Indian J. Plant Physiol .• Vol. XXVII, No.1 1'1'.97-103 (January 1984)
PARTITIONING OF DRY MATTER IN SUGARCANE: INFLUENCE OF GROWTH AND DEVELOPMENT, GENOTYPE AND CROP STANDS
A.K. SHRIVASTAVA. N.K.KACKER AND M. SINGH Indian Institute ofSugareane Research. Lucknow. India
(Revised: April 4. 1914)
SUMMARY
Partitioning of dry matter in sugarcane, a key factor in productivity. was influenced by growth. genotype (within Saccharum hybrids) and different crop stands viz. autumn and spring planted cane and ratoons. Partitionina of assimilates to mother- shoot at mitialstages of growth and translocation of lIC_ labelled aSSimilates from mother- shoot to tillers indicated that for their initial growth and development. tillers depended . upon mother- shoot.
INTRODUCTION
Distribution of assimilates during growth and development-the parti tioning, is a key factor in crop productivity (Loomis et. al., 1976). Sugarcane being a C, plant, photosynthetic processes per se should not limit productivity, However, realizing efficient partitioning of assimilates to the components of economic value, appears to be a logical strategy for improving productivity of this crop. Present studies were conducted to understand the phenomenon of partitioning of dry matter as influenced by growth and development, genotype and various crop stands viz., autumn, spring planted cane and ratoons in sugarcane.
MATERIALS AND METHODS
Crop stands used for studying partitioning of dry matter as a function of growth and development in sugarcane varieties Co 419,.Co 997 and Co 1148 and genotypic variation in varieties Co L 29, Co I 67, Co J 75, Co 312, Co 419, Co 453, Co S 510, Co 997, Co 1148, Co 1158. LG 7255 and Co LK 7701, were raised by transplanting pre-germinated single bud settlings in the month of
ml
98 A.K. SHRiVASTAVA, N.K. KACKER AND M. SINGH
ratoons (previous crop harvested in Feb., 1980) (first, second and third ratoon of sugarcane cultivar Co 1148), plant samples were obtained from well managed fields at the Institute's farm.
Partitioning of dry matter was studied ia sugarcane varieties Co 419,Co 997
and Co 1148 in relation to growth at different stages of tillering
(if
initiation oftillering (30-35 days after transplanting), (ii) when the rate of tiller production
was maximum (70-80 days after transplanting), (iii) when there were maximum
number of tillers (100-110 days after transplanting), grand growth phase (140 150; 170- 180 days after transplanting) and maturity phase (210-220; 280- 290 days after transplanting). Similarly, magnitude of differences in partitioning of dry matter due to genotypes was studied in twelve varieties of sugarcane at tillering (100-11 0 days after transplanting), grand growth phase (170-180 days after transplanting) and maturi ty phase (280-290 days after transplanting). Partitioning of dry matter in various crop stands viz. autumn, spring planted canes and ratoons of sugarcane variety Co 1148 were studied at the age of ten months. There were three replications with two plants in each.
At various sampling occasions, plants were dug upto 45 em in depth (covering about 70% of root mass) and roots were washed in running water. The plants were then separated into various parts viz., mother-shoot. tillers, leaf laminae. leaf sheaths and roots. For dry matter estimation, 50 g of uniformly chopped material was kept in the oven at 10rC for two hrs and then dried at 85°C to a constant weight.
For radio-tracer studies to work out dependence of tillers on mother shoot in initial stages, forty days old plants (with mother-shoot having 3-4 leaves-a stage before tiller initiation) of varieties Co 419, Co 997 and Co 1148
were fed with 14Coa (10 I£Ci/IO mM). Tillers were numbered as they emerged.
When six tillers were produced, plants were uprooted, washed and separated into mother-shoot, first tiller, second tiller. third tiller and other tillers. They were oven dried and powdered to pass through 20 mesh. Fifty mg of this powder was used to estimate activity as described by Burne and Criswell. (1973).
RESULTS AND DISCUSSION
It is seen from table-l that during earlier stages of growth. leaf
laminae, leaf sheaths and roots were the major competing sinks. This was because plant initially needs sufficient leaf area for corbon assimilation. and more root mass to support water and mineral requirements. Van DiIlewijn 1952) 'reported that in sugarcane, during earlier stages of growth, leaf develop·
PARTITIONING OF DRY MATTER IN SUGARCANE 99
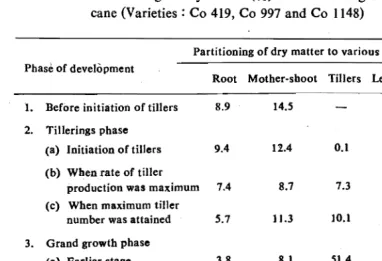
Table 1: Partitioning of dry matter (%) at various stages of growth in sugar cane (Varieties: Co 419, Co 997 and Co 1148)
Partitioning of dry matter to various morphologic components Phase of development
Root Mother-shoot Tillers Leaf laminae Leaf sheaths 1. Before initiation oftiIters 8.9 14.5 43.4 33.2 2. Tillerings phase
(a) Initiation of tillers 9.4 12.4 0.1 50.0 26.8 (b) Wben rate of tiller
production was maximum
(c) When maximum tiller number was attained
7.4 5.7 8.7 11.3 7.3 10.1 47.3 41.7 27.7 3J.3 3. Grand growth phase
(a) Earlier stage (b) Later stage
3.8 4.2 8.1 Il.9 51.4 55.5 3004 J8.9 16.6 9.2 4. Maturity
(a) When sugar accumulation states
(b) Harvest
3.7 1.9 13.7 J4.3 65.1 66.9 11.1 11.1 7.8 5.6
ment was considerably more than stalk growth. Before initiation of tiIIer~ng,
mother-shoot contributed 14.51% to total dry matter. The moment
tillering initiated, partitioning to mother-soot declined and after the forma tion of maximum number of tillers, prior to grand growth phase, partitioning to mother-soot decreased significanly. This with slight variation continued
upto elongation phase and thereafter increased. This would suggest that
for their initial growth, tillers depend upon mother-shoot and after that the plant enters into the grand growth phase, tillers as well as mother-shoot
start to grow independently. With increase in plant age, after the terminal
phase of growth, shoots become preferential sink for dry matter at the expense of other morphologic components like roots, leaf laminae and leaf sheaths. With further advancement of plant age, proportionate contribution of shoots other than mother-shoots compared to leaf laminae and leaf sheaths, increased
much faster so that at harvest, roots contribute 1.80%. leaf laminae
11.09%, leaf sheaths 5.54% to total dry weight of plants. Among stalks,
!Dother-shoot contributed 14.06
%
to total plant dry weight and 17.30%
to total100 A.K. SHRIVASTAVA, N.K. KACKER, AND M. SINGH
Table II : Partitioning of dry matter (in %) twelve cultivars of Saccharum
Mean parittioning (and its range) in Phase of development
Roots Mother-shoot
1. Tillering phase: Mean 4.35 9.73 .
Standard deviation 0.98 4.12
Range (%) 0 - - 2 2 - - - 4 4 - - 6 5---1010----15 15--20 3, 4, 7, I, 2, 5, 2, 3, 6, I, 4, 8. 9 5 10, 12 6, 8, 9, 7,9, 11,
12
II. Grand growth phase: Mean 1.53 13.46
Standard deviation 1.53 2.07
Range (%) 0 - - - 2 2 - - 4 4 - - 6 5-1010---15 15--20
4,1,95,6,1,2,3 2,3,4,6,1,5
7,10, 8,9, 10, 11.
11 12
In.
Maturity (Harvest) : Mean 2.4S 16.47Standard deviation 0.83 2.4S
Range (%) 0---22---44---6 10-15 15---20 20--25 1,4, 8, 2, 3, S, 6 2,5 1,3,4,6, 7 12 7,9.10, 8" 9, 10.
11 11
~
. -. ... -~ :-: , -
~
PARTITIONING OF DRY MATTER IN SUGARCA)lB 101
hybrids at tillering, grand growth phase and harvest
various morphologic components
Tillers Leaf laminae Leaf sheaths
20.05 39.75 26.79
4.49 5.53 4.93
10--15 15--20 20-25 30--35 35-40 40-45 lS-20 20--25 25--30 , 1,3 2. S. 6, 3,9 4, 5. 1. 6, 8. 2. 3, 7, , 4, 8, 10 1. 2. 3, 6,
7, 11. 12 10 9.11,12 I. 9. 10,
25--30 11,12
4,10
56.33 16.45 10.98
4.47 2.21 1.62
4S-55 55----65 6S-75 10-1S 1 5 - 2 0 2O-2S 5-10 10--1S U-20 1,2,3 4.5.6,7.8, 5,6.11 1,2,4.7. 3 S.6, 101.2,3,4.
9, 10, 11, 12 8.9, 10, 7,8,9,11,
12 12
61.80 12.20 6.19
5.42 2.25 1.20
50---60 60---70 70-80 5-1010-151S-20 4--S 5 - 8 8-10 3.6.7. 1,4,5, 2 4. S. 10. 1.2,3. 7 2, 4. 5. I. 6, 7 3,12
8,9,10, 11 6.8,9. 10,11
12,12 12
102 A.K. SHRIVASTAVA, N.K. KACKER, AND M. SINGH
stages of growth and development. Diffe rential partitioning of dry matter among different organs was therefore evident.
As regards dependence of tillers on mother-shoot, radio-tracer studies showed that after harvesting, plants fed with 14C02, maximum amount of acti vity in mother-shoot was 67.0, 65,0 and 41.0 % in Co 419, Co 997 and Co 1148, respectively. Movement of labelled assimilates (in terms of proportion of radio activity) from mother-shoot to tillers was of the same order in Co 419 and Co 997. In Co 1148, however, relatively more labelled assimilated were trans
located to tillers (Fig. I). This is similar to results of Burr et. al., (1957) and
flartt et. al., (1963) who have reported translocation throughout following exposure of a single leaf blade or a single stalk. This experiment gives an evi- ; dence that tillers in initial stages are dependent on mother-shoot. Similar results have been reported in wheat (Evans et.a I., 1975), rice (Ishizuka and Tanaka, 1963) and Spartina alterniflora (Lytle and Hull, 1980). But it is obvious from the partitioning data that at later stages the tiller as well as mother-shoot
become independent. Ifwe consider that tillers are morphologic and functio
nal units of sugarcane plants, mature tillers should become independent of parent culm or mother-shoot even though their vascular connections remain physiologically viable (as observed in other grasses by Williams (1964 and 1967) and St.- Pierre and Wright (1972).
It would be seem from table-2 that during tillering phase, mother
shoot, tillers, leaf laminae, leaf sheaths and roots contributed on an
average, 9.93, 20.05, 39.75,26.79 and 4.35%, respectively to total dry matter production; while, at grand growth phase the contribution of these components to total dry matter was 13.46, 56.33, 16.45, 10.98 and 1.53 per cent. At harvest, mother- shoot, leaf laminae, leaf sheaths and the roots contributed 16.47, 62.80,
12.20, 6.19 and 2.45% to total dry matter. It was evident that within the varie
ties, differences were more pronounced at tillering phase. This indicates that differences in tillering behaviour also determine partitioning of dry matter to some extent. Varying proportions of dry matter at different stages in various varieties indicate that activities of various competing sinks are controlled not only by growth and development but also by their genetic component.
It may be seen from Fig. 2 that the relative contribution of roots was
-!'
[:=:l
SPRINe
PLANT
ED
I
::
Q)
IIIIIm
AUTUMN
PLANTED
c
~
a;!V::~·:·:J
fiRST
RATOON
0
_
SECOND
RATOOM
.-
....
c
.
IIIDIII
THIRD
R
ATOO.N
....
~
...
c
Co~
....
~
~
30
i
z
0
...
Q.)20
a..
10
!liI
0
ROOT
Fig.1
:
Distribution
of
lie
among
the
different
plant
parts
of
sugarcane
varieties.
STALKS
LEAF
LAMIN.AE
LEAF
SHEATH
.
-
>
"...
....
IBM
MOTHER
SHOOT
mmm
FIRST
TILLER
> po.
_
SECOND
TIL
LER
~
c:J
THIRD
TILLER
l1li
OTHER
TILLERS
~
?
-
o
z
....
o
i"I
....
~
'0
40
~
~--
:l
o
~
~ en!
o
.-
....
c
::I ,.g-
....
10
.
o
0'
•
Co
419
Co997
Co
1148
Fig.
2:
Partitioning
of
dry
matter
among
different
plant
parts
(%)
in
different
crop
stands.
I
"
103
PARTITIONJNG OF DRY MATTER IN SUGARCANE
bution of leaf laminae and leaf sheaths increased at the cost of other com ponents like stalk. It thus appears that yields will be reduced considerably in third ratoon and this may not be economi cal. Variation in partitioning of dry ,matter indifferent crop stands of sugarcane indicate that activities of different sinks differ. Relatively more contribution of dry matter to roots in autumn compared
to'
spring and ratoon crops could be due to the fact, that the crop undergoes a longer period of low tempetatures. Mathur (1941) also observed that the growth of sugarcane was influenced by climatic conditions. In winter and spring ~heats also root growth was more dlle to longer period' of low tem peratures (Troughton, 1962).REFERENCES
Burr, 0.0., Hartt, C.E., Tanimoto, T., Takahashi, D. and Brodie, H.W. (1957). The circula tory system of sugarance plant, in Radio-Isotopes in Scientific Researcb. Pro. 1st (UNESCO) Int. Conf., 4 : 351-68
Evans. L.T., Wardlaw, I.F•.and Fiscber. R~A. (1975). Wheat, in Crop Physiology, Evanus, L.T. (cd), Cambridge Univ. Press, U.K. p. 101-49.
Hartt, C.E., Kortscbak, H.P., Forbes, A.J. and Burr. 0.0. (1t63). Translation of
lac
in sugar cane plant. PI. Physiol., 38: 30S·18.Hume, D.J. and Criswell, J.O. (1973}. Distribution and utilization of ICC-labelled assimilates in soybean. Crop Sci., 13 : 519-524.
Isbizuka, Y. and Tanaka, A. (1963) Studies on nutrio-pbysiolQ8Y of rice plant. Yokendo, Tokey. p.307.
;LQomis, R.S., Ng, E. and Hunt, W.P. (1976). Dynamics of development in crop production sys tems, in COl Metabolism and Plant Productivity, Burris, R.H. and Black, C.C. (eds), Univ. Park: Press, Baltimore, London and Tokyo, p. 26!1-86.
,Lytle, R. W. and Hull. R.J. (1980). Pbotoassimilate distribution in. Spanill(;l allernijlora I. Vege tative and fioral development. Acr. J., 71.: 933·7.
Mathur, R.N. (1941). Studies on tbe growth ofsugarcane and development ofsucroso at Saba jabanpur in relation to climatic conditions. Proc. 10th Ann. COli.,. STAI, 1941)
(original not available)
St.-Pierre, J.L. and Wright, M.J. (1972). Distribution of l·C pbotosyntbate in timotby.(Phleum parlen&e L.) during tbe vegetative growtb. Crop Sci., 12: 191-4.
Troughton, A. (1962). ' The ronts of temperate cereals (wbeat, barley oats and rye). C;omm. Bur. Pastures and Field Crops, Hlirley. p. 99.
Van Dtllewijn. C. (1952). Botany of sugarcane. tbe Chronica Botanica Co., U.S.A., p. 317. Williams, R.D. (1964). Assimilation and translocation in perennial grasses. Ann. Bot., 28:
419.26.