late G1 to M phase and
requiring phosphorylation of Ser-160 (22). Activation or re-pression of Swi6p-regulated genes is independent of Swi6p’s phosphorylation and hence, its localization (52). Aspartate substitution of serine 160 was shown to significantly impair nuclear localization, causing Swi6p to remain predominantly cytoplasmic throughout the cell cycle (though Swi6p is not completely excluded from the nucleus), whereas an alanine substitution at the same position leads to constitutive nuclear accumulation of Swi6p (52).
Interestingly, there is another convergence point between Clb2p and Swi6p which is independent of the SBF complex but dependent on the principal mitotic exit regulator Cdc14p. Clb2/Cdc28 was shown to lead to the liberation of Cdc14p from nucleolar sequestration in early anaphase (3). At anaphase onset, when the spindle starts to elongate, the phosphatase activity of Cdc14p was shown to be required for nuclear
mi-crotubule stabilization (25) and for the localization of spindle
midzone proteins like Cin8p (34). WhenCLB2is deleted, cells
are defective for early anaphase release of Cdc14p (3), and cdc14-1 clb2::LEU2double mutants do not start anaphase and arrest with unseparated spindle pole bodies (57), an arrest
phenotype shared withcin8-3 kip1⌬cells cultured at the
non-permissive temperature after release from G1synchronization
(20). Recently, it was shown thatCDC14overexpression can
bring about SPB separation in cin8-3 kip1⌬ cells (9). In an
unrelated study, Cdc14p was shown to be able to trigger
nu-clear import of Swi6pin vivoand to dephosphorylate Swi6p at
serine 160in vitro(22), a condition mimicked with the
swi6-S160A(nonphosphorylatable) nuclear allele we employed.
We therefore propose that incin8-3 kip1⌬cells, Swi6p may
have the potential to mediate the role of Cdc14p in nuclear MT stabilization through a similar mechanism to that by which Cdc14p leads to an increase in nuclear MT stability in wild-type cells (25). Cdc14p normally recruits Swi6p to the nucleus only in late M phase (22), but constitutive nuclear localization
of Swi6p (through expression of theS160Aallele) may be able
to stabilize spindle MTs in the compromised cin8-3 kip1⌬
background. This is supported by our finding that the nuclear version of Swi6p restores growth at the nonpermissive
temper-ature independently ofFCP1overexpression (Fig. 5B).
Our results demonstrating that certain SWI6alleles play a
part in MT stability raise the possibility that under normal conditions, transient expression of different phosphorylated forms and their restriction to specific cell cycle phases serves to limit the functional diversity of Swi6p. Constitutive expression of modified forms of Ser-160 may bring out Swi6p’s MT-sta-bilizing potential.
ACKNOWLEDGMENTS
This work was supported by grants from the National Institutes of Health (grant GM065885) and the Professional Staff Congress of the City University of New York.
We thank P. Lipke, S. Krishnamurthy, A. Kumar, L. Breeden, B. Andrews, and C. Forest for donating strains, plasmids, antibodies, primers, and other materials for this work, P. Lipke and R. Tal for critical comments on the manuscript, and R. Tal and E. Korolyev for technical assistance.
REFERENCES
1.Amon, A., M. Tyers, B. Futcher, and K. Nasmyth.1993. Mechanisms that help the yeast cell cycle clock tick: G2 cyclins transcriptionally activate G2 cyclins and repress G1 cyclins. Cell74:993–1007.
2.Archambault, J., et al.1997. An essential component of a C-terminal domain phosphatase that interacts with transcription factor IIF in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. U. S. A.94:14300–14305.
3.Azzam, R., et al.2004. Phosphorylation by cyclin B-Cdk underlies release of mitotic exit activator Cdc14 from the nucleolus. Science305:516–519. 4.Berlin, V., C. A. Styles, and G. R. Fink.1990. BIK1, a protein required for
microtubule function during mating and mitosis in Saccharomyces cerevisiae, colocalizes with tubulin. J. Cell Biol.111:2573–2586.
5.Buratowski, S.2003. The CTD code. Nat. Struct. Biol.10:679–680. 6.Cabanas, R., G. Castella, M. L. Abarca, M. R. Bragulat, and F. J. Cabanes.
2009. Thiabendazole resistance and mutations in the beta-tubulin gene of Penicillium expansum strains isolated from apples and pears with blue mold decay. FEMS Microbiol. Lett.297:189–195.
7.Carminati, J. L., and T. Stearns. 1997. Microtubules orient the mitotic spindle in yeast through dynein-dependent interactions with the cell cortex. J. Cell Biol.138:629–641.
8.Chee, M. K., and S. B. Haase.2010. B-cyclin/CDKs regulate mitotic spindle assembly by phosphorylating kinesins-5 in budding yeast. PLoS Genet. 6:e1000935.
9.Chiroli, E., G. Rancati, I. Catusi, G. Lucchini, and S. Piatti.2009. Cdc14 inhibition by the spindle assembly checkpoint prevents unscheduled centro-some separation in budding yeast. Mol. Biol. Cell20:2626–2637.
on September 8, 2020 by guest
http://ec.asm.org/
10.Costanzo, M., O. Schub, and B. Andrews.2003. G1 transcription factors are differentially regulated in Saccharomyces cerevisiae by the Swi6-binding pro-tein Stb1. Mol. Cell. Biol.23:5064–5077.
11.Cottingham, F. R., and M. A. Hoyt.1997. Mitotic spindle positioning in Saccharomyces cerevisiae is accomplished by antagonistically acting micro-tubule motor proteins. J. Cell Biol.138:1041–1053.
12.Crasta, K., H. H. Lim, T. H. Giddings, Jr., M. Winey, and U. Surana.2008. Inactivation of Cdh1 by synergistic action of Cdk1 and polo kinase is nec-essary for proper assembly of the mitotic spindle. Nat. Cell Biol.10:665–675. 13.Dahmus, M. E.1996. Reversible phosphorylation of the C-terminal domain
of RNA polymerase II. J. Biol. Chem.271:19009–19012.
14.Davidse, L. C., and W. Flach.1978. Interaction of thiabendazole with fungal tubulin. Biochim. Biophys. Acta543:82–90.
15.de Bruin, R. A., W. H. McDonald, T. I. Kalashnikova, J. Yates III, and C. Wittenberg.2004. Cln3 activates G1-specific transcription via phosphoryla-tion of the SBF bound repressor Whi5. Cell117:887–898.
16.Dubois, M. F., V. T. Nguyen, S. Bellier, and O. Bensaude.1994. Inhibitors of transcription such as 5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole and isoquinoline sulfonamide derivatives (H-8 and H-7) promote dephosphory-lation of the carboxyl-terminal domain of RNA polymerase II largest sub-unit. J. Biol. Chem.269:13331–13336.
17.Dynlacht, B. D., K. Moberg, J. A. Lees, E. Harlow, and L. Zhu.1997. Specific regulation of E2F family members by cyclin-dependent kinases. Mol. Cell. Biol.17:3867–3875.
18.Endow, S. A., et al.1994. Yeast Kar3 is a minus-end microtubule motor protein that destabilizes microtubules preferentially at the minus ends. EMBO J.13:2708–2713.
19.Fabrega, C., V. Shen, S. Shuman, and C. D. Lima.2003. Structure of an mRNA capping enzyme bound to the phosphorylated carboxy-terminal do-main of RNA polymerase II. Mol. Cell11:1549–1561.
20.Fitch, I., et al.1992. Characterization of four B-type cyclin genes of the budding yeast Saccharomyces cerevisiae. Mol. Biol. Cell3:805–818. 21.Gallicano, G. I.2001. Composition, regulation, and function of the
cytoskel-eton in mammalian eggs and embryos. Front. Biosci.6:D1089–D1108. 22.Geymonat, M., A. Spanos, G. P. Wells, S. J. Smerdon, and S. G. Sedgwick.
2004. Clb6/Cdc28 and Cdc14 regulate phosphorylation status and cellular localization of Swi6. Mol. Cell. Biol.24:2277–2285.
23.Ghiara, J. B., et al.1991. A cyclin B homolog in S. cerevisiae: chronic activation of the Cdc28 protein kinase by cyclin prevents exit from mitosis. Cell65:163–174.
24.Gonzalez-Ballester, D., A. de Montaigu, A. Galvan, and E. Fernandez.2005. Restriction enzyme site-directed amplification PCR: a tool to identify re-gions flanking a marker DNA. Anal. Biochem.340:330–335.
25.Higuchi, T., and F. Uhlmann.2005. Stabilization of microtubule dynamics at anaphase onset promotes chromosome segregation. Nature433:171–176. 26.Horak, C. E., et al.2002. Complex transcriptional circuitry at the G1/S
transition in Saccharomyces cerevisiae. Genes Dev.16:3017–3033. 27.Hoyt, M. A., L. He, K. K. Loo, and W. S. Saunders.1992. Two
Saccharomy-ces cerevisiae kinesin-related gene products required for mitotic spindle assembly. J. Cell Biol.118:109–120.
28.Huffaker, T. C., J. H. Thomas, and D. Botstein.1988. Diverse effects of beta-tubulin mutations on microtubule formation and function. J. Cell Biol. 106:1997–2010.
29.Huyett, A., J. Kahana, P. Silver, X. Zeng, and W. S. Saunders.1998. The Kar3p and Kip2p motors function antagonistically at the spindle poles to influence cytoplasmic microtubule numbers. J. Cell Sci.111(Part 3):295–301. 30.Jeong, S. J., et al.2005. Role of RNA polymerase II carboxy terminal domain
phosphorylation in DNA damage response. J. Microbiol.43:516–522. 31.Joshi, H. C.1998. Microtubule dynamics in living cells. Curr. Opin. Cell Biol.
10:35–44.
32.Kaiser, C. M., and S. A. Mitchell.1994. Methods in yeast genetics: a labo-ratory course manual. Cold Spring Harbor Labolabo-ratory Press, Cold Spring Harbor, NY.
33.Kamenski, T., S. Heilmeier, A. Meinhart, and P. Cramer.2004. Structure and mechanism of RNA polymerase II CTD phosphatases. Mol. Cell15: 399–407.
34.Khmelinskii, A., C. Lawrence, J. Roostalu, and E. Schiebel.2007. Cdc14-regulated midzone assembly controls anaphase B. J. Cell Biol.177:981–993. 35.Kimura, M., H. Suzuki, and A. Ishihama.2002. Formation of a
carboxy-terminal domain phosphatase (Fcp1)/TFIIF/RNA polymerase II (pol II) complex in Schizosaccharomyces pombe involves direct interaction between Fcp1 and the Rpb4 subunit of pol II. Mol. Cell. Biol.22:1577–1588. 36.Kobor, M. S., et al.1999. An unusual eukaryotic protein phosphatase
re-quired for transcription by RNA polymerase II and CTD dephosphorylation in S. cerevisiae. Mol. Cell4:55–62.
37.Kobor, M. S., et al.2000. A motif shared by TFIIF and TFIIB mediates their interaction with the RNA polymerase II carboxy-terminal domain phospha-tase Fcp1p in Saccharomyces cerevisiae. Mol. Cell. Biol.20:7438–7449. 38.Koch, C., A. Schleiffer, G. Ammerer, and K. Nasmyth.1996. Switching
transcription on and off during the yeast cell cycle: Cln/Cdc28 kinases acti-vate bound transcription factor SBF (Swi4/Swi6) at start, whereas Clb/Cdc28 kinases displace it from the promoter in G2. Genes Dev.10:129–141. 39.Korolyev, E., O. Steinberg-Neifach, and D. Eshel.2005. Mutations in the
yeast kinesin-like Cin8p are alleviated by osmotic support. FEMS Microbiol. Lett.244:379–383.
40.Krishnamurthy, S., X. He, M. Reyes-Reyes, C. Moore, and M. Hampsey. 2004. Ssu72 is an RNA polymerase II CTD phosphatase. Mol. Cell14:387– 394.
41.Kumar, A., et al.2004. Large-scale mutagenesis of the yeast genome using a Tn7-derived multipurpose transposon. Genome Res.14:1975–1986. 42.Lim, H. H., P. Y. Goh, and U. Surana.1996. Spindle pole body separation in
Saccharomyces cerevisiae requires dephosphorylation of the tyrosine 19 res-idue of Cdc28. Mol. Cell. Biol.16:6385–6397.
43.Nakaune, R., et al.1998. A novel ATP-binding cassette transporter involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Appl. Environ. Microbiol.64:3983–3988.
44.Nakaune, R., H. Hamamoto, J. Imada, K. Akutsu, and T. Hibi.2002. A novel ABC transporter gene, PMR5, is involved in multidrug resistance in the phytopathogenic fungus Penicillium digitatum. Mol. Genet. Genomics267: 179–185.
45.Palancade, B., M. F. Dubois, M. E. Dahmus, and O. Bensaude.2001. Tran-scription-independent RNA polymerase II dephosphorylation by the FCP1 carboxy-terminal domain phosphatase in Xenopus laevis early embryos. Mol. Cell. Biol.21:6359–6368.
46.Peterson, J. B., and H. Ris.1976. Electron-microscopic study of the spindle and chromosome movement in the yeast Saccharomyces cerevisiae. J. Cell Sci.22:219–242.
47.Roof, D. M., P. B. Meluh, and M. D. Rose.1992. Kinesin-related proteins required for assembly of the mitotic spindle. J. Cell Biol.118:95–108. 48.Rutherford, S. L.2000. From genotype to phenotype: buffering mechanisms
and the storage of genetic information. Bioessays22:1095–1105. 49.Saunders, W. S., D. Koshland, D. Eshel, I. R. Gibbons, and M. A. Hoyt.1995.
Saccharomyces cerevisiae kinesin- and dynein-related proteins required for anaphase chromosome segregation. J. Cell Biol.128:617–624.
50.Segal, M., D. J. Clarke, and S. I. Reed.1998. Clb5-associated kinase activity is required early in the spindle pathway for correct preanaphase nuclear positioning in Saccharomyces cerevisiae. J. Cell Biol.143:135–145. 51.Shaw, S. L., E. Yeh, P. Maddox, E. D. Salmon, and K. Bloom.1997. Astral
microtubule dynamics in yeast: a microtubule-based searching mechanism for spindle orientation and nuclear migration into the bud. J. Cell Biol. 139:985–994.
52.Sidorova, J. M., G. E. Mikesell, and L. L. Breeden.1995. Cell cycle-regulated phosphorylation of Swi6 controls its nuclear localization. Mol. Biol. Cell 6:1641–1658.
53.Solomon, F., C. L. D. Kirkpatrick, V. Praitis, and B. Weinstein.1992. Methods for studying the yeast cytoskeleton, p. 197–222.InK. Carraway and C. Carraway (ed.), The cytoskeleton. Oxford University Press, Oxford, United Kingdom.
54.Steinberg-Neifach, O., and D. Eshel.2000. Simultaneous expression of both MAT loci in haploid cells suppresses mutations in yeast microtubule motor genes. Mol. Gen. Genet.264:300–305.
55.Walker, G. M.1982. Cell cycle specificity of certain antimicrotubular drugs in Schizosaccharomyces pombe. J. Gen. Microbiol.128:61–71.
56.Wong, J., et al.2007. A protein interaction map of the mitotic spindle. Mol. Biol. Cell18:3800–3809.
57.Yuste-Rojas, M., and F. R. Cross.2000. Mutations in CDC14 result in high sensitivity to cyclin gene dosage in Saccharomyces cerevisiae. Mol. Gen. Genet.263:60–72.