Cell signalling and gene regulation in early
Xenopus
development
Katherine Alison Neal
October 1998
A thesis submitted to the University of London for the degree of Doctor of Philosophy
Division of Developmental Biology, National Institute for Medical Research,
ProQuest Number: 10014725
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10014725
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author. All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
A cknow ledgem ents
I am grateful to the Director of the Institute, Dr J. J. Skehel, and the Director
of Studies, Dr R. W. King for giving me the opportunity to do m y PhD at the
NIMR. Thanks to the Medical Research Council w ho provided the fu n d in g
for m y PhD.
A big thank you to all the members of the D ivision of D ev elo p m en ta l
Biology, past and present, for their advice, discussion and friendship. They
have made m y time at the Institute stim ulating and also great fun. Of all of
them, I ow e particular thanks to Branko Latinkic, w hom I worked closely
w ith during the first part of my PhD, and N iall Armes w hose en th u sia sm
and encouragem ent I w ill never forget.
I am grateful to all my friends and colleagues around the Institute for all the
good times that w e have shared.
I w ould like to thank m y parents and my sister Hilary for all their love and
support, I feel so lucky to have them as m y family. I am grateful to A liso n
Pryce for her friendship and for all the memorable days that w e spent in the
lovely Archway. A special thank you goes to Julian Mackintosh, w ith o u t
w hom I w onder if this thesis w ould ever have got into print!
Finally I w ould like to thank m y supervisor Dr Jim Smith. His w isd om ,
guidance and support have played a crucial role in m y training as a scientist.
Abstract
A bstract
The two activin type I receptors ALK-2 and ALK-4 have distinct effects on gene
expression in the Xenopus animal cap assay. Constitutively active ALK-4 (ALK-4*)
can reproduce the effects of activin treatment including the dose dependent induction
of progressively more dorso-anterior markers, whilst constitutively active ALK-2
(ALK-2*) induces only ventral mesodermal markers and counteracts the dorsalising
effects of ALK-4. The dorsal inducing properties of ALK-4* can be transferred to
ALK-2* by transfer of a small peptide loop (the p4-p5 loop) from the kinase domain of
one receptor to the other. In contrast, I show, that the equivalent region of the ALK-2*
receptor, when transferred to ALK-4*, cannot effectively counteract the dorsalising
effects of ALK-4*, suggesting that other regions of type I receptors are also involved in
determining signal specificity.
The chimeric receptor created by transfer of the ALK-2* GS domain and its flanking
sequences into ALK-4* (C9), and the chimeric receptor created by transfer of the
activation loop of the ALK-2* kinase (Cll) into ALK-4* cannot induce the full range of
ALK-4* responses, although they contain the ALK-4* (34-^5 loop. My results suggest
that ALK-4* regulates at least two different signalling events, one of which is specified
by C9 and C ll.
The Xenopus Brachyury (Xbra) gene is induced in response to the mesoderm inducing factors FGF and activin (and by the activin receptors ALK-2* and ALK-4*). I have
examined the transcriptional regulation of a pseudoallele of Xbra, Xbra2, in response to
FGF. My results indicate that 381 bp 5' of the Xbral transcription start site are
sufficient to confer responsiveness to FGF (and activin).
Examination of the Xbral 5' flanking sequence revealed a serum response-like element.
This suggested that activation of Xbral in response to FGF may occur in a similar way
to the activation of c-fos, through the activation of a ternary complex bound to an
SRE-like element. My results show that this is unSRE-likely. However, I provide preliminary
evidence that a serum response-like factor functions as a negative regulator of
Table of contents
A C K N O W L E D G E M E N T S ... 3
A B S T R A C T ...4
CHAPTER 1... 1 3
In t r o d u c t i o n... 1 3
Early Xenopus d ev elo p m en t... 14 M esoderm induction and D orsalisation... 14
E stablishm ent of the dorso-ventral a x is ...18
M esoderm inducing fa c to r s... 2 5
FGF family m e m b e r s...2 5
TGF-p superfamily m em b ers...2 8
A ctiv in ...2 9
C o n cen tration -d ep en d en t effects of activin... 3 0
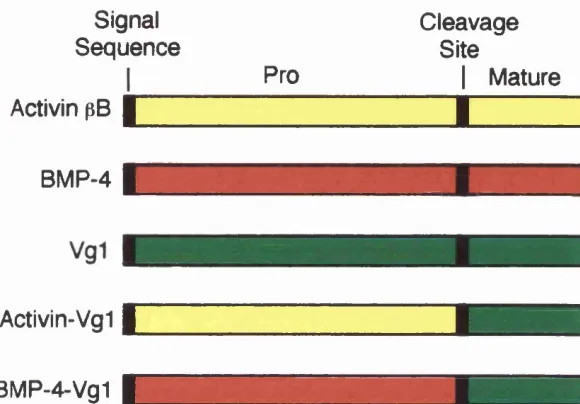
V g -1 ... 3 4
Xenopus nodal-related 1 - 4 ...3 6 B on e M orphogenetic Proteins (B M P s)... 3 6
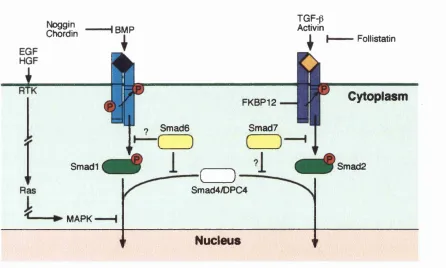
TGF-p signalling in early Xenopus d evelop m en t... 3 8 Sm ad s: m ediators of TGF-p family signalling... 4 3
P athw ay-restricted S m a d s ...4 5
C om m on-m ediator S m a d s ... 4 8
S m a d protein dom ains and their function... 5 0
Transcriptional regulation by S m a d s ... 51
Inhibitory S m a d s ... 5 3
Inhibition of S m a d s by MAP kinase p a th w a y s...5 4
R eceptor-interacting p ro tein s...55
Other m ediators of TGF-p signalling...5 6
X enopus Brachyury....5 6 Activation of X b ra e x p r e ss io n ...58
Table of contents
Transcriptional regulation of X b ra... 62 This s t u d y ... 6 4
CHAPTER 2... 6 5
Ma t e r ia l sa n d Me t h o d s...6 5 EMBRYOS AND EMBRYO MANIPULATIONS... 6 5
Obtaining Xenopus e m b r y o s ... 6 5 Microinjection of Xenopus e m b r y o s ...6 5 Animal ca p d is s e c tio n ...6 6
W hole mount antibody sta in in g ...6 6
MOLECULAR BIOLOGY TEC H N IQ UES... 6 7
DNA quantification and m anipulation... 6 7
Preparation and sto ra g e of co m p eten t bacteria...6 8
P lasm id transformation of com p eten t b a c te r ia ... 68
Sm all s c a le preparation of plasmid D N A ... 69
Large s c a le preparation of plasmid DNA...70
P o ly m er a se chain reaction...7 0
A g a ro se gel electrophoresis of DNA and R N A ...7 0
Purification of sp ecific DNA fragm ents from g e l s ...70
DNA s e q u e n c e a n a ly s is ... 71
In vitro tran scrip tion ...71 PLASMID CONSTR UC TS...7 3
L u cifera se-b a sed v e c to r s... 7 3
E F -1 a b a se d vector... 7 4
ALK-4* point mutant constructs; ALK-4*S^^^—>A and ALK-4*T’^®—>V...7 5
RNA ISOLATION AND ANALYSIS... 75
RNA iso la tio n ... 7 5
R N A ase protection ...76
LUCIFERASE AND p-GALACTOSIDASE A S S A Y S ... 78
Table of contents
FORMULATION OF FREQUENTLY USED BACTERIAL GROWTH MEDIA... 80
CHAPTER 3... 8 1
I n v e s t i g a t i o n o f t h e ALK-2* d o m i n a n t v e n t r a l i s a t i o n s i g n a l ...8 1
INTRODUCTION... 81
R ESU LTS... 87
The ability of ALK-2* to interfere with ALK-4*-mediated g e n e induction can n ot be recapitulated by transfer of the 181 region of ALK-2* into the ALK-4* b a c k b o n e ... 87
T he ability of ALK-2* to interfere with dorsal g e n e induction requires the ALK-4 receptor b ack b on e 8 9
O v er-exp ression of S m a d i or Sm adS d o e s not interfere with induction of dorsal g e n e s by ALK-4*. 92
ALK-2* d o e s not interfere with Sm ad 2/S m ad 4-m ed iated induction of dorsal g e n e s in Xenopus animal c a p s ... 92
D ISC U SSIO N ... 9 5
T he P4-P5 loop (181) d o e s not define all signalling properties of type I r e c e p to r s... 9 9
ALK-2* blocks dorsal g e n e induction upstream of S m ad sig n a llin g ... 100
Susceptibility to inhibition by ALK-2* requires s e q u e n c e s p resen t in ALK-4 that are not within the (34-(35 loop...100
T ype 1 receptor S m ad -in d ep en d en t signalling p a th w a y s... 100
CHAPTER 4... 1 0 2
ALK-4* SPECIFIES AT LEAST TWO DIFFERENT DOWNSTREAM SIGNALS... 1 0 2
INTRODUCTION... 102
R ESU LTS...104
T he signalling sp ecificities of the ALK-4* backbone chim eras, C 9 and C l 1, are qualitatively different to that of ALK-4*... 104
C o -ex p ressio n of C 9 and C l 1 d o e s not restore g o o s e c o /d induction... 109
Mutation of juxtam em brane serin e 172 and threonine 176 d o e s not affect ALK-4* signalling capacity. 110
A Sm ad4 blocks ALK-2*, C 9 and C 1 1 re sp o n se s, but not ALK-4*, C2 and C 4 r e s p o n s e s ... 111
C o-exp ression of S m ad 2 and Sm ad4, but not C9 or C l 1 and S m a d 4 , m im ics ALK-4* r e s p o n s e s .. .1 2 0
C 9 in d u ces se c o n d a r y a x e s ...120
Table of contents
ALK-4* s p e c ifie s at least two different dow nstream s i g n a ls ...123
C o -ex p ressio n of 0 9 and 0 1 1 d o e s not restore g o o s e c o /d activation...1 2 4 A Sm ad4 blocks ALK-2*, 0 9 and 0 1 1 re sp o n se s, but not ALK-4*, 0 2 and 0 4 r e s p o n s e s ... 124
O o-exp ression of Sm ad 2 and Sm ad4, but not 0 9 or 0 1 1 and Sm ad 4, m im ics ALK-4* r e s p o n s e s 126 Model: ALK-4* sp e c ifie s at least two different dow nstream sig n a ls ... 128
0 9 in d u ces seco n d a ry a x e s ...130
CHAPTER 5... 1 3 2 I d e n t i f i c a t i o n o f s e q u e n c e s w ith in -2.1 kb o f t h e Xb r a2 t r a n s c r i p t i o n s t a r t s i t e t h a t RESPOND TO BOTH F G F AND ACTIVIN... 1 3 2 INTRODUCTION...132
R ESU LTS... 133
R em oval of vector s e q u e n c e s from Xbra2pG L2 reporter co n str u c ts... 133
Norm alising luciferase activity... 137
Tim e co u r se an alysis of -866Xbra2pGL2 activation in r e sp o n se to FGF... 140
S e q u e n c e within 381 bp of the transcriptional start site is sufficient to elicit a r e sp o n s e to FGF (and a c tiv in )...140
Xenopus Sm ad2 a ctiv a tes a luciferase reporter construct containing 381 bp of X b ra2 5 ' flanking s e q u e n c e ... 14 3 D ISC U SSIO N ...145
S e q u e n c e s within -381 bp of the Xdra2transcription start site respond to both FGF and activin ...1 4 5 S e q u e n c e s within -381 bp of the Xbra2 transcriptional start site respond to S m a d 2 ... 146
CHAPTER 6... 1 4 8 A TERNARY COMPLEX FACTOR LIKE ELK-1 IS UNLIKELY TO BE INVOLVED IN THE TRANSCRIPTIONAL ACTIVATION OF X B R A 2... 1 4 8 INTRODUCTION...148
R ESU LTS... 153
A dominant negative ternary com plex factor red u ces FGF and activin induction of e n d o g e n o u s X b r a 2...15 4 An activated ternary com p lex factor red u ces FGF and activin induction of X b ra 2...158
Table of contents
S e q u e n c e within 8 6 6 bp, but not 381 bp, of the transcriptional start site of X b ra 2 is sufficient to elicit
a resp o n se to Elk-1 V P16 and Elk-1 AB V P 1 6 ...164
An activated form of S R F blocks the activation of two luciferase reporter constructs, containing 8 6 6 bp or 381 bp of X b ra2 5 ' flanking s e q u e n c e , by FGF and activin... 166
An SRF-engrailed fusion protein c a u s e s animal cap ex ten sio n and a c tiv a te s a lu ciferase reporter construct containing 381 bp of X b ra2 5 ' flanking s e q u e n c e ...167
D ISC U SSIO N ...171
Elk-1 A 307 and Elk-1 V P16, but not Elk-1 AB A 307 and Elk-1 AB V P16, reduce th e level of X b ra 2 induced by FGF or a ctiv in ... 171
X b ra 2 promoter s e q u e n c e s b etw een -8 6 6 and -381 bp of th e transcriptional start site respond to Elk-1 V PElk-16 and Elk-Elk-1 AB V P Elk-16... Elk-172
A role for an SRF-like factor a s a negative regulator of m esoderm formation in Xenopus em b ryos. 173 R E F E R E N C E S ... 1 7 6 FIGURES AND TABLES CHAPTER 1 Figure 1.1 The three signal m o d e l... 17
Figure 1.2 C ross sectio n through a wild type and UV ventralised Xenopus em bryo... 19
Figure 1.3 C om parison of Drosophila and Xenopus Wnt signal transduction p ath w ays... 2 3 Figure 1.4 T he FGF signalling pathw ay...2 8 Figure 1.5 M odels for long-range signalling by a ctiv in ... 3 2 Figure 1.6 Chimeric Vg 1 co n stru cts... 3 5 Figure 1.7 Model for activin/TGF-p receptor a ctivation ...3 9 Figure 1.8 ALK-2* cou n teracts th e dorsalising effects of ALK-2*...4 3 Figure 1.9 Model show ing positive and negative regulation of TG F-p/Sm ad sig n a llin g ...4 5 Figure 1.10 Different functional dom ains in S m a d s ... 51
Figure 1.11 W hole mount in situ hybridisation show ing ex p ressio n of Xenopus Brachyury.... 57
Table 1.1 M olecules that re sc u e th e UV p h en o ty p e...21
Table of contents
CHAPTER 2
T able 2.1 C onstructs u sed a s tem p la tes for in vitro transcription... 72
T able 2 .2 C onstructs u se d a s tem p la tes for R N A ase protection p r o b e s ...77
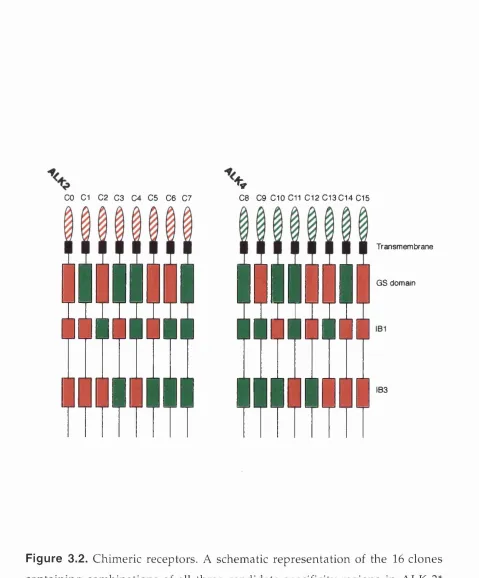
CHAPTER 3 Figure 3.1 Structure of type I receptor k inase and alignm ent of three r e g io n s s e le c te d for a n a ly s is ... 8 2 /8 3 Figure 3 .2 ALK-2VALK-4* Chimeric rec e p to r s...85
Figure 3 .3 Chimeric receptors with the ALK-2* IB1 do not interfere withi dorsal g e n e induction by ALK-4* or activin... 88
Figure 3 .4 ALK-4* s e q u e n c e s confer susceptibility to inhibition of d o rsa l g e n e activation by ALK-2*... 90/91 Figure 3 .5 S m a d i d o e s not interfere with dorsal g e n e activation by ALIK-2* or activin... 9 3 /9 4 Figure 3 .6 ALK-2* d o e s not interfere with dorsal g e n e activation in anim al c a p s ex p ressin g S m ad2 and S m a d 4 ...9 6 /9 7 Figure 3 .7 ALK-2* cou n teracts th e dorsalising effects of ALK-4* u p stream of S m ad sig n a llin g ... 98
CHAPTER 4 Figure 4.1 ALK-2*/ALK-4* Chimeric rec e p to r s...- ... 103
Figure 4 .2 C om parison of th e concen tration -d ep en d en t induction of X b ra, Pintallavis and goosecoid in r e sp o n s e to ALK-4* and C 9... 105
Figure 4 .3 C om parison of th e con cen tration -d ep en d en t induction of Xbra, Pintailavis and goosecoid in r e sp o n s e to ALK-4* and C l 1 ... 107
Figure 4 .4 C l 1 in d u ces five tim es m ore Xbra than is ev er s e e n in r e s p o n s e to ALK-4*... 108
Figure 4 .5 C o -ex p ressio n of C 9 and C l 1 d o e s not restore goosecoid e x p r e s s io n ... 110
Figure 4 .6 S e q u e n c e alignm ent of cytoplasm ic juxtam em brane region of typ e I recep to rs...112
Figure 4 .7 Mutation of juxtam em brane serin e1 7 2 and threonine 1 7 6 d o e s not affect ALK-4* signalling c a p a c it y ... 113
Figure 4 .8 A Sm ad4 d o e s blocks ALK-2*, C9 and C l 1 r e sp o n s e s, but not ALK-4* r e s p o n s e s ... 115
Figure 4 .9 ALK-2*/ALK-4* Chimeric r ecep to rs... 116
Figure 4 .1 0 A Sm ad4 d o e s not block ALK-4*, C2 and C 4 r e s p o n s e s ... 117
Figure 4.11 C o-exp ression of S m a d 2 with S m ad4, but not C9 or C l 1 with S m ad 4, mimics ALK-4* r e s p o n s e s ...119
Table of contents
Figure 4 .1 3 Mode! to explain why co-exp ression of Sm ad 2 and S m ad4, but not C9 or C l l ,
m im ics ALK-4* r e s p o n s e s ... 127
Figure 4 .1 4 (A) Model show ing ALK-4* sp ecifies two dow nstream sign als. (B) Model
sh ow in g ALK-4* sp e c ifie s S m ad -4 d ep en d en t and S m ad 4-in d ep en d en t s i g n a ls ...129
T able 4.1 A Sm ad4 d o e s blocks m esoderm induction in resp o n se to ALK-2*, 0 9 and 0 1 1 ,
but not ALK-4* 0 2 and 0 4 ... 118
T able 4 .2 ALK-4* and 0 9 can induce secon d ary a x e s ... 122
CHAPTER 5
Figure 5.1 A 2 0 0 nucleotide protected fragment, recogn ised by the X b ra2 R N A ase protection p r o b e ....1 3 4 Figure 5 .2 S ch em atic representation of the region of X b ra2 u se d to m ake th e X b ra 2 R N A ase
protection p r o b e ... 136
Figure 5 .3 Normalistaion of luciferase activity in X b ra2 5' flanking s e q u e n c e rep o rtera ssa y s... 138 Figure 5 .4 Tim e co u rse of luciferase activity induced by FGF in animal ca p extracts from
em b ryos injected with -8 6 6 X b ra 2 p G L 2... 139 Figure 5 .5 X b ra 2 luciferase reporter constructs containing 381 bp of X b ra 2 5' flanking s e q u e n c e
respond to FGF... 141
Figure 5 .6 S e q u e n c e s upstream of -381 in the Xb ra2 5' flanking s e q u e n c e are d isp en sa b le for
r e sp o n s e of X b ra2 to FGF and activin...142 Figure 5 .7 X b ra 2 luciferase reporter constructs containing 381 bp of X b ra 2 5' flanking s e q u e n c e
respond to S m a d 2 ... 14 4
CHAPTER 6
Figure 6.1 1.2 kb of the X b ra2 5' flanking s e q u e n c e ... 149 Figure 6 .2 (A) Primary structure of SR F and the TCFs. (B) T he Grappling Hook m odel for ternary
co m p lex form ation... 151
Figure 6 .3 (A) S ch em a tic representation of activation of c-fos via activation of a ternary
com p lex. (B) Model for activation of X b ra2 via activation of a ternary c o m p le x ...153 Figure 6 .4 Primary structure of Elk-1 and four derivatives...155
Figure 6 .5 Elk-1 a 3 0 7 , but not Elk-1 a B a 3 0 7 , red u ces the level of induced in r esp o n se
to FGF and activin... 156
Figure 6 .6 Elk-1 V P16, but not Elk-1 a B V P16, red u ces th e level of X b ra2 induced in resp o n se to
FGF and a ctiv in ...159
Figure 6 .7 Primary structure of S R F and a constitutively active derivate... 161
Table of contents
Figure 6 .9 S R F-V P 16 inhibits elongation of animal c a p s cultured with FGF and activin... 163
Figure 6 .1 0 Elk-1 V P16 and Elk-1 aB VPI 6 activate ex p ressio n of -866X bra2pG L2, but do
not activate -3 81X b ra2p G L 2... 165
Figure 6.11 SR F -V P 16 blocks activation of -866Xib/a2pGL2 and -381Xbra2pGl_2 in r e sp o n s e to
FGF and a ctiv in ...167
Figure 6 .1 2 SRF-En'’ c a u s e s animal c a p s to extend and form protrusions... 169
Figure 6 .1 3 SRF-En'’ activates exp ression of -381X b/a2pG L 2...170
T able 6.1 Sum m ary of effects of the Elk-1 and SR F derivatives on the ex p ressio n of e n d o g e n o u s
____________________________________ Chapter 1____________________________________
Chapter 1
Introduction
Formation and patterning of the m esoderm are fundam ental to the
establishm ent of the vertebrate body plan. For this reason, the way in w h ich
m esoderm is induced and patterned is an intense area of study. Work o n
embryos of the amphibian, Xenopus l a e v i s , have provided researchers w ith
the m ost insight into the cellular and molecular interactions that occur
during these two processes.
The X e n o p u s embryo is an excellent system in w hich to study the m echanism s controlling m esoderm formation and patterning, for several
reasons. X e n o p u s embryos can be obtained in large numbers and they
develop outside the mother, making them accessible to experim ental
m anipulation at all developm ental stages. Furthermore, the embryos are
large in size which makes them relatively easy to microinject and dissect
into pieces that can be assayed for their inductive potential. In addition to
this an efficient procedure for generating transgenic X e n o p u s embryos has
recently been developed. Thus, transgenesis can be used to ectopically
express genes of interest, to direct expression of m odified gene products
w hich interfere with the function of their endogenous, w ild type
counterparts, and to analyse the spatial expression of promoters in the
embryo (Kroll and Amaya, 1996).
The first part of this thesis concerns the signalling specificities of ALK-2 and
ALK-4 w hich are receptors for the m esoderm inducing factor activin.
C onstitutively active forms of these receptors transduce clearly distinct
signals in X en op us (Armes and Smith, 1997). The second part of the study is
focused on the transcriptional regulation of X e n o p u s B r a ch y u r y , a target
gene for the m esoderm inducing factors FGF and activin and the activin
____________________________________ Chapter 1____________________________________
Early
X e n o p u s
d e v e l o p m e n tDuring its developm ent in the mother, the X e n o p u s oocyte becom es
polarised along its anim al-vegetal axis, but remains radially sym m etrical
around this axis. The animal-vegetal polarisation is clearly visible by a
difference in pigm entation, the animal hem isphere is dark and the vegetal
hem isphere is light. Oocytes contain stores of potential regulatory
m olecules, proteins and RNAs that serve to control em bryonic
developm ent prior to the onset of zygotic transcription, w hich begins at the
m id-blastula transition (MET) (Kirschner et al., 1985), w hen the em bryo
contains -4000 cells. Several RNAs are know n that are specifically localised
to a thin layer of cytoplasm (the cortex) underlying the plasma membrane in
the vegetal hem isphere, including the TGF-p family member Vg-1, w h ich
may be involved in m esoderm induction (Weeks and M elton, 1987), as w ill
be discussed later.
The radial symmetry of the oocyte is broken at fertilisation as a result of
sperm entry. This triggers a cytoplasmic shift, about half w ay through the
first cell cycle, w hich involves m ovem ent of the subcortical cytoplasm
relative to the plasma membrane and the cortex (Vincent and Gerhart, 1986;
Gerhart et al., 1989). The ventral side of the embryo, where the sperm enters
the oocyte, forms where the vegetal cytoplasm m eets the animal cortex,
giving rise to ventral structures like blood and m esenchym e. The dorsal
side, w hich gives rise to axial structures, such as notochord, som ites and
neural tissue, forms where the animal cytoplasm m eets the vegetal cortex.
M e s o d e r m Indu cti on a n d D o r s al is at io n
At early cleavage stages Xenopus embryos can be considered to consist of tw o
cell types: prospective ectoderm in the animal hem isphere and prospective
endoderm in the vegetal hem isphere. M esoderm arises during blastula
stages as a result of inductive interactions in w hich cells from the vegetal
half of the embryo act on cells in the equatorial region (marginal zone) of
the embryo causing them to form m esoderm rather than ectoderm .
____________________________________ Chapter 1____________________________________
expiant (animal cap) from a blastula stage embryo in contact w ith vegetal
pole cells. Cultured alone, the animal cap forms atypical epidermis; in the
presence of vegetal pole cells, it forms a variety of m esoderm al cell types
(N ieuw koop, 1969).
Vegetal pole cells induce two types of mesoderm. Those from the dorsal side
of the embryo tend to induce dorsal cell types, such as notochord and
som ites, w hile lateral and ventral vegetal blastomeres induce ven tral
structures such as blood and m esenchym e (Boterenbrood and N ieu w k o o p ,
1973; Dale et al., 1985). This is consistent w ith the finding that, at the 32-cell
stage, only dorsal-vegetal cells are able to induce dorsal structures (Dale and
Slack, 1987a) and to cause axial duplication w hen transplanted to the ven tral
side of a host 32-cell stage embryo (Gimlich, 1986). It seem s likely that this
activity represents a dorsal mesoderm inducing signal.
These data provided som e insight into how m esoderm is formed and
patterned in Xenopus. H owever, they appeared to contradict the fate map of
X e n o p u s embryos w hich show s that about 60% of somitic m uscle com es from the ventral half of the embryo (Dale and Slack, 1987a). In experim ents
in w hich blastulae were divided in half and cultured separately, ven tral
halves typically formed extreme ventral structures containing little or n o
m uscle (Dale and Slack, 1987b), suggesting that a signal from the dorsal half
of the embryo is required to produce the correct range of m esoderm al tissues
along the dorso-ventral axis. This process has been called 'dorsalisation'
(Smith and Slack, 1983; Dale and Slack, 1987b). Evidence for the existence of a
dorsal signal comes from experiments in w hich dorsal and ventral regions
from early gastrulae are juxtaposed. Isolated dorsal marginal zone tissue
forms notochord, w ith some muscle and neural tissue, w hile ventral
m arginal zone cells form blood, m esenchym e and m ésothélium . But, if the
tw o pieces are combined, although the dorsal tissue continues to
differentiate as notochord, the ventral marginal zone forms large am ou n ts
of m uscle (Slack and Forman, 1980; Dale and Slack, 1987b).
The source of the dorsalising signal in prospective m esoderm is the
____________________________________ Chapter 1____________________________________
organiser of an early gastrula stage embryo to the ventral side of another
gastrula, it was discovered that the organiser is capable of respecifying the
fate of host cells in the area, resulting in axial duplication (Spemann and
M angold, 1924; Smith and Slack, 1983). Due to the similarity of the axial
duplications caused by transplant of either dorsal vegetal cells from a 32-cell
stage embryo or the organiser region of an early gastrula stage embryo, it was
reasoned that the dorsal organiser activity may be induced to form as a result
of inductive signals emanating from underlying vegetal cells.
The processes of m esoderm induction and dorsalisation have been resolved
into the 'three signal model' (Fig. 1.1) (Smith and Slack, 1983). According to
this m odel, a signal from the ventral and lateral vegetal cells induces
ventral m esoderm , w hich comprises about 300° of the marginal zone, and
the dorso vegetal cells emit a signal that induces the formation of dorsal
m esoderm , or organiser, which makes up the remaining 60° of the m arginal
zone. Originally these two signals were considered to occur early in
developm ent, before the onset of zygotic transcription at the MET, w ith a
third signal, acting later, passing from the organiser region to expand the
size of the area of dorsal m esoderm (Fig. 1.1). Recently, how ever, there has
been evidence to suggest that the initial inductive signals do not pass from
the vegetal hem isphere to the animal hem isphere until after the MET
(Wylie et al., 1996). W ylie et al. have show n that w hen recombinants of
animal and vegetal explants are cultured together for a short, one h ou r
culture period, vegetal explants from late blastulae are able to induce
com petent animal caps to express M y o D . Vegetal explants from early
(preMET) blastulae, however, do not induce identical animal caps to express
M y o D , over the same period of time (Wylie et al., 1996). This dem onstrates that the cell interactions involving signals from the vegetal portion of the
embryo that are required to induce M y o D occur only after zygotic
transcription has started. It remains to be tested whether the same is true for
other markers.
Since the three signal m odel was devised, intensive effort has been made to
Chapter 1
Corticocytoplasinic rotation
Animal
.J
VegetalVentral Dorsal
Dorsalisation Mesoderm induction
anim al pole Marginal z o n e VM
o>
vegetal pot Localisation of
dorsal determ inants Induction of Ventral
an d Lateral M esoderm
anim al pole
Marginal z o n e VM VLM DLM DM vegetal pole
Figure 1.1. The three signal model (modified from Smith, 1989).
Embryological experiments described in the text predict the m odel show n.
As a result of oogenesis and cortical rotation, three territories are established:
the animal hemisphere, the ventro-vegetal blastomeres and the
dorso-vegetal blastomeres. The dorso-vegetal blastomeres induce mesoderm of a
corresponding character in the overlying animal cells. The dorsal
____________________________________ Chapter 1____________________________________
patterning. With the discoveries that have been made, it has been necessary
to make modifications to the model. Notably, the three signal m odel regards
ventral m esoderm as a 'ground state' that is adopted by lateral and ventral
regions in the absence of dorsalising signals. H ow ever, it appears that
ventral m esoderm formation requires active signalling and is not sim ply a
passive process (Sive, 1993), as w ill be discussed later.
E s t a b l i s h m e n t of t h e d o r so -v e nt r a l ax is
As m entioned above, the dorso-ventral axis is established at fertilisation as a
result of sperm entry w hich triggers cortical rotation. Ultraviolet (UV)
irradiation is effective at blocking the formation of the dorso-ventral axis
w hen applied at two different times: firstly w hen applied to the vegetal p o le
of mature ovarian oocytes, and secondly w hen applied to fertilised eggs
shortly after sperm entry, thereby preventing cortical rotation (H olw ill et al.,
1987; Elinson and Pasceri, 1989). Both treatments result in radially
symm etrical embryos that, at post gastrula stage, consist of three concentric
germ layers, the ectoderm, mesoderm and endoderm , that are of ven tral
character (Fig 1.2). The natures of the pre-fertilisation and post-fertilisation
dorsalising activities are not understood, although they are clearly separable;
normal developm ent can be restored in embryos that have been subjected to
UV irradiation just after fertilisation by tipping them so that cortical rotation
is re-imposed (Scharf and Gerhart, 1980), w hile eggs that are UV irradiated as
oocytes undergo normal cortical rotation and cannot be rescued by tipping
(Elinson and Pasceri, 1989). As UV irradiation of the vegetal pole of m ature
oocytes prevents the formation of a dorso-ventral axis, it seem ed possible
that a com ponent(s) of the dorsal pathway, or a factor that influences this
pathway, is established during oogenesis and localised to the cortex of the
vegetal pole before oocyte maturation. The vegetal localisation of an oocyte
dorsalising activity has been confirmed by cytoplasmic transfer experim ents
(H olowacz and Elinson, 1995), how ever the m olecular character of this
activity is not known.
Several lines of evidence indicate that, in the post-fertilisation U V -sen sitiv e
Chapter 1
Epidermis
Ventral Mesoderm
Endoderm
B
Neural tube
Somite
Notochord
Epidermis
Endoderm
LPM
Figure 1.2. Two events pattern the early embryo (taken from Heasm an,
1997). UV irradiation experiments reveal that at least two events pattern the
embryo: (A) a UV-insensitive event establishes a post-gastrula-stage embryo
consisting of three concentric germ layers - ectoderm, ventral mesoderm and
ventral endoderm and (B) a UV-sensitive event produces tissue of dorsal
type, including neural tissue, somites, notochord and dorsal endoderm, and
also produces dorso-ventral and anterior-posterior axes in the embryo. LPM,
____________________________________ Chapter 1____________________________________
cytoplasmic m ovem ents by disorganising m icrotubules, thereby p reven tin g
the norm al displacem ent of a dorsalising activity to the dorsal side of the
embryo (Gerhart et al., 1989; Houliston, 1994). It is likely that the m a n u a l
tipping of a 1-cell stage embryo that has been UV irradiated post-fertilisation
restores the displacem ent of a dorsalising activity to the dorsal side of the
embryo, thereby restoring the dorso-ventral axis.
A suprising number of m olecules that are expressed in early embryos h a v e
been identified as sufficient to 'rescue' (restore the dorso-ventral axis) the
U V -ventralised phenotype (Table 1.1). In such experim ents candidate
proteins are overexpressed by injecting mRNA, or dominant negative form s
of those m RNAs, into early cleavage stage U V-ventralised embryos. R esults
from experiments like this have suggested roles in dorsal axis formation for
all the m olecules listed in Table 1.1. For example Xwnt- 8 and a processed
form of Vg-1 (see later) are sufficient to induce the formation of a com plete
dorso-ventral axis (Thomsen and Melton, 1993), and activin and g o o s e c o i d
are capable of inducing an incomplete dorso-ventral axis (Cho et al., 1991;
Smith and Harland, 1991). It is important to note that m ost of the m o lecu les
show n in Table 1.1 (marked *) are absent or expressed at very low levels as
maternal m RNAs, and are only expressed strongly after the onset of zygotic
transcription (after MET). Thus, it seem s likely that these m o lecu les
function downstream of UV -sensitive maternal events and that som e of
them m ay represent the dorsalising activity of the organiser region. Further
research is necessary to distinguish w hich, if any, of these factors are
norm ally in volved in establishing the dorso-ventral axis, and of th ose
involved, w hat relationship they share w ith each other.
Recently som e of the essential com ponents of the dorsal, U V -sen sitiv e
pathw ay have been identified, p-catenin is maternally expressed at the R N A
and protein level (DeMarais and Moon, 1992). The protein is asym m etrically
localised, accum ulating in dorsal, but not ventral nuclei by the 16-32 cell
stage (Larabell et al., 1997). When ectopically expressed in UV ven tralised
embryos, (3-catenin is sufficient to mimic the endogenous
___________________________________Chapter 1____________________________________
Tab le 1.1 M olecules that rescue the UV phenotype, (adapted from
Heasman, 1997).
C o m p l e t e a x is R e f e r e n c e
*Xwnt- 8 (Smith and Harland, 1991)
"noggin (Smith and Harland, 1992a)
V g l (modified (Thomsen and Melton, 1993)
form)
Xwnt-8b (Cui et al., 1995)
"Xnrl and Xnr2 (Jones et al., 1995)
"Siam ois (Lemaire et al., 1995)
"chordin (Sasai et al., 1994)
I n c o m p l e t e
a x i s
"goosecoid (Cho et al., 1991)
activin (Smith and Harland, 1991)
"Xbra (Smith et al., 1991)
Xdsh (Sokol et al., 1995)
X w n tll (Ku and Melton, 1993)
dom. neg. BMP (Graff et al., 1994)
receptor
"Xnr4 (Joseph and Melton, 1997)
____________________________________ Chapter 1____________________________________
axes (Funayama et al., 1995; Guger and Gumbiner, 1995). Evidence to suggest
that p-catenin is vital for the establishm ent of the dorso-ventral axis in
X e n o p u s embryos comes from experiments that show that depletion of
maternal p-catenin transcripts from Xenopus oocytes prevents the formation
of a dorso-ventral axis (Heasman et al., 1994a). The dorsal deficiencies of
such embryos can be rescued by ectopic expression of p-catenin m R N A
(Heasman et al., 1994a).
p-catenin is a hom ologue of the D r o s o p h i l a protein Arm adillo (Peifer et al.,
1992). Arm adillo is a com ponent of the D r o s o p h i l a W ingless signal
transduction pathway (Fig 1.3), suggesting that p-catenin m ay itself lie
dow nstream of a X e n o p u s Wnt (Xwnt) signal. In keeping w ith this, the
dorsal deficiencies of p-catenin depleted embryos, that can be rescued by the
injection of p-catenin mRNA, cannot be rescued by Xwnt-8 , even th o u g h
Xwnt- 8 RNA has strong dorsalising activity in wild-type embryos (H easm an
et al., 1994b). It is not known which endogenous Xwnt, if any, fu nction s
upstream of p-catenin. One possible candidate is Xwnt8b which is m aternally
expressed and has potent axis forming ability (Cui et al., 1995) (Table 1.1). A
second Xwnt, X w n tll, is maternally expressed w ith transcripts localised to
the vegetal hem isphere. H ow ever it does not have strong axis form ing
capacity (Ku and Melton, 1993) (Table 1.1), w hich argues against a role for
X w n tll upstream of p-catenin (Ku and Melton, 1993).
It is likely that the ability of p-catenin to alter gene expression and cell fate
in volves its interaction with LEF/Tcf class transcription factors (Behrens et
al., 1996). One such factor, XTcf-3, is a Ubiquitous protein in early X e n o p u s
embryos, and it has been show n to associate with p-catenin using the yeast
two-hybrid system (Behrens et al., 1996; Molenaar et al., 1996). Injection of a
dom inant negative form of XTcf-3 into the dorsal side o f X e n o p u s embryos
blocks the formation of the endogenous dorso-ventral axis and, m oreover,
blocks the ability of ectopic p-catenin to induce secondary axes (Molenaar et
al., 1995), placing XTcf-3 alongside or downstream of p-catenin (Fig. 1.3). The
D r o s o p h i l a LEF/Tcf family member p a n g o l i n is required in v i v o for
Chapter 1
Drosophila Xenopus
Frizzled? Wg
ExtraceHular space
Cytoplasm
ZW3 (Shaggy)
PangoNn
Nucleus Pan-t- arm
Actlvalion of transcription (Engrailed Labial lAx)
Xfz? XWnt?
ExtraceHular space
Cytoplasm
Xdsh?
p-Catanin
Nucleus XTcf-3 + pcat
Activation of transcription (Siamois?, Goosecoid?)
XTcf-3
Figure 1.3. A comparison of the D r o s o ph i l a and X e n o p u s Wnt signal transduction pathways (adapted from Heasman, 1997). Wnt signalling
promotes the inactivation of zw3/GSK3, which leads to the accumulation of
Armadillo/(3-catenin. Armadillo/(3-catenin may then interact with HMG-box
transcription factors which regulate the expression of specific genes. ?
indicates uncertain components. Although pangolin is depicted here in the
________________________________ Chapter 1____________________________________
interacts w ith Armadillo and functions downstream of Arm adillo to
transduce the W ingless signal (Fig 1.3) (Brunner et al., 1997).
The serine-threonine kinase zeste-white-3 (zzv3) (Peifer et al., 1994) acts as an
inhibitor of the W ingless pathway in D r o s o p h i l a by altering the stability of
soluble Arm adillo (Pai et al., 1997). A dom inant-negative form of the
Xen opu s hom ologue of zw3, GSK3, causes ectopic axis formation w h e n
expressed in X e n o p u s embryos, but is unable to rescue p-catenin-deficient
embryos (Wylie et al., 1996). Thus, GSK3 appears to act in a similar upstream
position to p-catenin as zw3 does to Armadillo in D r o s o p h i l a (Fig 1.3)
(D om inguez et al., 1995; He et al., 1995; Pierce and Kimelman, 1996). N on e of
the endogenous signalling com ponents that lie upstream of GSK3 and
p-catenin are known.
It is not certain whether UV treatment of oocytes or embryos damages the
GSK3/p-catenin pathway directly or indirectly, although there is evidence to
suggest that the effects are indirect. Firstly, UV irradiation of the vegetal
poles of fertilised eggs causes the localisation of Siamois, a transcription
factor downstream of p-catenin, in the vegetal rather than the equatorial
region of the embryo (Brannon and Kimelman, 1996). This supports the idea
that UV irradiation of the egg disrupts cytoplasmic m ovem ents, rather than
the GSK3/p-catenin pathway directly. Secondly, normal developm ent can be
restored in UV irradiated fertilised eggs by manual tipping before the first
cell d ivision (Scharf and Gerhart, 1980). This suggests that the dorsalising
pathway is not irreparably damaged by UV treatment of fertilised eggs.
Thirdly, like wild-type oocytes, UV-irradiated oocytes can be dorsalised
(made to form exaggerated dorsal and anterior structures), by lith iu m
treatment at the 32-cell stage, w hich again suggests that the dorsalising
pathway is not irreparably damaged by UV treatment of the oocyte (Elinson
and Pasceri, 1989).
Classical experiments have show n that the exposure of embryos, betw een
early cleavage stages and MBT, to lithium ions causes dorsalisation (Kao et
al., 1986; Kao and Elinson, 1989). Lithium has been thought to act by
____________________________________ Chapter 1____________________________________
enzym e inositol m onophosphatase (Berridge et al., 1989). Indeed, there is
evidence to suggest that the IP3 pathway is in volved in dorso-ventral
patterning in Xenopus embryos (Busa and Gimlich, 1989; Kume et al., 1997).
Recent evidence, how ever, suggests that dorsalisation caused by lith iu m
treatment m ay be due to its effects on the GSK3/p-catenin pathway. L ith iu m
inhibits the enzym e, GSK3, resulting in the accum ulation of p-catenin and
consequently causes dorsalisation (Klein and Melton, 1996; Stambolic et al.,
1996).
M e s o d e r m ind u cin g f a c t o r s
A number of signalling m olecules have been identified that have the
capacity to respecify prospective ectodermal tissue to form m esoderm rather
than ectoderm. These factors mimic the activity of endogenous m esod erm
inducing factor(s) found in vegetal pole cells. They include members of the
fibroblast growth factor (FGF) family and members of the TGF-p superfam ily
(Smith, 1995), The exact nature of the endogenous m esoderm inducing
factor(s) remains obscure. H owever, the m esoderm inducing factors that
have been identified provide useful tools to investigate how m esoderm is
induced and patterned in early Xenopus embryos.
The best characterised m esoderm inducing factors are activin, a member of
the TGF-p superfamily purified from m edium conditioned by a Xenop us cell
line, and FGF (Sive, 1993; Smith, 1993). A ddition of either factor to isolated
animal pole explants, which normally form ectodermal tissue, results in
their differentiation into mesoderm al cell types. These two factors exert
different effects, w ith high concentrations of activin inducing dorsal and
axial structures, and FGF inducing ventral tissues (Green et al., 1990). In
terms of the three signal model, it did seem conceivable that FGF acts as the
ventral vegetal inducing signal and activin as the dorsal vegetal in d u cin g
signal.
FGF family m e m b e r s
____________________________________ Chapter 1____________________________________
m esoderm , indicating that FGF signalling is required in the formation of
m esoderm in Xenopus embryos (Amaya et aL, 1991; Amaya et al., 1993). This
suggested that an FGF family member may be a m esoderm inducing factor,
w hich w as supported by the findings that FGF-2 and eFGF are both expressed
m aternally (Kimelman et al., 1988; Isaacs et al., 1992). H ow ever, w hen a
dom inant negative form of an activin receptor is ectopically expressed in
X e n o p u s embryos, the resulting embryos contain no embryonic m esod erm at all (Hem m ati-Brivanlou and Melton, 1992), even though animal caps
expressing the same dominant negative activin receptor can still be induced
to form ventral m esoderm by FGF (Hem m ati-Brivanlou and M elton, 1992).
This indicates that FGF does not require an intact activin signalling pathway
to induce mesoderm. Together these data suggest that, in n orm al
d evelopm ent, FGF is not a m esoderm inducing factor; if it were, embryos
injected w ith truncated activin receptors should still be able to form the
ventral m esoderm that FGF is capable of inducing.
A lthough FGF may not be acting as a m esoderm inducing factor, evid en ce
suggests that FGF-signalling is required for normal m esoderm form ation.
Expression of a truncated FGF receptor prevents exogenous activin from
inducing the full spectrum of m esoderm al genes in animal caps (Cornell
and K imelm an, 1994; LaBonne and W hitman, 1994), suggesting that FGF
m ay have a role in activin-m ediated m esoderm induction in vivo.
A role for FGF signalling during gastrulation seem s likely. Several FGFs are
expressed during gastrulation (Isaacs et al., 1992; Tannahill et al., 1992),
including eFGF w hich is expressed in the blastopore lip and later in the
dorsoposterior m esoderm . Dissociated gastrula m esoderm cells cultured in
the presence of eFGF retain mesoderm al character that is otherwise lost in
culture (Isaacs et al., 1992), suggesting that FGF is in volved in m aintenance
of m esoderm al cell types. More recent experiments indicate that FGF is
in volved in m aintaining expression of the m esoderm specific gene,
X en op us Br a c h y ur y , {Xbra), and in the m aintenance of posterior m esoderm al cell types (Isaacs et al., 1994; Schulte-Merker and Smith, 1995).
The m ost convincing evidence for this has come from experim ents u sin g
____________________________________ Chapter 1____________________________________
receptor under the control of a CMV promoter w hich drives expression
from the early gastrula stage. In these embryos, m esoderm -specific genes,
like Xbra, were activated normally, but their expression was transient and
had disappeared by the mid-gastrula stage (Kroll and Amaya, 1996). These
transgenic embryos resembled those produced by injection of m R N A
encoding a truncated FGF receptor into fertilised eggs, supporting the idea
that FGF signalling is required for the m aintenance and differentiation of
m esoderm , rather than the induction of mesoderm.
It has been demonstrated that FGF exerts its effects through the Ras/ERK
(extracellular signal regulated kinase) MAPK (m itogen-activated protein
kinase) pathway (Gotoh et al., 1995; LaBonne et al., 1995; Um bhauer et al.,
1995). This pathway serves to link signals from the cell surface to
cytoplasmic and nuclear events. The Ras/ERK MAPK pathw ay is o n e
exam ple of what are generically termed 'MAPK' pathways. A ll such
pathways have a three-com ponent protein kinase cascade consisting of a
serine/th reon in e protein kinase, MAPKKK (mitogen-activated protein
kinase kinase kinase), w hich phosphorylates and activates a dual-specificity
protein kinase, MAPKK (mitogen-activated protein kinase kinase), w hich in
turn phosphorylates and activates another serine/th reon in e protein kinase,
MAPK, w hich translocates to the nucleus and regulates transcriptional
responses to growth factor stim ulation (Fig. 1.4) (Marshall, 1995). In the
Ras/ERK MAPK pathway Raf corresponds to MAPKKK, MEK corresponds to
MAPKK and ERK corresponds to MAPK (Fig. 1.4) . The expression of active
forms of MEK or MAPK in animal caps induces the formation of m eso d erm
of the kind elicited by FGF. Furthermore activated MEK, like FGF, induces
different types of m esoderm in a dose-dependent manner (Green et al., 1992;
Umbhauer et al., 1995). Expression of a X e n o p u s MAPK phosphatase, on the
other hand, blocks the induction of m esoderm by FGF and prevents the
form ation of notochord and posterior m esoderm in w hole em bryos
(Umbhauer et al., 1995). It appears that activation of MAPK (ERK kinase) is
Chapter 1
FGF Reœptor
tyrosine kinase
Cell
membrane
Inactive Ras Active RasGD P
P j G rb 2 GAPs
S O S
Raf
C m a pO
i
X Tr»neci#ofr\
fa c to rs /
i
Gene Expression
Figure 1.4. The FGF signalling pathway. Schematic representation of the
events that follow FGF binding to the tyrosine kinase FGF receptor. Ligand
binding stimulates the activity of the tyrosine kinase receptor w hich
activates ras by causing it to exchange GDP for GIF. Activated ras
phosphorylates the serine/threonine kinase raf, which in turn
phosphorylates MAP Kinase Kinase (MEK, for MAPK/ERK Kinase), w hich
Chapter 1
TGF-p su p e rfa m ily m e m b e rs
A ctivin
As m entioned above, activin induces animal pole tissue of blastula stage
X e n o p u s embryos to form m esoderm rather than ectoderm (Smith, 1995b) and ectopic expression of a dominant-negative activin receptor results in the
d evelopm ent of embryos containing little or no m esoderm (H em m
ati-Brivanlou and Melton, 1992). Together these data provided strong support
for a role for activin as an endogenous mesoderm inducing factor. H ow ever,
it has since been found that this particular truncated activin receptor does
not specifically block activin signalling, it also blocks that of BMP and a
processed form of Vg-1 (Kessler and Melton, 1994; Schulte-Merker et al.,
1994a). Thus, TGF-p signalling is vital for the formation of m esoderm in
X e n o p u s embryos, but the extent to w hich activin itself is in volved cannot be assessed from these experiments. More recently, use of a new design of
dom inant-negative activin receptor that specifically inhibits activin, but n o t
Vg-1 or any other know n mesoderm inducer, suggests that activin has an
essential role in early X e n o p u s developm ent (Dyson and Gurdon, 1996).
Embryos expressing this new design of dom inant-negative activin receptor
have reduced anterior structures, indicating that activin signalling is
required for normal developm ent. Looking at the time at w hich m esod erm
is induced, expression of this dominant negative receptor slightly delays the
onset of expression of the pan-mesodermal marker X e n op u s B r a c h y u r y , and
reduces initial expression of this gene to 20% of the w ild-type level (Dyson
and Gurdon, 1996), implicating activin in m esoderm induction. In keeping
w ith this, activin protein is present in X e n o p u s eggs and early em bryos
(Fukui et al., 1994; Oda et al., 1995).
In conflict w ith a role for activin as an endogenous m esoderm in d u cin g
factor in X e n o p u s embryos, overexpression of the activin-binding protein
follistatin, at levels sufficient to block the effects of ectopic activin
adm inistration, does not perturb m esoderm formation (Schulte-Merker et
____________________________________ Chapter 1____________________________________
m esoderm inducing factor it must be present in the embryo in a form
inaccessible to follistatin. In addition to this, m ouse embryos w ith
h om ozygous m utations in both activin A and activin B form m esod erm
norm ally (Matzuk et aL, 1995; Smith, 1995a), suggesting that activin
signalling is not required for m esoderm formation in the m ouse, and, by
extension, that it may not be required in any vertebrate embryo. W h eth er
activin is required for m esoderm induction in X e n o p u s is still not k n ow n .
H ow ever, if not activin itself, it does seem likely that an activin-like activity
is required for m esoderm induction in Xen opu s embryos.
C o n c e n t r a t i o n - d e p e n d e n t e ffe c ts of activin
One remarkable property of activin is that it is capable of inducing different
m esoderm al cell types at different concentrations. Low concentrations of
activin induce the formation of ventral and posterior cell types, and activate
the expression of ventral and posteriorly-expressed genes, such as Xbra,
w hile high concentrations induce the formation of dorsal and anterior
tissues and activate the expression of dorsal and anteriorly-expressed genes,
such as g o o s e c o i d (Green and Smith, 1990; Green et al., 1992; Green et al.,
1994; Gurdon et al., 1994; Symes et al., 1994; W ilson and M elton, 1994;
Gurdon et al., 1995). This property of activin was first demonstrated in
experiments using disaggregated-reaggregated animal cap explants (Green et
aL, 1994; Symes et aL, 1994; W ilson and Melton, 1994). If dispersed an im al
caps are exposed to a w ide range of concentrations of activin and analysed
after only a few hours, genes such as Xbra are expressed over a w ide range of
concentrations. H ow ever, if cells are reaggregated and cultured to n eu ru la
stages, Xbra expression becomes refined to a narrow w in d ow of activin
doses. Thus, precise interpretation of activin concentrations in
disaggregated-reaggregated animal cap explants requires cell-cell interactions
and takes som e time to occur (Green et aL, 1994; Symes et aL, 1994; W ilso n
and M elton, 1994). It appears that a more direct and rapid interpretation of
activin concentration is made in intact animal caps. W hen activin soaked
beads are sandwiched between animal cap explants, cells that are close to the
dorso-____________________________________ Chapter 1dorso-_____________________________________
anterior marker g o o s e c o i d , w hilst those that are further away activate Xbra
(Gurdon et aL, 1994; Gurdon et aL, 1995; Jones et aL, 1996a).
The sim plest interpretation of these results is that a concentration gradient
of activin is established in the responding animal cap explants, w ith in
w hich animal cap cells respond to predetermined thresholds of activin
activity by expressing particular m esoderm al markers and adopting
particular cell fates Fig. 1.5A). Alternatively it could be that the long-range
effects of activin are mediated by a relay of short-range signals. For exam ple,
activin may diffuse over a short range to induce the expression of g o o s e c o i d ,
but a second signal released by these go os ecoid- express ing cells is responsible
for inducing Xbra in adjacent cells (Fig. 1.5B).
Recently there have been several lines of evidence to support the m odel that
activin can act as a m orphogen and does not initiate a cell-cell sign allin g
relay in X e n o p u s tissues. Firstly, Gurdon et al. performed experim ents in
w hich non-com petent cells (endoderm from a stage-40 tadpole) w ere
inserted between activin m RNA-expressing vegetal tissue and responding
animal tissue (Gurdon et aL, 1994). These-non-com petent cells cannot
induce Xbra in an animal cap sandwich, nor can they express Xbra in
response to m esoderm inducing signals. Flowever, an Xl?m-inducing signal
from the activin expressing vegetal tissue was transmitted successfully
through the non-com petent layer to the animal tissue (Gurdon et aL, 1994).
This result was not dependent on the non-com petent cells synthesising a
secondary inducing factor, since the same results were obtained in
experiments in w hich the layer of non-com petent cells w as treated w ith the
protein synthesis inhibitor cycloheximide (Gurdon et aL, 1994). Secondly, it
was found that a constitutively active activin receptor (ALK-4*), in the
absence of ligand, was unable to generate long-range signalling in
conjugated animal caps (Jones et aL, 1996a). Thirdly, experim ents u sin g
dominant negative forms of an activin receptor dem onstrated that the m o st
distant responses to activin require a functional activin signalling pathway
(M cDowell et aL, 1997). Finally, it has been show n that activin can diffuse
Chapter 1
A. Morphogen gradient
Threshold 2
Threshold 1
Activin source
B. Signal relay
Signal 1
Signal 2
Activin source
Figure 1.5. Models for long-range signalling by activin (taken from Dale,
1997). (A) M orphogen gradient. In this model, activin diffuses th ro u g h o u t
the layer of cells and forms a concentration gradient, with high levels close
to the source and low levels distally. At the lowest level of activin, cells
differentiate as epidermis (yellow cells), but above a prespecified
concentration (threshold 1) they activate expression of Xb ra (red cells). As
the concentration increases, a second prespecified value (threshold 2) is
reached, and cells activate expression of gsc (blue cells). While this sam e
threshold may also repress Xbra, it is also possible that Xbra is repressed by
gsc. (B) Signal relay. There are many variants of this model but, in the one
illustrated here, activin (signal 1) diffuses over a short range and induces gsc;
activin also causes a second signal to be released (signal 2), which acts over a
short range to induce Xbra in adjacent cells. The essential feature of this
model is that Xbra is not induced by activin directly, but by a secondary