Who’s in charge now? Coral functional group shifts along the Belizean
Barrier Reef over 20 years
Zachary Locklear, John Bruno, Catie Alves, Melanie McField, Richard Aronson
Abstract
Having diverse functions within ecosystems helps increase ecosystem health, yet
maintaining that functionality can prove difficult when conditions are stressful or functional
redundancy fails. To test whether or not functional diversity has been altered, c
oral reef functional group abundances were tracked over three decades along the Belizean Barrier Reef System and modeled against human impacts and protection status to see if localized impacts were having an effect. Competitive, stress-tolerant, and weedy corals based on Grime’s C-S-R framework were grouped and tracked through time. Stress-tolerant species experienced a strong decline, while weedy and competitive species had a much smaller decline. These shifts have led to a historically unprecedented co-dominance between stress-tolerant species and weedy species in Belize, which has decreased the functional diversity of the reef through the loss of the key reef-building taxa Orbicella. Localized impacts were not found to be important for any functional groups, and future work should include regional and global factors such as sea surface temperature and temperature anomalies to understand what might have driven this shift in functional diversity.Introduction
A central paradigm in community ecology is that ecosystems benefit from a diverse assemblage of species, via ann in increase insing ecosystem functionality. This is especially true for foundation species, as they form the basis of an ecosystem and have a huge effect on the health of an ecosystem. For instance, in grassland plant communities, increased grass species diversity can help increase primary productivity, nutrient cycling (Hector et al., 1999), and plant biomass (Tilman et al., 1997). These processes facilitate a more diverse ecosystem further up the trophic web through niche specialization and resource partitioning (Hooper et al., 2005). This is particularly important for threatened habitats such as coral reefs, as some evidence indicates that greater diversity helps lead to increased resistance and resilience to disturbance through functional redundancy (Pillar et al., 2013; Walker, Kinzig, & Langridge, 1999). As coral reefs continue to face uncertainty in the face of climate change and overfishing (O. Hoegh-Guldberg et al., 2007; T. P. Hughes et al., 2003), understanding how and why patterns of coral diversity have changed is crucial for developing management plans, despite the controversy over the effectiveness of local management in the face of climate change (Cox, Valdivia, McField, Castillo, & Bruno, 2017; Huntington, Karnauskas, & Lirman, 2011). In particular, measuring coral diversity against parameters such as temperature anomalies, human population density, and management level could help determine what exactly is driving these changes.
use overlap, known as complementarity (Leduc et al., 2015; McGill, Enquist, Weiher, & Westoby, 2006). Functional groups are defined by how organisms interact with their environment through a suite of traits, ranging from physical adaptations (coral colony morphology, feeding structures) to behavioral and temporal responses such as diel vertical migration or larval stage release and duration (Cadotte,
Carscadden, & Mirotchnick, 2011). These variations in traits lead to different resource consumption patterns, decreasing functional redundancy and potentially increasing ecosystem functioning through more effective resource use and subsequent nutrient cycling and energy transfer between trophic levels (Hooper et al., 2005; Petchey, 2003). These traits are inherently linked to different life history
characteristics which are potentially easier to measure and are inherent to each species (Darling, Alvarez-Filip, Oliver, McClanahan, & Côté, 2012). By grouping species based on life history
characteristics, we can measure functional diversity and assess changes in functional groups over time, indicating overall ecosystem function and health.
In coral reef ecosystems, four different scleractinian coral functional groups have been postulated based on life history characteristics, derived from a number of biologically relevant species traits such as colony size, growth rate, and fecundity (Darling et al., 2012). These groups include competitive, tolerant, weedy, and generalist species, an expansion of the competitive – stress-tolerant – ruderal framework previously put forth by Grime et al. (Grime, 1977). For the generalist functional group, there were no species that were sampled across all periods, and therefore were not included in this study. Competitive coral species, such as Acropora spp., have extremely quick growth rates, often an order of magnitude greater than the other functional groups (Darling et al. 2012). Combined with the large colony size and branching morphologies, these corals are excellent at growing quickly, shading substrate, and outcompeting other benthic organisms for light and other resources. However, when faced with physical or physiological stress, such as storms or thermal anomalies, these corals perform much worse than other functional groups (Madin & Connolly, 2006; McClanahan et al., 2007). In addition, the white band syndrome that spread across the Caribbean in the 1970s decimated this functional group, altering the community structure of these reefs (Richard B. Aronson & Precht, 2001). These characteristics indicate that increased stress would lead to lower abundances, though the change in cover is likely to be ecologically insignificant in Belize.
On the other end of the functional group spectrum, stress-tolerant species such as certain
Diploria and Orbicella spp., have been characterized as having large, long-lived domed colonies with
slow growth rates, episodic broadcast spawning strategies, and large corallites (Darling et al. 2012). These strategies are thought to help take advantage of consistently stressful environmental conditions by allowing the corals to outlast storms through a compact domed morphology that reduces
The final functional group are weedy species, akin to the ruderal species in Grime’s C-S-R framework (Grime 1977; Darling et al. 2012). In the Caribbean, these species include the branching
Porites spp., some Agaricia spp., and some Madracis spp. along with others (Darling et al., 2012). As the name suggests, these species in general reproduce quickly, with shorter generation times recorded for species in the Indo-Pacific. However, it must be noted that Darling et al. (2012) did not show differences in generation length across functional groups in the Caribbean due to a lack of data. They are also characterized by their brooding reproductive strategy, which might help make up for their lack of fecundity when compared to the other functional groups (Darling et al. 2012), especially in heavily disturbed and low coral cover areas like Belize. By keeping their brood closer, they can settle and spread in high densities (Bak & Engel, 1979) within a localized area that fits their niche. They are also the smallest of the functional groups, on average less than a meter across, which could have implications for reef rugosity, calcification rates, and ecosystem function (Alvarez-Filip, Carricart-Ganivet, Horta-Puga, Iglesias-Prieto, & McCorkle, 2013). Some of these corals have been rising in abundance in response to declines of overall coral cover in the Caribbean (Green, Edmunds, & Carpenter, 2008), indicating that they may be dealing with increasingly stressful conditions better than competitive and stress-tolerant functional groups.
To test whether or not coral functional group abundances have shifted over time and space, we used the Belizean Barrier Reef System (BBRS) as our study location. The BBRS is the second longest barrier reef in the world, provides millions of dollars a year in economic value through fishing, tourism, and ecosystem services, and has been studied extensively since the late 1990s. It also has undergone environmental changes, such as increased sea surface temperature (Chollett, Mumby, Mü Ller-Karger, & Hu, 2012) and a number of bleaching events (Aronson, Precht, Toscano, & Koltes, 2002; McField et al., 2005). These characteristics gave us the ability to construct a unique, decadal, spatially explicit study spanning the entirety of the reef. We created statistical models testing site-specific coral functional group abundances against protection status, and local human population density to determine which functional group or groups are dominating Belize now and what might be able to explain the shifts. This information is part of an ongoing literature examining how corals have been degraded over the last several decades worldwide, and how ecosystem function might change as a result.
Methods
Study site and field surveys
Image analysis
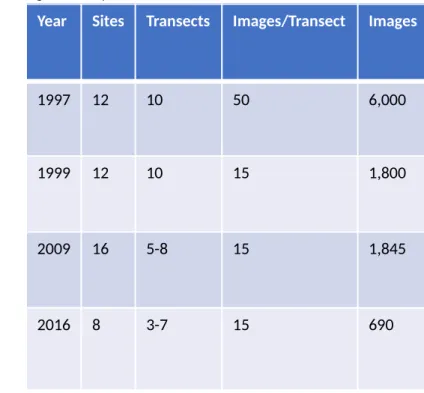
For 1997, 1999, and 2009, images were extracted from transect videos using the software package Adobe at 1 fps and subset to approximately 65-100 images per transect using Automater in MacOSX. These were then randomly subset to 50 images using R (R Core Package 2007). These final 50 images were put through quality control tests by researchers, making sure images were clear, not dark, etc. If an image did not meet standards, then it was removed from analysis. From this set of 50 images per transect, 50 were analyzed for benthic cover using Coral Point Count in 1997, 15 and 1999, while 15 images per transect were analyzed in Coral Net for the 2009 data (Table 1). For 2016, images were taken every 5 seconds during dives with a GoPro, at a swimming rate of 5-7 minutes per 30 meters, ranging between 60-90 images per transect. These images were put through similar quality control procedures, with the end dataset of 15 images per transect, and were analyzed using Coral Net. The difference in image sampling per transect was due to different research teams completing image analysis. The 1997 sampling rate of 50 images/transect was deemed oversampling and reduced to 15 images for 1999, 2009, and 2016.
Figure 1: Map of Belize sites
Year Sites Transects Images/Transect Images
1997 12 10 50 6,000
1999 12 10 15 1,800
2009 16 5-8 15 1,845
2016 8 3-7 15 690
Table 1: Surveying intensity across years
Statistical analysis
To compare the coral functional group abundances through time, we created gamma-family mixed-effect linear models with a log link in R for each separate functional group with coral cover as the dependent variable. Our covariates included fixed effects of protection status of the sites, population density within 50 km, and year, whereas site was a random effect. Protection status was gleaned from Cox et al. 2017 and the Belize Fisheries Department
(http://protectedareas.gov.bz/management_plans/). Population density within 50 km was calculated using the SEDAC gridded global population density layer, version 4 (CIESEN, 2005). Each raster layer was loaded into ArcGIS 10.3 and then subset to the 50 km buffers around each site. These cells were then averaged for a population density value for each site. The layer for the year 2000 was used to calculate population density for 1997 and 1999 years, the 2010 layer for the 2009 data, and the 2015 layer for the 2016 data.
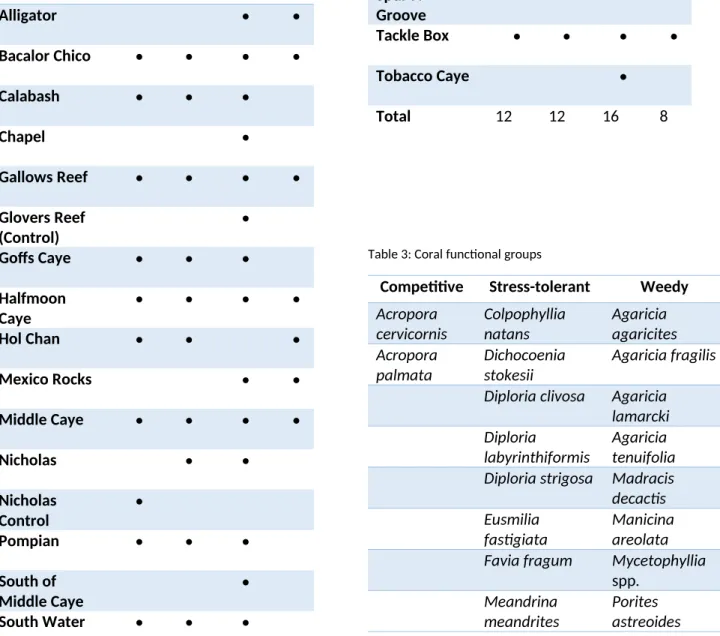
Table 2: Site survey frequency
Site 1997 1999 2009 2016
Alligator
Bacalor Chico
Calabash
Chapel
Gallows Reef
Glovers Reef (Control)
Goffs Caye
Halfmoon Caye
Hol Chan
Mexico Rocks
Middle Caye
Nicholas
Nicholas Control
Pompian
South of Middle Caye
South Water
Spur N Groove
Tackle Box
Tobacco Caye
Total 12 12 16 8
Table 3: Coral functional groups
Competitive Stress-tolerant Weedy
Acropora cervicornis Colpophyllia natans Agaricia agaricites Acropora palmata Dichocoenia stokesii Agaricia fragilis
Diploria clivosa Agaricia
lamarcki Diploria
labyrinthiformis
Agaricia tenuifolia
Diploria strigosa Madracis
decactis Eusmilia
fastigiata
Manicina areolata
Favia fragum Mycetophyllia
spp. Meandrina
meandrites
Porites
Montastraea cavernosa
Porites porites
Orbicella spp.* Siderastrea
spp.**
Stephanocoenia intersepta
* Orbicella annularis, favelota, and franksii were all combined into Orbicella spp., since the species distinction did not occur prior to the 1997 and 1999 data collection.
Results
Region-wide trends
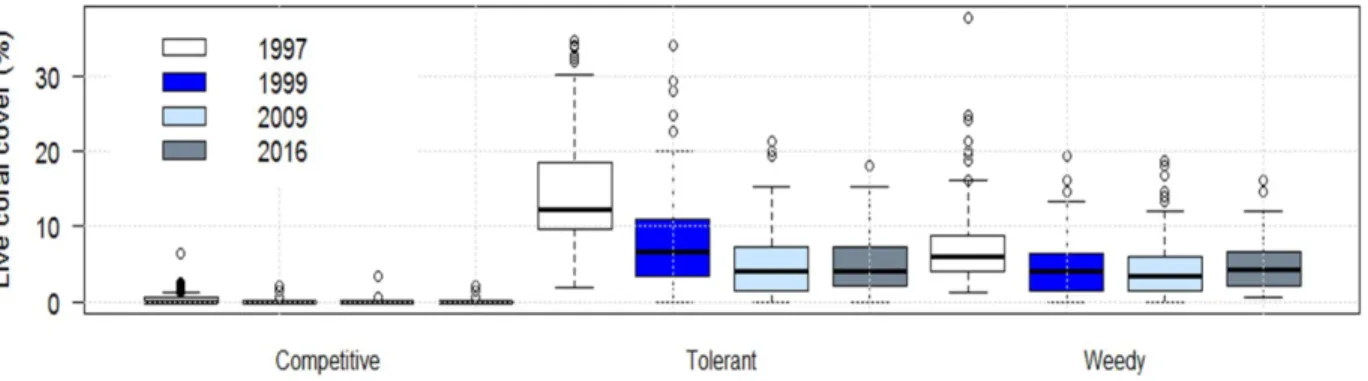
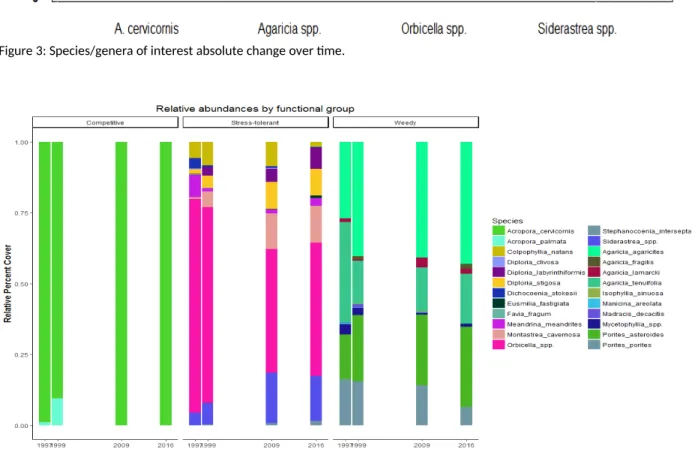
When comparing functional group total cover through time, we found that competitive corals did not significantly change (Figure 2), which was unsurprising due to their previously low cover. However, stress-tolerant corals declined between 1997 and 1999, falling to similar coverage (~5%) as weedy corals by the 2009-2016 sampling dates (Figure 2). Weedy corals decreased slightly between 1997 and 1999, and remained relatively constant thereafter. This indicates a change in the relative functional group composition, with weedy and stress-tolerant corals currently co-dominant across the BBRS, versus stress-tolerant dominance in the late 1990s (Figure 2). This change can be attributed to the loss of a key genus in the stress-tolerant group – Orbicella spp. Figure 3 shows the change in abundance of species/genera of interest, which were chosen due to their dominance within functional groups over time (Figure 4). The pattern of absolute Orbicella loss closely mirrors that of the stress-tolerant category in Figure 1 (see Figure 3), and Orbicella contributed to approximately 70% of the stress-tolerant loss of coral cover. in The importance of Orbicella loss is further backed upindicated by the failure of
Siderastrea spp. to increase in absolute abundance (Figure 2), despite increasing to second in relative
abundance within the stress-tolerant group (Figure 3). This indicates a lack of compensatory increase from other stress-tolerant corals, despite a large amount of vacated substrate. In short, we see an overall decline in coral cover across the region through time, with changes in the relative abundance of functional groups driven by the loss of Orbicella spp. and the stasis of other groups.
Figure 3: Species/genera of interest absolute change over time.
Figure 4: Relative abundance of coral species over time for each functional group.
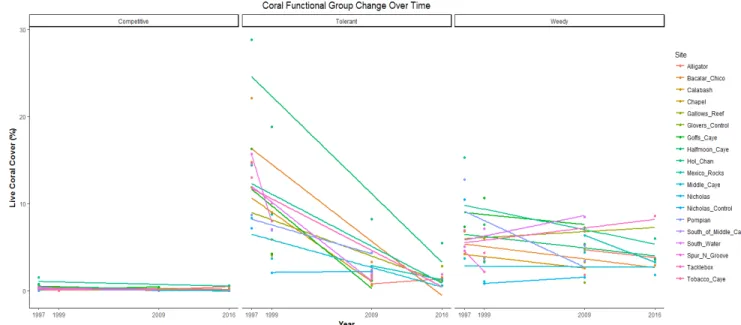
Figure 5: Coral functional group change using site level means over time. Solid lines indicate linear models with tolerant corals showing the greatest decline.
Figure 6: Human population density within 50 km correlated with live coral cover. Blue line is a linear model with an r2 = 0.009,
indicating no relationship between the variables.
Model Results
Initial model results indicate that year was the only significant covariate in determining
stress-tolerant abundances (p < 0.0001), while no covariates were significant in determining
weedy abundances. These results indicate that our models are missing some significant
covariates that could also explain the changes, such as ocean temperatures, SST anomalies, and
herbivorous fish biomass.
Discussion
and a historically significant reef building species in the Caribbean (Land & Goreau, 1970), the loss in this genus is particularly concerning, but expected due to the outbreak of Caribbean yellow-band disease and warming temperatures that help perpetuate disease transmission (Bruno et al., 2007; Harvell et al. 2009). Along with disease, declines in Orbicella spp. could have been driven by two major stressors that occurred in 1998 in Belize: a mass bleaching event and Hurricane Mitch. While the bleaching event itself had relatively little effect on mortality (5-10%) in the Caribbean as a whole (Goreau, McClanahan, Hayes, & Strong, 2000), some sites in Belize such as Glovers Reef and Channel Caye experienced up to 100% coral mortality (Aronson et al., 2000; Goreau et al., 2000), indicating the Belize might have been more affected by the bleaching. More specifically, Orbicella annularis colonies have been shown to decrease or halt their growth rate during the entirety of a bleaching event (Goreau & Macfarlane, 1990; Mendes & Woodley, 2002). Subsequent recovery of coral cover from bleaching can be repressed due to the slow growth rate of stress-tolerant corals, and when combined with declining recruits, can reducing the capacity for population growth between bleaching events. The storm which followed had no very little effect on the Orbicella spp. due to the genera’s domed morphology, but when combined with bleaching it could potentially have had a long term impact through a reduction of 20% in juvenile coral recruits (Mumby, 1999). These events were quickly followed by another outbreak of yellow band disease (Bruckner and Bruckner, 2006), which could have help further facilitate the loss of Orbicella spp. Our results agree with the prevailing literature that coral cover is declining across the Caribbean (Gardner, 2003), with Orbicella spp. in particular declining in abundance across Belize, which has several
implications for functional ecology and reef resilience in a changing climate.
An intriguing question following the loss of the dominant Orbicella is whether or not functional diversity declined with it. While we have little data to quantify this phenomenon, inferences can be made based on available data and theory. Since Orbicella performs a key function within the ecosystem as a foundation, reef-building organism, it has a much larger function to fill than any of the other stress-tolerant corals. This functionality has largely been lost on the reef with no compensatory increase from other stress-tolerant corals. Species such as Diploria spp. and Siderastraea spp. have remained at low levels, recently around 1% cover, while Agaricia is now the dominant genera across the reef (Figure 3). In order for functional diversity to work, a decline in abundance of one species from the group needs to be counterbalanced by increased abundances of other species from the group for the same level of function. Therefore, the BBRS has shown a decline in functional diversity over time, despite relatively small declines in species richness. This could be counteracted by an increase in other stress-tolerant species, though their slow growth and broadcast spawning strategies might decrease their potential of becoming quickly established in a dominant macroalgae system. Instead, another option for increased functional diversity and reef resilience is a shift towards a weedy coral dominated system, though this too has its issues.
macroalgae (Rasher & Hay, 2010), while juvenile Agaricia growth rates were decreased by 60-99% by the presence or shading by macroalgae (Box & Mumby, 2007). In addition, the colony size, subsequent rugosity, and calcification rates differ between the two functional groups extensively (Darling et al., 2012; Alvarez-Filip et al., 2013). Alvarez-Filip et al. (2013) modeled the community shift away from
Orbicella and towards weedy dominated, and they found that community calcification and reef rugosity
declined significantly over time, even with an increase in coral cover to high levels for Belize (~45%). In addition, weedy species are not nearly close enough to the abundances necessary to replace the functions Orbicella provided. All this indicates that these low lying, small colonies might not be able to act as reef-builders and replace Orbicella and other stress-tolerant species on the reef, showing that the BBRS may not be very resilient right now to future climate change.
Our study found little to no variation among sites by 2016 across all three functional groups, indicating that local stressors may not be driving patterns of coral community composition. Local impacts such as overfishing and coastal development certainly don’t aid coral reef resilience by reducing herbivory and increasing pollution (Hughes et al., 2003). However, the lack of spatial variation in coral functional groups across a range of human population densities indicates that the bigger influence on the BBRS coral community is warming ocean temperatures, disease outbreaks, and bleaching outbreaks brought on by climate change. This finding was supplemented by the model results, where human population density and local management had no effect on the functional group abundances. This study supports a broader body of literature that localized stressors on corals are not the main cause of coral cover decline (Bruno and Valdivia, 2016), and that less focus should be placed on these stressors and more so on fighting global climate change.
Works Cited
Alvarez-Filip, L., Carricart-Ganivet, J. P., Horta-Puga, G., Iglesias-Prieto, R., & McCorkle, D. C. (2013). Shifts in coral-assemblage composition do not ensure persistence of reef functionality. Scientific
Reports, 3(1), 3486. https://doi.org/10.1038/srep03486
Aronson, R. B., Precht, A. W. F., Toscano, M. A., & Koltes, K. H. (2002). The 1998 bleaching event and its aftermath on a coral reef in Belize. https://doi.org/10.1007/s00227-002-0842-5
Aronson, R. B., & Precht, W. F. (2001). White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia, 460(1/3), 25–38. https://doi.org/10.1023/A:1013103928980
Aronson, R. B., Precht, W. F., Macintyre, I. G., & Murdoch, T. J. T. (2000). Ecosystems: Coral bleach-out in Belize. Nature, 405(6782), 36–36. https://doi.org/10.1038/35011132
Bak, R. P. M., & Engel, M. S. (1979). Distribution, abundance and survival of juvenile hermatypic corals (Scleractinia) and the importance of life history strategies in the parent coral community. Marine
Biology, 54(4), 341–352. https://doi.org/10.1007/BF00395440
Box, S., & Mumby, P. (2007). Effect of macroalgal competition on growth and survival of juvenile Caribbean corals. Marine Ecology Progress Series, 342, 139–149.
https://doi.org/10.3354/meps342139
Bruckner, A., & Bruckner, R. (2006). Consequences of yellow band disease (YBD) on Montastraea annularis (species complex) populations on remote reefs off Mona Island, Puerto Rico. Diseases of
Aquatic Organisms, 69(1), 67–73. https://doi.org/10.3354/dao069067
Bruno, J. F., Selig, E. R., Casey, K. S., Page, C. A., Willis, B. L., Harvell, C. D., … Melendy, A. M. (2007). Thermal Stress and Coral Cover as Drivers of Coral Disease Outbreaks. PLoS Biology, 5(6), e124. https://doi.org/10.1371/journal.pbio.0050124
Bruno, J. F., & Valdivia, A. (2016). Coral reef degradation is not correlated with local human population density. Scientific Reports, 6, 29778. https://doi.org/10.1038/srep29778
Cadotte, M. W., Carscadden, K., & Mirotchnick, N. (2011). Beyond species: functional diversity and the maintenance of ecological processes and services. Journal of Applied Ecology, 48(5), 1079–1087. https://doi.org/10.1111/j.1365-2664.2011.02048.x
Chollett, I., Mumby, P. J., Mü Ller-Karger, F. E., & Hu, C. (2012). Physical environments of the Caribbean Sea. https://doi.org/10.4319/lo.2012.57.4.1233
Cox, C., Valdivia, A., McField, M., Castillo, K., & Bruno, J. (2017). Establishment of marine protected areas alone does not restore coral reef communities in Belize. Marine Ecology Progress Series, 563, 65– 79. https://doi.org/10.3354/meps11984
Darling, E. S., Alvarez-Filip, L., Oliver, T. A., McClanahan, T. R., & Côté, I. M. (2012). Evaluating life-history strategies of reef corals from species traits. Ecology Letters, 15(12), 1378–1386.
https://doi.org/10.1111/j.1461-0248.2012.01861.x
Gardner, T. A. (2003). Corals Long-Term Region-Wide Declines in Caribbean. Ecology Ecol. Monogr,
958(67). https://doi.org/10.1126/science.1086050
Goreau, T., McClanahan, T., Hayes, R., & Strong, A. (2000). Conservation of Coral Reefs after the 1998 Global Bleaching Event. Conservation Biology, 14(1), 5–15. https://doi.org/10.1046/j.1523-1739.2000.00011.x
Green, D., Edmunds, P., & Carpenter, R. (2008). Increasing relative abundance of Porites astreoides on Caribbean reefs mediated by an overall decline in coral cover. Marine Ecology Progress Series, 359, 1–10. https://doi.org/10.3354/meps07454
Grime, J. P. (1977). Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. The American Naturalist, 111(982), 1169–1194.
https://doi.org/10.1086/283244
Grimsditch, G. D., & Salm, R. V. (n.d.). Coral Reef Resilience and Resistance to Bleaching IUCN Global Marine Programme. Retrieved from http://www.icriforum.org/sites/default/files/2006-042.pdf Harvell, D., Altizer, S., Cattadori, I. M., Harrington, L., & Weil, E. (2009). Climate change and wildlife
diseases: When does the host matter the most? Ecology, 90(4), 912–920. https://doi.org/10.1890/08-0616.1
Hector, A., Schmid, B., Beierkuhnlein, C., Caldeira, M. C., Diemer, M., Dimitrakopoulos, P. G., … Lawton, J. H. (1999). Plant Diversity and Productivity Experiments in European Grasslands. Science,
286(5442). Retrieved from http://science.sciencemag.org/content/286/5442/1123.long
Hoegh-Guldberg, O. (1999). Climate change, coral bleaching and the future of the world’s coral reefs.
Marine and Freshwater Research, 50(8), 839. https://doi.org/10.1071/MF99078
Hoegh-Guldberg, O., Mumby, P. J., Hooten, A. J., Steneck, R. S., Greenfield, P., Gomez, E., … Hatziolos, M. E. (2007). Coral Reefs Under Rapid Climate Change and Ocean Acidification. Science, 318(5857). Retrieved from http://science.sciencemag.org/content/318/5857/1737
Hooper, D. U., Chapin, F. S., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S., … Wardle, D. A. (2005). EFFECTS OF BIODIVERSITY ON ECOSYSTEM FUNCTIONING: A CONSENSUS OF CURRENT KNOWLEDGE. Ecological Monographs, 75(1), 3–35. https://doi.org/10.1890/04-0922
Hughes, T. P., Baird, A. H., Bellwood, D. R., Card, M., Connolly, S. R., Folke, C., … Roughgarden, J. (2003). Climate Change, Human Impacts, and the Resilience of Coral Reefs. Science, 301(5635). Retrieved from http://science.sciencemag.org/content/301/5635/929
Hughes, T. P., & Tanner, J. E. (2000). RECRUITMENT FAILURE, LIFE HISTORIES, AND LONG-TERM DECLINE OF CARIBBEAN CORALS. Ecology, 81(8), 2250–2263.
https://doi.org/10.1890/0012-9658(2000)081[2250:RFLHAL]2.0.CO;2
Huntington, B. E., Karnauskas, M., & Lirman, D. (2011). Corals fail to recover at a Caribbean marine reserve despite ten years of reserve designation. Coral Reefs, 30(4), 1077–1085.
https://doi.org/10.1007/s00338-011-0809-4
Land, L. S., & Goreau, T. F. (1970). Submarine lithification of Jamaican reefs. Journal of Sedimentary
Research, 40(1). Retrieved from http://jsedres.geoscienceworld.org/content/40/1/457
Leduc, A. O. H. C., da Silva, E. M., & Rosenfeld, J. S. (2015). Effects of species vs. functional diversity: Understanding the roles of complementarity and competition on ecosystem function in a tropical stream fish assemblage. Ecological Indicators, 48, 627–635.
Madin, J. S., & Connolly, S. R. (2006). Ecological consequences of major hydrodynamic disturbances on coral reefs. Nature, 444(7118), 477–480. https://doi.org/10.1038/nature05328
McClanahan, T., Ateweberhan, M., Graham, N., Wilson, S., Sebastián, C., Guillaume, M., & Bruggemann, J. (2007). Western Indian Ocean coral communities: bleaching responses and susceptibility to extinction. Marine Ecology Progress Series, 337, 1–13. https://doi.org/10.3354/meps337001 McField, M. D., Bood, N., Fonseca, A., Arrivillaga, A., Franquesa Rinos, A., & MarÍa Loreto Viruel, R.
(2005). Status of the Mesoamerican Reef after the 2005 Coral Bleaching Event. Retrieved from https://www.coris.noaa.gov/activities/caribbean_rpt/SCRBH2005_05.pdf
McGill, B. J., Enquist, B. J., Weiher, E., & Westoby, M. (2006). Rebuilding community ecology from functional traits. Trends in Ecology & Evolution, 21(4), 178–185.
https://doi.org/10.1016/j.tree.2006.02.002
Mendes, J., & Woodley, J. (2002). Effect of the 1995-1996 bleaching event on polyp tissue depth, growth, reproduction and skeletal band formation in Montastraea annularis. Marine Ecology
Progress Series, 235, 93–102. https://doi.org/10.3354/meps235093
Miller, J., Muller, E., Rogers, C., Waara, R., Atkinson, A., Whelan, K. R. T., … Witcher, B. (2009). Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs, 28(4), 925–937. https://doi.org/10.1007/s00338-009-0531-7
Mokany, K., Ash, J., & Roxburgh, S. (2008). Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. Journal of Ecology, 96(5), 884– 893. https://doi.org/10.1111/j.1365-2745.2008.01395.x
Mumby, P. J. (1999). Bleaching and hurricane disturbances to populations of coral recruits in Belize. Retrieved from http://www.int-res.com/articles/meps/190/m190p027.pdf
Petchey, O. L. (2003). Integrating methods that investigate how complementarity influences ecosystem functioning. Oikos, 101(2), 323–330. https://doi.org/10.1034/j.1600-0706.2003.11828.x
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F., & Duarte, L. D. S. (2013). Functional redundancy and stability in plant communities. Journal of Vegetation Science, 24(5), 963–974. https://doi.org/10.1111/jvs.12047
R Development Core Team. 2007. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna (www.R-project.org)
Rasher, D. B., & Hay, M. E. (2010). Chemically rich seaweeds poison corals when not controlled by herbivores. Proceedings of the National Academy of Sciences of the United States of America,
107(21), 9683–8. https://doi.org/10.1073/pnas.0912095107
Tilman, D., Knops, J., Wedin, D., Reich, P., Ritchie, M., & Siemann, E. (1997). The Influence of Functional Diversity and Composition on Ecosystem Processes. Science, 277(5330). Retrieved from
http://science.sciencemag.org/content/277/5330/1300
van Woesik, R., Franklin, E. C., O’Leary, J., McClanahan, T. R., Klaus, J. S., & Budd, A. F. (2012). Hosts of the Plio-Pleistocene past reflect modern-day coral vulnerability. Proceedings of the Royal Society of London B: Biological Sciences, 279(1737). Retrieved from
Walker, B., Kinzig, A., & Langridge, J. (1999). Original Articles: Plant Attribute Diversity, Resilience, and Ecosystem Function: The Nature and Significance of Dominant and Minor Species. Ecosystems,