biological phenomena, is manifest by networks of interacting mediators that can be inferred to be interactive via various computational algorithms (11,12). This concept is especially important given the volume of data needed to de-fine drivers and biomarkers of complex diseases (10,13). Recently, we utilized computational modeling techniques with measured circulating inflammatory mediators to describe dynamic mediator networks that suggest novel interactions and possible biomarkers in the setting of trauma and sepsis (14–17). Using similar techniques, we have also demonstrated that PALF participants who survive with their native liver exhibit inflammatory network structures similar to LTx recipients (18).
Opportunities associated with computational modeling of inflammatory responses in PALF fall into two broad categories. First, patterns of inflammation there is evidence of immune or
inflam-matory dysregulation in acute liver failure (3–7) associated with outcome, we hypothesize that markers of immune or inflammatory dysregulation in PALF could be leveraged to predict outcome.
Computational modeling of complex biological systems is emerging as a tool to investigate the unpredictable behavior of biologic pathways (8), including those pathways operant in inflammation (9,10). Inflammation, similar to many complex INTrODUCTION
Pediatric acute liver failure (PALF) is a complex, dynamic and rapidly evolving clinical syndrome whose onset is not completely understood (1,2). Identifying patients likely to recover spontaneously or die without liver transplant (LTx), or not benefit from LTx, is necessary to in-form LTx decisions. Unfortunately, clini-cal or laboratory biomarkers that predict outcome reliably are not available. As
acute Liver Failure
Ruben Zamora,
1,2Yoram Vodovotz,
1,2Qi Mi,
1Derek Barclay,
1Jinling Yin,
1Simon Horslen,
3David Rudnick,
4Kathleen M Loomes,
5and Robert H Squires
61Department of Surgery, University of Pittsburgh, Pittsburgh, Pennsylvania, United States of America; 2Center for Inflammation and Regenerative Modeling, McGowan Institute for Regenerative Medicine, University of Pittsburgh, Pittsburgh, Pennsylvania, United States of America; 3Seattle Children′s Hospital, Seattle, Washington, United States of America; 4St. Louis Children′s Hospital, St. Louis, Missouri, United States of America; 5Children′s Hospital of Philadelphia, Philadelphia, Pennsylvania, United States of America; and 6Department of Pediatrics, University of Pittsburgh, Pittsburgh, Pennsylvania, United States of America
The absence of early outcome biomarkers for pediatric acute liver failure (PALF) hinders medical and liver transplant decisions. We sought to define dynamic interactions among circulating inflammatory mediators to gain insights into PALF outcome sub-groups. Serum samples from 101 participants in the PALF study, collected over the first 7 d following enrollment, were assayed for 27 inflammatory mediators. Outcomes (spontaneous survivors [S, n = 61], nonsurvivors [NS, n = 12] and liver transplant patients [LTx, n = 28]) were assessed at 21 d post-enrollment. Dynamic interrelations among mediators were defined using data-driven algorithms. Dynamic Bayesian network inference identified a common network motif, with HMGB1 as a central node in all patient subgroups. The networks in S and LTx were similar, and differed from NS. Dynamic network analysis suggested similar dynamic connectivity in S and LTx, but a more highly interconnected network in NS that increased with time. A dynamic robustness index calculated to quantify how inflammatory network connectivity changes as a function of correlation stringency differentiated all three patient subgroups. Our results suggest that increasing inflammatory network connectivity is associated with nonsurvival in PALF and could ultimately lead to better patient outcome stratification.
Online address: http://www.molmed.org doi: 10.2119/molmed.2016.00183
Address correspondence to Yoram Vodovotz, Department of Surgery, University of Pittsburgh, W944 Biomedical Sciences Tower, 200 Lothrop Street, Pittsburgh, PA 15213, USA. Phone: 412-648-3758; Fax: 412-383-5946; E-mail: [email protected].
and biological interactions may be as-sociated with clinical outcomes. Current models to predict death or spontaneous survival in PALF using clinical, biochemi-cal and demographic data are inadequate (19,20), and it is unlikely that a single inflammatory mediator determined at a single time point along the dynamic PALF trajectory would be sufficient to reliably predict outcome (6). Therefore, computational modeling analyses could fill a critical void in medical decisions. Second, identifying potential regulatory points in dynamic networks of immune or inflammatory dysregulation could identify opportunities for selective therapeutic interventions.
To address these two opportunities, we hypothesize that key mediators involved in inflammatory interactions, as well as subgroup-specific dynamic patterns of these mediators, can distinguish rele-vant patient outcomes. Identifying these dynamic interconnections of selected immune and inflammatory cytokines could serve to distinguish among PALF outcome groups, enhance LTx decision- making, identify disease mechanisms and suggest novel therapeutic oppor-tunities for PALF. Herein, we sought to extend our previous studies in PALF (18) to determine if a broader and deeper computational analysis, based on a larger dataset, would yield further insights.
MaTErIaLS aND METHODS
Selection of PaLF Patients
This study was approved and carried out in accordance with the guidelines of the Institutional Review Boards at each participating institution listed in Acknowledgments, and the National Institutes of Health provided a Certifi-cate of Confidentiality. Participants were enrolled in the PALF Study if they met the entry criteria (1) (see Supplementary Materials and Methods) and after written informed consent was obtained from the children’s parents or guardians. After enrollment, demographic and clinical data were recorded daily for up to 7 d,
with a single daily serum sample for research scheduled to be collected on the calendar day of enrollment (d 0) or with the first morning blood draw following enrollment and daily for up to 7 d (d 1–d 7), or until death, LTx or discharge from hospital. Outcomes were assigned on d 21 as survival without LTx (S), death without LTx (NS) or LTx. Since not all patients in each outcome group had research samples obtained on the same days, patients were selected for this analysis if they had at least three daily samples with at least 100 μL of serum available.
assays of Inflammatory Mediators We measured 25 cytokines and chemokines that serve as biomarkers for the complex inflammatory response using the Luminex™ 100 IS system (Luminex) and the Human 25-plex® Luminex beadset (Millipore). This antibody bead kit includes eotaxin, granulocyte- macrophage colony-stimulating factor (GM-CSF), interferon (IFN)-α2, IFN-γ, interleukin (IL)-1β, IL-1 receptor antago-nist (IL-1RA), IL-2, soluble IL-2 receptor α chain (sIL-2Rα), IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12p40, IL-12p70, IL-13, IL-15, IL-17A, IFN-γ-inducible protein of 10 kDa (IP-10; CXCL10), monocyte chemotactic protein-1 (MCP-1; CCL2), monokine induced by γ-interferon (MIG; CXCL9), macrophage inflam-matory protein (MIP)-1α, MIP-1β, and tumor necrosis factor (TNF)-α. In addition, we assayed HMGB1 using a commercially available enzyme-linked immunoabsorbent assay (ELISA) (Shino-Test) and the nitric oxide (NO) reaction products NO2– + NO
3– using the nitrate reductase method (Cayman Chemical).
Statistical analysis and Data-driven Modeling
Our analytic strategy was to apply a stepwise series of statistical and data- driven modeling techniques aimed at discovering principal drivers, intercon-nected networks and potential key regu-latory nodes based on data on systemic
inflammation. For the analyses, we used the raw data (time series for each indi-vidual) without applying any transfor-mation. We detail these analyses below.
Dynamic Bayesian network (DBN) inference was carried out to define the most likely single network structure that best characterizes the dynamic interactions among systemic inflamma-tory mediators across all time points, in the process suggesting likely feedback structures that define central nodes. The networks might also suggest possi-ble mechanisms by which progression of the inflammatory response differs within a given patient subgroup. In this analysis, time courses of unprocessed inflammatory mediator measurements from each patient were used as input for a DBN inference algorithm, imple-mented in Matlab® essentially as de-scribed previously for gene array data (21) and modified by our group for the study of systemic acute inflammation (18,22,23).
total number of mediators analyzed. The total number of connections represents the sum of the number of connections across all time intervals for all patients in a given subgroup.
Dynamic robustness indexwas calculated to quantify how network connectivity (as defined by DyNA) changes as a function of correlation stringency in each patient subgroup over seven consecutive time periods. This metric was calculated for each time period and patient subgroup using the lowest (0.70) and highest (0.95) correlation stringency and network connectivity (C) as follows: (C0.7-C0.95)/(0.95–0.7).
All supplementary materials are available online at www.molmed.org.
rESULTS
Clinical Criteria and Demographics of PaLF Participants
There were 1,144 patients enrolled in the PALF study at the time of this analysis. We restricted our cohort to those with at least three daily samples containing at least 100 μL of serum and identified 101 participants: S (n = 61), NS (n = 12) and LTx (n = 28). The demo-graphic data for study participants are shown in Supplementary Table S1. Non-survivors were younger than Non-survivors and LTx recipients. The schematic of the statistical and computational methods and the rationale for their use in this study are depicted in Supplementary Figure S1.
Differential Trajectories of
Inflammatory Mediators in Different PaLF Outcomes
Time courses of 27 mediators (shown in Supplementary Figure S2) identified median levels of eotaxin, IL-6, IL-8, IL-10, IL-15, IP-10, MCP-1, MIG and NO (measured as NO2–/NO
3–) with distributions that significantly differed (P < 0.05) among the three PALF subgroups (Supple-mentary Table S2).
DBN analysis Identifies a Common Signature in PaLF
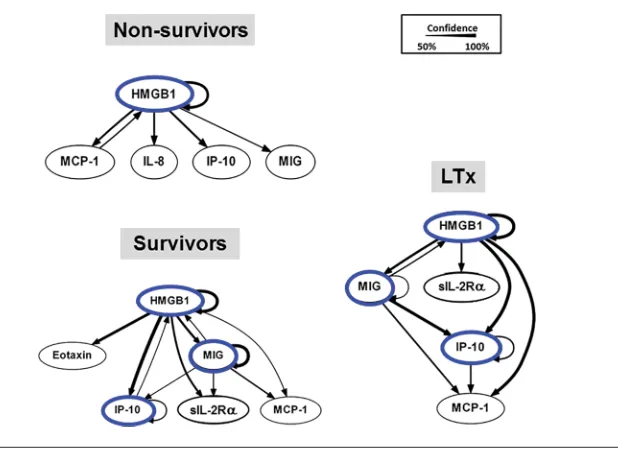
With a much larger cohort than our previous report (18), we utilized DBN inference to determine if mediator feed-back structures and possible regulatory architectures in inflammatory networks of S, NS and LTx recipients could be discerned from the time courses of circu-lating inflammatory mediators in PALF participants. A key feature of dynamic networks is the presence of mediator feedback, which can be inferred algo-rithmically, so we focused on mediators that exhibit self-feedback as central nodes, similar to our recent studies in PALF, trauma and sepsis (15,17,18,24). Results of this analysis on each of the three PALF subgroups are shown in Figure 1. Though data were segregated by outcome group before being subjected to DBN inference, the algorithm did not make assumptions about the connectiv-ity of the network in any outcome group. In NS, DBN inference suggested a pri-mary network driven by a core motif of HMGB1, which was inferred to affect the downstream production of MCP-1, IL-8, IP-10 and MIG. In contrast, in both the S and LTx groups, the DBN pattern sug-gested a network regulated by switching between HMGB1 and the chemokines IP-10 and MIG, each of which drives its own expression. In the S and LTx groups, HMGB1 was inferred to affect the downstream production of sIL-2Rα and MCP-1. Collectively, and in agree-ment with our prior study (18), we found similar inflammatory networks in the S and LTx groups that differ from the NS, which suggests the presence of a com-mon inflammatory signature associated with clinical outcomes in liver inflamma-tory diseases such as PALF.
Dynamic Network analysis of Circulating Inflammatory Mediators Serves to Differentiate PaLF Patients
We next sought to define, in a more granular fashion, the time evolution of dynamic networks of systemic inflamma-tion of PALF participants. Accordingly, we employed DyNA (14,15) to discern
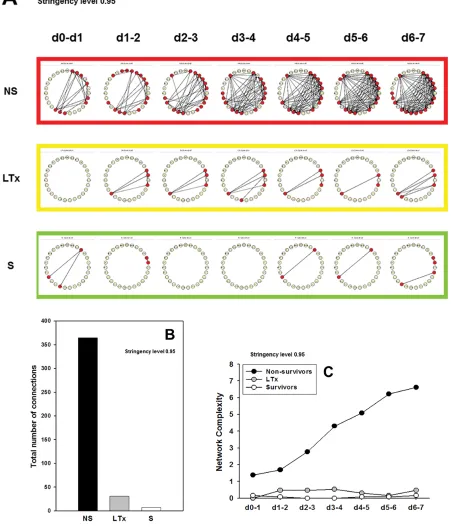
and compare the interconnections among inflammatory mediators in the PALF subcohorts over defined ranges of time. We hypothesized that patients with worse clinical outcomes would have more robust dynamic networks of inflammation, in line with the concept of pathology driven by self-sustaining inflammation. To quantify the robust-ness of a given dynamic network, we performed DyNA using increasing stringency levels from 0.7 (a correlation value commonly used to characterize trajectories that move in parallel either up or down) (14,15) up to 0.95, as shown in Supplementary Figure S3. This anal-ysis resulted in a marked decrease in the number of significant mediators and connections in all groups, though with greater complexity in NS. Subsequently, we repeated the detailed DyNA using a stringency level of 0.95 (Figure 2). DyNA showed an overall low and sim-ilar network pattern and total number of connections over all time intervals in the S and LTx groups. However, a much more complex network pattern (Figure 2A) and higher total number of connections (Figure 2B) were observed in NS. Fur-thermore, the dynamic connectivity of systemic inflammation in NS rose con-stantly over time, while remaining essen-tially unchanged in S and LTx patients (Figure 2C).
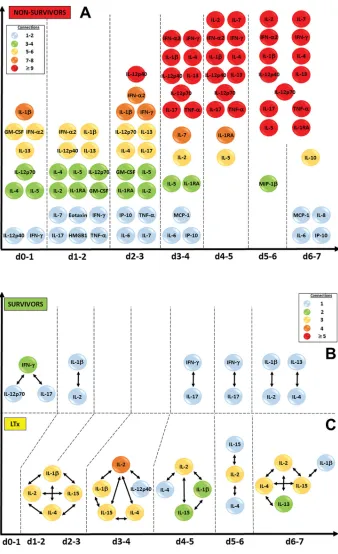
stringency in the dynamic networks observed in PALF patients, reasoning that a higher degree of inflammatory network robustness reflects a greater resistance to self-resolution, and likely to therapy as well. While DyNA further reinforced the similarity between S and LTx suggested by DBN inference and demonstrated a very low degree of net-work connectivity across time at a high stringency of correlation, the robustness index clearly differentiated among the three PALF subgroups. Our findings suggest either that inflammation ob-served in S and LTx is at a chronic and relatively stable level, or that small flares of inflammation are resolved. In contrast, NS exhibited a strikingly different pat-tern: network connectivity was already higher than S and LTx upon initiation of the study and rose steadily through d 7. Examination of the connected me-diators in NS suggests a major inflection point of systemic inflammation between d 2 and d 3 after enrollment, leading to a large increase in connectivity out to d 7. Chemokines and DAMPs such as HMGB1 appear to play an early role in nonsurvivors, with later inflammation involving Th1, Th2 and Th17 pathways. These findings suggest that a large num-ber of inflammatory pathways and mech-anisms are operant in patients destined not to survive PALF, leading to a state of self-sustaining or self-propagating in-flammation driven by both innate and T cell–dependent mechanisms.
Our computational analysis using established dynamic network discovery algorithms (DBN, DyNA) also repeatedly points to a number of mediators (e.g., HMGB1, MIG, IP-10, IL-6 and MCP-1) as possible biomarkers in PALF. These anal-yses suggest that dynamic patterns of sys-temic inflammation could reflect networks, which in turn identify key inflammatory mechanisms operant in, or characteristic of, PALF. Our present findings support and extend prior findings regarding dy-namic networks of systemic inflammation in PALF, namely that S and LTx exhibit similar dynamic networks that differ from those of NS PALF patients (18).
We next hypothesized that network robustness was higher in NS versus S and LTx, and that this feature underlies the time-dependent rise in inflammatory connectivity in NS. As shown in Figure 4, this hypothesis was supported by analy-sis of a dynamic robustness index, calcu-lated to quantify how inflammatory network connectivity changes as a func-tion of correlafunc-tion stringency, which showed that NS had higher network robustness at all time intervals compared with S and LTx. The robustness index was lowest in LTx, intermediate in S and highest in NS.
Because nonsurvivors, as a group, were significantly younger than survivors (see Supplementary Table S1), we considered age as a possible confounder when ana-lyzing mediator time courses. To rule out the role of age, first we selected all sur-vivors (n = 45 patients) whose ages, as a group, were not statistically significantly different (P = 0.053) from those of the nonsurvivors as shown by the median and Q1–Q3 [S: 3.3, 1.1–9.9]. Similar to the
whole S cohort (n = 61 patients), in the age-matched survivors DyNA demon-strated a significant decrease in the total number of connections and network complexity compared with NS (Figure 5). Similarly, as for the whole S cohort, the DBN pattern of this smaller group sug-gested a network regulated by switching between HMGB1 and the chemokine MIG, each of which drives its own ex-pression (Supplementary Figure S4).
DISCUSSION
Several mediators emerged as possible drivers or biomarkers of interest in PALF based on our combined analyses: HMGB1, MIG, IP-10, IL-6 and MCP-1. HMGB1 not only plays a critical intracellular role as a DNA chaperone, chromosome guardian, autophagy sustainer and protector from apoptotic cell death, but also senses and coordinates the cellular stress response as the prototypic alarm/danger molecule (26). Our network analyses suggest an inflammatory chain reaction that includes HMGB1, which exhibits self-feedback behavior that may act as a central node in all PALF subgroups. In this dynamic network, HMGB1 is inferred to interact with the chemokines MIG, IP-10 and MCP-1. Elevated MIG and IP-10 mRNA has been reported in liver failure (27,28), and circulating levels of both chemokines have been implicated in chronic hepatitis C (29). We have shown that MCP-1 is a central node in hepatocyte inflammatory responses to stress, a role perhaps related to the ability of circulating MCP-1 levels to segregate outcomes in human blunt trauma (15). Interestingly, in addition to our prior demonstration of the central role of MCP-1 in stressed hepatocytes (15), greatly increased MCP-1 levels were observed in livers from patients with fulminant hepatic failure (30). We demon-strated that IL-6 was elevated but not con-nected to other inflammatory mediators in dynamic networks in mice undergoing trauma/hemorrhage (14). In this study, IL-6 was also elevated with very low connectivity in NS, but not in S and LTx, suggesting that inflammatory hypocon-nectivity, like hyperconhypocon-nectivity, might be a hallmark of pathology.
Increased HMGB1 levels have been reported in patients with ALF, sug-gesting that HMGB1 is released from injured liver tissue (31). Although a specific role for HMGB1 in PALF has not previously been reported, the ob-served HMGB1-centered network in all PALF groups, and finding HMGB1 to be present in early inflammation networks prior to a massive increase in network connectivity in NS, might suggest a com-mon DAMP/chemokine inflammatory Figure 3. Significant circulating inflammatory mediators and connections in PALF patients
process driving liver injury in PALF. Thus, HMGB1 might represent a novel therapeutic target in PALF.
There are some unavoidable limita-tions to our study. First, unlike other
clinical settings, such as traumatic injury, we were unable to determine the onset of systemic inflammation in PALF pa-tients, as the onset of PALF cannot be determined. With only 8 d of samples
analyzed, changes in dynamic networks before or after this time period still need to be identified. The same applies to samples from participants with short, mild events or severe, rapid progression to death or transplantation who were excluded from this analysis, given the requirement for at least three available samples. In addition, our analysis was not able to determine if individuals can transition between a “survival” network to a “nonsurvival” network. Unsuper-vised hierarchical clustering based on unprocessed inflammatory mediator data (not shown) could not segregate patient subgroups, raising a cautionary note that standard statistical analysis of mediators singly or as a group might be of limited use for predicting patient out-comes. Finally, although we identified a set of mediators in the context of dy-namic networks that could potentially be considered as outcome biomarkers, we acknowledge the difficulties involved in translating these complex analyses and hypotheses into a diagnostic test.
The present results validate the core findings from our previous study of PALF (18) and also suggest the pres-ence of overly robust self-sustaining and highly connected inflammation networks in nonsurvivors. Despite a heterogeneous inflammatory response in PALF participants, these studies suggest that a network-based analysis has the potential to clearly segregate spontaneous survivors and LTx recip-ients from nonsurvivors, and sponta-neous survivors from LTx recipients. While similarities between S and LTx groups also suggest that some of the LTx patients might have a “survivable” disease, differences with the NS group points to a distinct group of patients who might not benefit from LTx. Our study also suggests a common inflam-matory network regulated by HMGB1 and the chemokines IP-10 and MIG, which might represent either a gen-eral “liver signature” in inflammatory conditions such as PALF or the central contribution of the liver to other inflam-matory diseases.
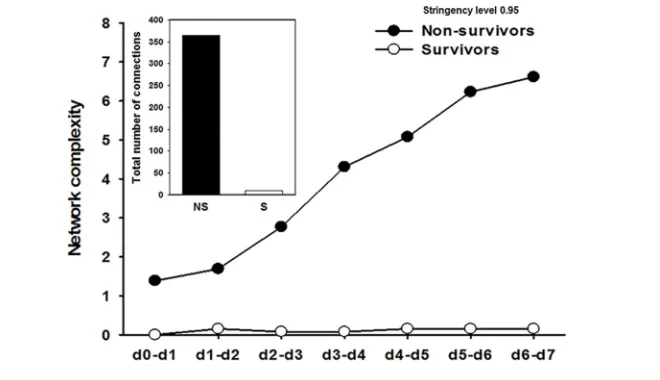
Figure 5. Dynamic network analysis (DyNA) of circulating inflammatory mediators in PALF patients (age-matched survivors versus nonsurvivors). Circulating inflammatory media-tors in serum samples from PALF spontaneous survivors (S, n = 45 patients) that closely matched nonsurvivors (NS, n = 12 patients) were measured and DyNA (stringency level = 0.95) was performed, as described in Materials and Methods. The figure shows the network complexity during each of the seven time frames calculated, as described in Materials and Methods, and the total number of connections for each group of patients over all time intervals (inset).
CONCLUSION
Identifying immune/inflammatory networks that reflect dynamic changes in the inflammatory response could lead to opportunities for diagnosis or directed therapeutic intervention, enhance liver transplant decisions and improve patient outcomes in PALF. In this regard, our studies also identified biomarkers such as HMGB1 as potential therapeutic tar-gets in PALF. More generally, the use of novel computational methodology could streamline studies of other inflammatory diseases.
aCKNOWLEDGMENTS
This work was supported by National Institutes of Health/National Institute of Diabetes and Digestive and Kidney Diseases grant UO1 DK072146 (A Multi-Center Group to Study Acute Liver Failure in Children). We would like to thank Regina M Hardison, Barbara Pavliakova Walters, Sharon M Lawlor and Steven H Belle from the Epidemiology Data Center (University of Pittsburgh) for their invaluable help. This work was supported by A Multi-Center Group to Study Acute Liver Failure in Children and the collaborative efforts of the following current and former principal and co- investigators of the Pediatric Acute Liver Failure Study (by site):
University of Pittsburgh: Robert H Squires, MD, and Benjamin L Shneider, MD; Cincinnati Children′s Hospital: John Bucuvalas, MD, and Mike Leonis, MD, PhD; Lurie Children′s Hospital of Chicago: Estella Alonso, MD; University of Texas Southwestern (Dallas): Norberto Rodriguez-Baez, MD; Seattle Children′s Hospital: Karen Murray, MD, and Simon Horslen, MB, ChB; Children′s Hospital Colorado (Aurora): Michael R Narkewicz, MD; St Louis Children′s Hospital: David Rudnick, MD, PhD, and Ross W Shepherd, MD; University of California at San Francisco: Philip Rosenthal, MD; Hospital for Sick Children (Toronto, Canada): Vicky Ng, MD; Riley Hospital for Chil-dren (Indianapolis, IN): Girish Subbarao, MD; Emory University (Atlanta, GA): Rene Romero, MD; Children′s Hospital
of Philadelphia: Elizabeth Rand, MD, and Kathy Loomes, MD; Kings
College–London (England): Anil Dhawan, MD; Birmingham Children′s Hospital (England): Dominic Dell Olio, MD, and Deirdre A Kelly, MD Texas Children′s Hospital (Houston): Saul Karpen MD PhD; Mt. Sinai Medical Center (Miami Beach, FL): Nanda Kerkar, MD; Univer-sity of Michigan (Ann Arbor): M James Lopez, MD, PhD; Children′s Hospital Medical Center (Boston): Scott Elisofon, MD, and Maureen Jonas, MD; Johns Hopkins University (Baltimore, MD): Kathleen Schwarz, MD; Columbia Univer-sity (New York, NY): Steven Lobritto, MD. We acknowledge the dedicated research coordinators at each site.
DISCLOSUrE
The authors declare that they have no competing interests as defined by
Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
rEFErENCES
1. Squires RH Jr, et al. (2006) Acute liver failure in children: the first 348 patients in the pedi-atric acute liver failure study group. J. Pediatr. 148:652–58.
2. Squires RH Jr. (2008) Acute liver failure in children. Semin. Liver Dis. 28:153–66. 3. Lee WM. (1993) Acute liver failure. New Engl.
J. Med. 329:1862–72.
4. Brown KE, Tisdale J, Barrett AJ, Dunbar CE, Young NS. (1997) Hepatitis-associated aplastic anemia. New Engl. J. Med. 336:1059–64. 5. Rolando N, et al. (1990) Prospective study of
bacterial infection in acute liver failure: an analysis of fifty patients. Hepatology. 11:49–53.
6. Bucuvalas J, et al. (2013) Immunophenotype predicts outcome in pediatric acute liver failure. J. Pediatr. Gastroenterol. Nutr. 56:311–15. 7. Lee WM. (2003) Acute liver failure in the United
States. Semin. Liver Dis. 23:217–26. 8. Kitano H. (2002) Systems biology: a brief
overview. Science. 295:1662–64.
9. Vodovotz Y, Csete M, Bartels J, Chang S, An G. (2008) Translational systems biology of inflam-mation. PLoS Comput. Biol. 4:e1000014.
10. Vodovotz Y, Billiar TR. (2013) In silico modeling: methods and applications to trauma and sepsis. Crit. Care Med. 41:2008–14.
11. Alon U. (2007) Network motifs: theory and experimental approaches. Nat. Rev. Genet. 8:450–61.
12. Zak DE, Aderem A. (2009) Systems biology of innate immunity. Immunol. Rev. 227:264–82. 13. Gough NR, Yaffe MB. (2011) Focus issue:
con-quering the data mountain. Sci. Signal. 4:eg2. 14. Mi Q, et al. (2011) A dynamic view of trauma/
hemorrhage-induced inflammation in mice: prin-cipal drivers and networks. PLoS.One. 6:e19424. 15. Ziraldo C, et al. (2013) Central role for MCP-1/
CCL2 in injury-induced inflammation revealed by in vitro, in silico, and clinical studies. PLoS One. 8:e79804.
16. Almahmoud K, et al. (2015) Impact of Injury Severity on Dynamic Inflammation Networks Following Blunt Trauma. Shock. 44:101–09. 17. Almahmoud K, et al. (2015) Prehospital
Hypo-tension Is Associated With Altered Inflammation Dynamics and Worse Outcomes Following Blunt Trauma in Humans. Crit. Care Med. 43:1395–1404. 18. Azhar N, et al. (2013) Analysis of serum
inflam-matory mediators identifies unique dynamic networks associated with death and spontaneous survival in pediatric acute liver failure. PLoS One. 8:e78202.
19. Sundaram V, et al. King’s College Hospital Cri-teria for non-acetaminophen induced acute liver failure in an international cohort of children. J. Pediatr. 162:319–23, e311.
20. Lu BR, et al. (2013) Evaluation of the liver injury unit scoring system to predict survival in a mul-tinational study of pediatric acute liver failure. J. Pediatr. 162:1010–16, e1011–14.
21. Grzegorczyk M, Husmeier D. (2011) Improve-ments in the reconstruction of time-varying gene regulatory networks: dynamic programming and regularization by information sharing among genes. Bioinformatics. 27:693–99.
22. Emr B, et al. (2014) Removal of inflammatory ascites is associated with dynamic modification of local and systemic inflammation along with prevention of acute lung injury: in vivo and in silico studies. Shock. 41:317–23.
23. Zaaqoq AM, et al. (2014) Inducible protein-10, a potential driver of neurally controlled interleukin-10 and morbidity in human blunt trauma. Crit. Care Med. 42:1487–97.
24. Namas RA, et al. Temporal Patterns of Circulating Inflammation Biomarker Networks Differentiate Susceptibility to Nosocomial Infec-tion Following Blunt Trauma in Humans. Ann. Surg. 263:191–98.
25. Tanaka R, Csete M, Doyle, J. (2005) Highly optimised global organisation of metabolic net-works. Syst. Biol. (Stevenage). 152:179–84. 26. Kang R, et al. (2014) HMGB1 in health and
disease. Mol. Aspects Med. 40:1–116.
27. Koniaris LG, et al. (2001) Cytokine-responsive gene-2/IFN-inducible protein-10 expression in multiple models of liver and bile duct injury suggests a role in tissue regeneration. J. Immunol. 167:399–406.
during acute liver injury through the induction of CXCR2 on hepatocytes. J. Immunol. 167: 7077–83.
29. Larrubia JR, Benito-Martinez S, Calvino M, Sanz-de-Villalobos E, Parra-Cid T. (2008) Role of chemokines and their receptors in viral persistence and liver damage during chronic hepatitis C virus infection. World J. Gastroenterol. 14:7149–59.
30. Czaja MJ, Geerts A, Xu J, Schmiedeberg P, Ju Y. (1994) Monocyte chemoattractant protein 1 (MCP-1) expression occurs in toxic rat liver injury and human liver disease. J. Leukoc. Biol. 55:120–26.
31. Oshima G, et al. (2012) Increased plasma levels of high mobility group box 1 in patients with acute liver failure. Eur. Surg. Res. 48:154–62.