1
Short Title: Genome-wide identification of BR-regulated genes
1 2
3
Title: Comparative transcriptomic analysis to identify brassinosteroid 4
response genes 5
6
Author names and affiliations: Xiaolei Liua,1, Hongxing Yangc, Yuan Wange,
7
Zhaohai Zhua, Wei Zhanga, Jianming Li a,b,d,1
8
a
Shanghai Center for Plant Stress Biology and Center of Excellence for
9
Molecular Plant Sciences, Chinese Academy of Sciences, 201602 Shanghai,
10
China
11
b
Guangdong Key Laboratory for Innovative Development and Utilization of
12
Forest Plant Germplasm, College of Forestry and Landscape Architecture,
13
South China Agricultural University, 510642 Guangzhou, China
14
c Shanghai Key Laboratory of Plant Functional Genomics and Resources,
15
Shanghai Chenshan Plant Science Research Center, Chinese Academy of
16
Sciences, Shanghai Chenshan Botanical Garden, 201602 Shanghai, China
17
d
Department of Molecular, Cellular, and Developmental Biology, University of
18
Michigan, 830 N University, Ann Arbor, MI 48109–1048, USA
19
e
Department of Botany and Plant Science, University of California Riverside,
20
92507 Riverside, USA
21 22
1
To whom correspondence should be addressed. E-mail:
23
[email protected], [email protected]
24 25
One-sentence summary: 26
A transcriptomic database of 4498 differentially expressed genes revealed that
27
ABI4 participates in brassinosteroid response by binding to the BAK1 promoter
28
and inhibiting transcription.
29
30
Author contributions: Jianming Li conceived the research plans and Xiaolei
31
Liu conducted the experiments, Hongxing Yang analyzed the data. Wei Zhang
32
and Zhaohai Zhu assisted in the experiments. Yuan Wang assisted in
33
analyzing figures and writing the manuscript. Xiaolei Liu wrote the manuscript 34
and Jianming Li modified it. 35
36
The authors responsible for distribution of materials integral to the results
37
presented in this manuscript in accordance with the policy described in the
38
Instructions for Authors are Xiaolei Liu ([email protected]) and Jianming Li
39
([email protected] or [email protected]).
40
41
Keywords: BR, Arabidopsis, RNA-seq, ChIP-Seq, ABI4
42 43
Abstract 44
Brassinosteroids (BRs) are plant growth-promoting steroid hormones. BRs
45
affect plant growth by regulating panels of downstream genes. Much effort has
46
been made to establish BR-regulated gene expression networks, but there is
47
little overlap among published expression networks. In this study, we built an
48
optimal BR-regulated gene expression network using the model plant
49
Arabidopisis (Arabidopisis thaliana). Seven- and 24-day-old seedlings of the
50
constitutive photomorphogenesis and dwarfism (cpd) mutant and bri1-701
51
(brassinosteroid-insensitive 1-701) brl1 (BRI1-like receptor genes 1) brl3 triple
52
mutant seedlings were treated with brassinolide (BL), and RNA sequencing
53
(RNA-seq) was used to detect differentially expressed genes (DEGs). Using
54
this approach, we generated a transcriptomic database of 4498 DEGs and
55
identified 110 transcription factors that specifically respond to BR at different
56
stages. We also found that, among the identified BR-responsive transcription
57
factors, ABSCISIC ACID-INSENSlTIVE4 (ABI4), an ethylene response factor
58
(ERF) transcription factor, inhibits BR-regulated growth. Compared to wild-type
plants, theabi4-102 mutant was less sensitive to brassinazole (BRZ) and more
60
sensitive to BR. Next, we performed a chromatin immunoprecipitation followed
61
by high-throughput sequencing (ChIP-seq) assay and established that ABI4
62
binds directly to the BAK1 (BRI1-associated receptor kinase 1) promoter and
63
inhibits transcription. These results provide insight into BR-responsive gene
64
functions in regulating plant growth at different stages and may serve as a
65
basis for predicting gene function, selecting candidate genes, and improving
66
the understanding of BR regulatory pathways.
67 68
Introduction 69
Brassinosteroids (BRs) are growth-promoting steroid hormones with important
70
roles in plant development. BR biosynthesis and signal defective mutants
71
typically present round and dark green leaf, short petiole, dwarf, and male
72
sterility phenotypes. Many components of the BR signaling pathway have
73
been described. When BR is perceived by the BRASSINOSTEROID
74
INSENSITIVE1 (BRI1) protein, a transmembrane serine/threonine kinase,
75
BRI1 is activated and BR signals are transduced through the phosphorylation
76
of downstream proteins (Li and Chory 1997). BRASSINOSTEROID
77
INSENSITIVE2 (BIN2) is another negative regulator of BR response that has a
78
conserved kinase domain and C-terminal domain (Li et al. 2001, Li and Nam
79
2002). In the absence of BR, BIN2 phosphorylates transcription factors
80
BRASSINAZOLE-RESISTANT 1 (BZR1) and BRI1-EMS-SUPPRESSOR 1
81
(BES1) to inhibit their activity (Yin et al. 2002, Wang et al. 2002, He et al.
82
2002).
83
Plant hormones often act through transcription factors to regulate
84
downstream gene expression. BR regulates plant development through
85
transcription factors that either induce or repress downstream genes. Many
86
transcription factors have been identified as participating in downstream BR
87
signaling pathways. BES1 and BZR1 are two important transcription factors in
88
the BR signaling pathway. BES1 shares 88% identity with BZR1 and has
similar protein domains: a nuclear localization signal (NLS) in the N terminal, a
90
serine-rich domain in the central part, and a PEST domain in the C terminus
91
(Yin et al. 2002). BES1/BZR1 also interacts with many transcription factors,
92
such as BIM1, MYB30, MYBL2, and HAT1 to induce or reduce the expression
93
of downstream genes and integrate BR and other signaling pathways (Yin et al.
94
2005, Li et al. 2009, Ye et al. 2012, Zhang et al. 2014).
95
To fully understand how BR regulates plant growth and division through
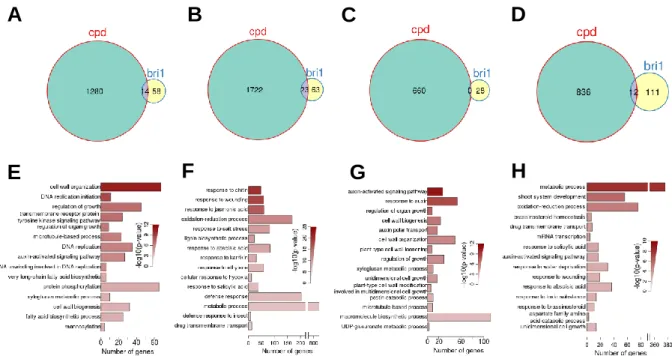
96
downstream genes, multiple research groups have identified the direct target
97
genes of BES1 and BZR1 through microarrays and chromatin
98
immunoprecipitation (ChIP) on Affymetrix tiling arrays (ChIP-chip) assays. Yu
99
used the bes1-D mutant for a ChIP-chip experiment and identified 1609
100
putative BES1 target genes (Yu et al. 2011). Sun et al. performed a ChIP-chip
101
analysis using BZR1-CFP transgene plants, and 2260 loci linked to 3410
102
genes were identified as BZR1 targets (Sun et al. 2010). However, fewer than
103
expected BR regulated genes have been identified, and we believe that there
104
are other, as yet undiscovered, transcription factors involved in
105
gene-regulating in the BR signaling pathway and more BR responsive genes
106
are needed to be identified.
107
Microarray studies using BR mutants have identified large numbers of
108
BR-responsive genes, and these data suggest that BR regulates multiple
109
cellular processes and interacts with other pathways (Mussig, Fischer and
110
Altmann 2002, Goda et al. 2004, Nemhauser, Mockler and Chory 2004, Guo et
111
al. 2009, Yu et al. 2011, Sun et al. 2010, Goda et al. 2002). Müssig performed
112
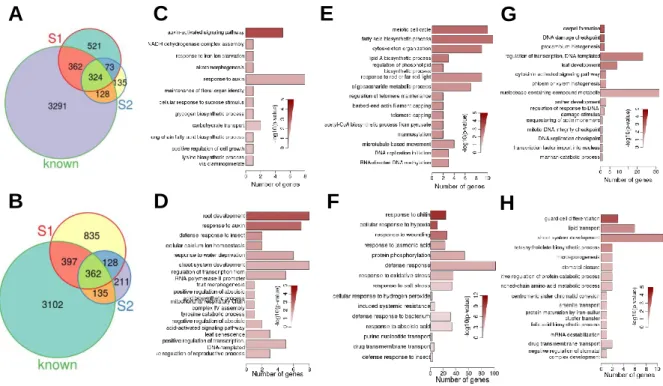
Affymetrix Arabidopsis (Arabidopsis thaliana) genome arrays using wild-type,
113
dwf1-6, and CONSTITUTIVE PHOTOMORPHOGENESIS and DWARFISM
114
(CPD)-antisense plants grown under two conditions to identify BR responsive
115
genes, identifying several BR-regulated genes (Mussig et al. 2002, Goda et al.
116
2002, 2004). Goda et al. (2002, 2004) performed microarray studies where
117
BR-deficient mutants [de-etiolated-2 (det2) and bri1-5] were treated with BL to
118
identify BL-regulated genes. Although many microarrays and ChIP-chip
analyses have aimed to identify BR responsive genes, the overlap among the
120
gene sets has been poor. This lack of overlap among gene lists is likely a
121
result of the different experimental conditions, plant tissues, and
122
developmental stages used, as well as insufficient sensitivity of the tools.
123
In this study, we have used a comparative RNA sequencing (RNA-seq)
124
approach to identify BR-responsive DEGs in Arabidopsis. For this, we made
125
use of the cpd mutant and to avoid falsely identifying non-BR induced genes,
126
we make use of the bri1-701 brl1 brl3 triple mutant as a negative control.
127
Analyses of RNA-seq data for 7- and 24-day-old seedlings identified 3002 and
128
1496 DEGs, respectively. Among the DEGs, we identified 110 genes encoding
129
transcription factors specifically respond to BR at different stage. Through
130
phenotype analysis, we found that ABI4, one of the 110 transcription factors,
131
inhibits BR-regulated growth, demonstrating that BR regulates downstream
132
genes through ABI4. By ChIP followed by high-throughput sequencing
133
(ChIP-Seq), we show that ABI4 directly binds to the BAK1 promoter and
134
inhibits BAK1 transcription. Our data support the notion that BR affects plant
135
growth by using transcription factors to regulate the expression of downstream
136
genes. These data help integrate the BR transcriptional network and guide
137
future works addressing plant responses to BRs.
138 139
Results 140
Characterization of cpd and bri1-701 brl1 brl3 triple mutants 141
cpd is a severe BR biosynthesis mutant and displays small, dwarf, dark green
142
and rounded leaf, and sterility phenotypes. Here we use the salk_023532
143
T-DNA insertion mutant of CPD (hereafter cpd), which has a BR-sensitive
144
phenotype. When cpd seeds were sown on half-strength Murashige and
145
Skoog (MS) medium with 1% sucrose, 0.8% agar, and 1 μM BL, the seedlings
146
are as sensitive as wild-type (Fig. 1A). bri1-701 is the T-DNA insertion mutant
147
of BR receptor BRI1 and is sterile. BRL1 and BRL3 are BRI1 homologs that
148
can bind BL with high affinity (Cano-Delgado et al. 2004). The bri1-701 brl1
brl3 triple null mutant showed the same phenotype with cpd and is insensitive
150
to BL (Fig. 1A). When cpd and bri1-701 brl1 brl3 were grown in soil for 24 days,
151
they also showed the same retarded phenotype (Fig. 1B).
152
BL treatments induce gene expression changes, and this offers a
153
convenient route to detecting DEGs, typically through the RNA-seq. As cpd is
154
a strong BR biosynthesis mutant, the BR induced genes were inhibited
155
significantly and the expression of BR related genes are very low. When
156
treated with BL, the gene expression will change obviously, it is convenient to
157
detect the DEGs through the RNA-seq. These RNA-seq datasets can be used
158
to investigate the molecular responses to short-term BR treatment in
159
Arabidopsis. Because of long-term BR deficiency, there might be secondary
160
effects in BR deficient mutants, and the expression of stress-related genes and
161
BR-related genes that are not transducted by BR receptor BRI1, might change
162
when treated with BR. Therefore, to avoid such genes in our list of DEGs, here
163
we use the bri1-701 brl1 brl3 triple mutant – that presents the same
164
phenotypes as cpd – as a negative control.
165 166
RNA-seq analysis of BL-treated and non-treated cpd and bri1-701 brl1
167
brl3 mutants 168
To identify DEGs during the response to BRs, an RNA-seq study was
169
performed using 7-day-old cpd and bri1-701 brl1 brl3 seedlings. At this
170
developmental stage, cells are rapidly elongating and dividing. Before
171
collecting the plant material, the seedlings were inoculated with 1 μM BL for 2
172
h. Expression analysis identified 3002 genes that were significantly
173
differentially regulated (Fig. 2A and 2B, Supplemental Table S1, S27).
174
To obtain an overview of the functional pathways in which the BR
175
responsive genes are involved, Gene Ontology (GO) enrichment analysis for
176
biological processes was performed for the RNA-seq data (Fig. 2C and 2DF).
177
The most significantly enriched GO terms in the category of biological process
178
(p < 0.05) for upregulated DEGs of the 7-day-old seedlings included cell wall
organization, DNA replication initiation, and regulation of growth (Fig. 2C). The
180
highly significant enrichment of genes involved in cell wall organization among
181
upregulated DEGs suggested a regulatory function of BR on cell growth and
182
cell division through regulating cell wall organization and DNA replication
183
initiation. In the 1722 BR downregulated genes, maximum downregulation was
184
identified for genes responding to chitin and wounding. The enrichment of
185
jasmonic acid, ethylene, and abscisic acid response genes indicates
186
antagonism between these and BR.
187
To determine whether the developmental stage of the materials
188
contributed to the discrepancies in the detection of the BR-regulated genes,
189
we also performed another RNA-seq assay using 24-day-old seedlings. At this
190
stage, vegetative tissues and cell elongation has slowed, and reproductive
191
tissue cells begin to divide and elongate. A total of 1496 genes were identified
192
with significantly altered expression in BR-treated cpd plants (Fig. 2E and 2F,
193
Supplemental Table S1, S3). We identified 660 upregulated genes and 836
194
downregulated genes. GO analysis of the genes was also performed to
195
facilitate the global analysis of BR regulated gene expression and evaluate the
196
gene functions at the 24-day-old stage. Upregulated genes were enriched with
197
genes involved in the auxin-activated signaling pathway, regulation of organ
198
growth, and cell wall biogenesis, among others. Among commonly
199
downregulated genes, metabolic process, shoot system development,
200
oxidation-reduction process, and brassinosteroid homeostasis genes were
201
significantly enriched (Fig. 2G and 2H). Taken together, these transcriptome
202
results demonstrate that BR had a significant effect on the transcription of a
203
subset of genes, through multiple mechanisms at both the 7- and 24-day-old
204
stages.
205
A total of 1940 BR upregulated genes and 2558 BR downregulated genes
206
were detected in the two examined different stage seedlings. Among these,
207
20.5% of the genes (397/1940) and 19.2% (490/2558) were commonly up- or
208
down-regulated at both developmental stages (Fig. 3A and 3B). When
comparing the 3002 DEGs of 7-day-old seedlings with 1496 DEGs from
210
24-day-old seedlings, significantly more BR responsive genes were up or
211
downregulated at the 7- than 24-day-old stage, indicating more genes respond
212
to BR at the rapid cell elongation and division stage. We also found that only
213
397 were upregulated at both developmental stages [representing 31%
214
(397/1280) of the upregulated genes in the 7-day-old seedlings list and 60%
215
(397/660) of the upregulated genes in the 24-day-old seedlings list], and that
216
490 genes were downregulated at both developmental stages [representing
217
28.5% (490/1722) of the downregulated genes in the 7-day-old seedlings list
218
and 58.6% (490/836) of the downregulated genes in the 24-day-old seedlings
219
list] (Fig. 3A and 3B). From these comparisons, we can see that the common
220
genes regulated both in 7- and 24-day-old seedlings are less than specifically
221
regulated genes in 7- and 24-day-old seedlings. This comparison and the poor
222
overlap between them suggest that BR responses at different stages might
223
have both common and distinct mechanisms.
224 225
Specific DEGs identified in 7- and 24-day-old seedlings 226
Many microarrays and ChIP-chip analyses of BR responsive genes have been
227
performed by multiple research groups using various BR mutants at different
228
development stages, collectively identifying 8102 BR-regulated genes (4105
229
BR-induced genes and 3997 BR-repressed genes) (Goda et al. 2004, Guo et
230
al. 2009, Nemhauser et al. 2004, Yu et al. 2011). Comparing our results with
231
those 8102 BR-regulated genes, we found greater overlap for the 24-day-old
232
seedlings list (68.5% upregulated and 59.4% downregulated) than for the
233
7-day-old seedlings list (53.6% upregulated and 44.1% downregulated) (Fig.
234
3A and 3B) revealed stage specification is a very important factor for the
235
discrepancies of gene expression. To identify stage-specific effects of BR, we
236
compared 729 upregulated and 1174 downregulated BR responsive genes
237
newly identified in different stages and looked for genes that were specifically
238
regulated in 7- and 24-day-old seedlings. Among them, 1356 DEGs (521
upregulated and 835 downregulated) were uniquely identified in 7-day-old
240
seedlings, and 346 DEGs (135 upregulated and 211 downregulated) were
241
uniquely identified in 24-day-old seedlings, and 201 genes were commonly
242
regulated (Supplemental Table S4 and S5). Examination of the 73 identified
243
upregulated genes shared between the two stages revealed several key
244
functional genes, including genes involved in the auxin-activated signaling
245
pathway, NADH dehydrogenase complex assembly, and response to iron ion
246
starvation (Fig. 3C). Root development, response to auxin, and defense
247
response to insect genes were downregulated both at the 7- and 24-day-old
248
stages (Fig. 3D). The meiotic cell cycle, fatty acid biosynthetic process, and
249
cytoskeleton organization genes were upregulated specifically at the 7-day-old
250
stage (Fig. 3E), suggesting the importance of the meiotic cell cycle in the BR
251
regulated tissues. Response to chitin, cellular response to hypoxia, and
252
response to wounding genes were downregulated by BR specifically at the
253
7-day-old stage (Fig. 3F). Among the genes upregulated only in the 24-day-old
254
seedlings, carpel formation, DNA damage checkpoint, and procambium
255
histogenesis genes were significantly enriched (Fig. 3G). Guard cell
256
differentiation, lipid transport, and shoot system development genes were
257
downregulated only at the 24-day-old stage (Fig. 3H). GO enrichments
258
showed it is quite different between specific up/downregulated genes in
259
7-day-old and 24-day-old seedlings. Our transcriptome results demonstrate
260
that BR-responsive programs in seedlings at different stage have many active
261
processes specifically.
262
263
Confirmation of the RNA-Seq results 264
To validate our RNA-seq data, eight genes were randomly selected for
265
RT-qPCR analysis: BR6OX2, BEH2, IBH1, KIDAR1, AT3G07010, BEE1,
266
BZR1 and BES1. From the RT-qPCR results, BL treatment significantly
267
reduced the gene expression of BR6OX2, BEH2, and IBH1 both in 7-day-old
268
and 24-day-old cpd seedlings (Fig. 4A and 4B). KIDAR1, BEE1, AT3G07010,
BZR1 and BES1 gene expression were higher after BL treatment in cpd
270
seedlings and plants (Fig. 4A and 4B). And in BL treated bri1-701 brl1 brl3
271
seedlings, the expression of these genes did not change significantly (Fig. 4A
272
and 4B). The RT-qPCR results were consistent with those of the RNA-seq.
273
These results indicate that our RNA-seq data is reliable. We also detected the
274
expression of KIDAR1 by fusing its putative promoter region to a GUS gene. In
275
the T1 transgenic plants harboring this fusion, GUS staining signals were
276
detected in rosette leaves, and the GUS signal could be significantly enhanced
277
by BL treatment (Fig. 4C).
278 279
Transcription factors that respond to BR in Arabidopsis seedlings 280
As we found the BR responsive genes identified in this study have little overlap
281
with the 1609 BES1 and 3410 BZR1 targets (Supplemental Figure S1A and B,
282
Supplemental Table S6), indicating that other transcription factors also
283
participate in BR regulating gene expression. Through further investigation of
284
our RNA-seq data, we identified 110 transcription factors among the BR
285
responsive genes, including transcription factors belonging to the ERF (n = 34),
286
bHLH (n = 14), HDZIP (n = 9), MYB (n = 26), C2H2 (n = 14), GRF (n = 4),
287
GATA (n = 4), ZFHD (n = 3), and BES1 (n = 2) families. Among them, 28 ERF
288
genes were downregulated at both the 7- and the 24-day-old stages and 6
289
ERFs and 2 BES1 homolog genes were upregulated at the 24-day-old stage.
290
Fourteen bHLH genes and 3 ZFHD genes were specifically upregulated at the
291
7-day-old stage. Twenty-six MYB genes and 14 C2H2 genes were specifically
292
downregulated at the 7-day-old stage. Nine HDZIP genes were specifically
293
downregulated at the 24-day-old stage. Four GRF genes and four GATA genes
294
were specifically upregulated at the 24-day-old stage (Table 1, Supplemental
295
Table S7). We also identify several known BR-related transcription factors that
296
are upregulated by BR, but only at defined developmental stages. BEE1 (BR
297
Enhanced Expression1), BEE2, and BEE3, which encode putative basic
298
helix-loop-helix (bHLH) proteins (Friedrichsen et al. 2002), were specifically
upregulated in the 7-day-old stage. Another bHLH transcription factor,
300
AT1G26945 (KIDAR1/PRE6), was also specifically upregulated in the
301
7-day-old stage. We also found that AIF1 was upregulated specifically at the
302
24-day-old stage and is involved in BR-regulated cell elongation and growth.
303
These results suggest that different tissues recruit different transcriptional
304
factors to respond to and transmit the BR signal at different stage. Furthermore,
305
transcription factor genes were more likely downregulated by BR treatment
306
than upregulated, and they were more likely to be downregulated in 7-day-old
307
than 24-day-old seedlings. Therefore, future work must now aim to understand
308
the functions of these transcription factors that have been newly implicated in
309
BR signaling.
310 311
abi4 is hypersensitive to BR and insensitive to BRZ 312
Different ERFs respond to BR specifically at different stage suggested an
313
important role of them and prompted us to investigate the role of ERFs in BR
314
response. For this, we ordered all the mutants of BR responsive ERFs and
315
found the abi4-102 (cs3837) mutant has a longer petiole than wild-type (which
316
is one of the typical phenotypes of plants overproducing BRs) (Fig. 5A). Next,
317
we performed BR response assays using the BR biosynthesis inhibitor
318
brassinazole (BRZ) (Asami et al., 2000). We found that abi4-102 is relatively
319
insensitive to BRZ, both in the dark and light (Fig. 5B to 5D). ABI4
320
overexpressing plants (ABI4-cFLAG) showed a dwarf phenotype and were
321
hypersensitive to BRZ both in the dark and light (Fig. 5E to 5H). In a root
322
elongation assay, abi4-102 is more sensitive to BL than wild-type, and
323
ABI4-cFLAG is almost as sensitive to BL as wild-type (Fig. 5I). To test whether
324
the phenotype of abi4-102 was correlated with the strength of BR signaling, we
325
measured the expression levels of the BR-related marker genes, including the
326
BR-suppressed genes CPD, DWF4 and a BR-induced gene Saur_AC1 by
327
RT-qPCR. Compared to the wild-type, in abi4-102, the expression of CPD and
328
DWF4 were decreased a bit, while the expression of Saur_AC1 was much
higher, and in ABI4-cFLAG, the expression of CPD and DWF4, Saur-AC1 are
330
recovered (Fig. 5J), indicating that the BR signaling outputs were increased in
331
abi4-102. These results indicate that in abi4-102, the BR signal is amplified,
332
suggesting that BR regulates gene expression through the ABI4 transcription
333
factor.
334 335
ABI4 inhibits BR-regulated plant growth by binding and inhibiting the 336
expression of BAK1
337
To test whether ABI4 represses downstream BR related genes and to identify
338
candidate target genes, we performed a ChIP-Seq assay for ABI4. For the
339
ChIP-Seq assay, a FLAG antibody was used to pull down the putative DNA
340
sequences from transgene ABI4-FLAG seedlings. ABI4-FLAG was enriched in
341
the 2-kb upstream regions of a total of 2343 genes in Arabidopsis seedlings
342
(Supplemental Figure S2, Supplemental Table S8). From the ChIP-Seq
343
analysis, we found that ABI4 binds TGGGCC motif (Fig. 6A) and ABI4 directly
344
binds BAK1 (BRI1 Associated receptor Kinase 1) promoter, and the homologs
345
of BAK1, SERK1, SERK2, while ABI5 and RD26, known targets of ABI4, were
346
used as positive controls. The binding peaks and fold enrichment values for
347
these genes are given in Table 2 and Supplemental Figure S3. We used
348
RT-qPCR to test whether the binding of ABI4 to BAK1 affects BAK1 transcript
349
levels. In the abi4-102 mutant, BAK1 transcript levels were increased relative
350
to wild-type (Fig. 6B). CHIP-qPCR assay showed that ABI4 bound to the
351
promoter regions of BAK1 (Fig. 6C). Taken together, our data indicate that BR
352
regulates plant growth through the ABI4 transcription factor, which binds to the
353
BAK1 promoter and inhibits BAK1 gene expression.
354 355
Discussion 356
In this study, we aimed to investigate the transcriptomic response of
357
Arabidopsis to BR and hoped to identify new genes and proteins involved in
358
BR-mediated regulation of plant growth. Many microarray and ChIP-IP studies
by multiple groups have investigated BR regulation, but RNA-seq analysis has
360
been relatively underutilized in the search for BR responsive genes. Also, to
361
our knowledge, very few studies searching for BR responsive genes have
362
used strong BR defective mutants, such as cpd and bri1-701 brl1 brl3, as
363
genetic tools. In this work, instead of analyzing the long-term BL treatment
364
response, we focused on identifying genes differentially expressed soon after
365
treatment, and compared lists of BR-regulated DEGs at two developmental
366
stages (7- and 24-day-old seedlings). We found that BR positively regulates
367
cell tip growth, cell proliferation, cell wall polysaccharide biosynthesis, DNA
368
methylation and auxin responses, and brassinosteroid biosynthesis. BR also
369
repressed the ethylene biosynthesis and signaling pathway, ABA signaling
370
pathway, jasmonic acid biosynthesis and signaling pathway, and fungus
371
defense responses. Among the DEGs responsive to short-term BR treatment,
372
1557 of 3002 are BR-responsive genes found in 7-day-old seedlings, and 547
373
of 1496 are found in 24-day-old seedlings. We also found that the transcript
374
levels of an AP2/ERF transcription factor encoding gene, ABI4, were
375
downregulated, and that ABI4 can bind to the promoter of BAK1, thereby
376
inhibiting its expression and plant growth. Taken together, our results
377
contribute to a more comprehensive understanding of the BR regulatory
378
network.
379
From these results, we can see that the number of DEGs identified for the
380
24-day-old plants was less than for the 7-day-old seedlings, which might be a
381
consequence of the developmental stage of these mutants in which all organs
382
are stuck in the maturation phase and the change of vegetative stage to
383
reproductive stage of plants. Differences in gene regulation patterns across
384
reproductive stages have been reported (Zhang et al. 2017). The 24-day-old
385
cpd is transforming from the vegetative stage to the reproductive stage, and
386
regulation of some genes within this context might have a stronger negative
387
impact on gene expression, so there are fewer DEGs than at the 7-day-old
388
stage. In addition, it is possible that many genes related to plant development
have already achieved maximal expression levels by 24 days, and thus the BL
390
treatment does not result in any further increase in expression.
391
BES1 is the key transcription factor of the BR signaling pathway, and has
392
six homologs: BES1, BZR1, BEH1, BEH2, BEH3, and BEH4. We found that
393
only BEH1 and BEH2 were downregulated by BR treatment (Supplemental
394
Table S7). The bes1-D and bzr1-1D mutant plants display different phenotypes
395
when grown in light because of their different biological functions in plant
396
development (Yin et al. 2002, Wang et al. 2002). The distinct functions of
397
BES1 and BZR1 indicate the different functions of these gene families. While
398
BZR1 has dual roles in regulating BR biosynthesis and growth responses (He
399
et al. 2005), BEH1–4 might also function differently. BES1 and BZR1 have
400
been well-studied, but the functions of BEH1–4 are unclear. Our results verify
401
the differences between these families and help to understand the biological
402
functions of BEH1–4. In future work, it would be interesting to compare the
403
affinities of BEH1, BEH2, BEH3, and BEH4 for gene promoters.
404
Ethylene response factor (ERF) is one of the largest transcription factor
405
families in Arabidopsis. The ERF transcription factors have an AP2/ERF-type
406
DNA-binding domain and diverse functions have been described for ERFs at
407
various developmental stages (Chandler 2018). ERF can be regulated by
408
many hormones such as ethylene, auxin, ABA (Fujimoto et al. 2000). The
409
significantly changed expression of ERF indicated the crosstalk between these
410
hormones. Recently, a stress-inducible AP2/ERF transcription factor TINY was
411
proved to inhibit BR regulated growth and positively regulates drought
412
responses (Xie et al. 2019) indicating the role of ERF in the interaction
413
between BR and ABA. Now we also found 34 ERFs (including TINY) can be
414
regulated by BR. Twenty-eight ERFs were downregulated at the 7-day-old
415
seedling stage and the 24-day-old stage specifically, and 6 ERFs were
416
upregulated in 24-day-old seedlings indicating important and specific functions
417
for ERF in BR regulating plant growth. Here we found one of the AP2/ERF
418
transcription factors, ABI4, also inhibits BR regulated growth by inhibiting
BAK1 expression. BAK1 was the co-receptor of BR signaling pathway, the
420
binding and inhibiting of BAK1 by ABI4might represent a feedback mechanism
421
for BR signaling. These results indicate the BR might recruit different ERFs to
422
bind and regulate different developmental response genes at different stage
423
and interacted with different hormones through different ERFs.
424
BR and ABA function antagonistically through multiple signaling
425
components during different development process such as seed germination
426
and root length. Many paper have been published to reveal the interaction and
427
crosstalk between BR and ABA. BR deficient and perception mutants showed
428
hypersensitivity to ABA in seed germination (Zhang, Cai and Wang 2009,
429
Steber and McCourt 2001). BIN2, negative regulator of BR signaling,
430
phosphorylates and stabilizes ABI5 to regulate ABA response during seed
431
germination (Hu and Yu 2014). BIN2 can also phosphorylate SnRK2.3, a
432
positive regulator of ABA signaling, on T180 to promote its kinase activity to
433
promote ABA signaling (Cai et al. 2014). Wang et al found that ABA inhibits BR
434
signaling by ABI1 and ABI2, which dephosphorylate BIN2 to regulate its
435
activity (Wang et al. 2018). Here, we find another transcription factor of ABA
436
signaling pathway, ABI4, also involved in BR response through binding and
437
inhibiting the expression of BAK1. ABI4 was an ABA responsive gene, the BR
438
responsiveness of ABI4 and the BR related phenotypes indicated ABI4 might
439
also participate in the crosstalk between BR and ABA. ABA could probably
440
regulate BR signaling through ABI4 regulating the gene expression of BAK1.
441
While few studies have detected the crosstalk between BAK1 and other
442
proteins, it should be a new start of future work.
443
We also identified genes involved in DNA replication initiation, cutin
444
biosynthetic process, chloride transmembrane transport were significantly
445
upregulated, whereas genes involved in response to wounding, response to
446
chitin, response to jasmonic acid, defense response to insect, response to
447
karrikin, and ethylene-activated signaling were significantly downregulated. BR
448
regulates cell elongation and cell division synergistically with another plant
hormone. Here we report that in BR-treated cpd seedlings, there are significant
450
changes in the expression of 35 genes involved in DNA replication, 60
451
wounding response genes, 48 chitin response genes, and 49 jasmonic acid
452
response genes. Although we do not know how BR regulates DNA replication
453
and response to wounding, chitin, jasmonic acid, and karrikin, we believe the
454
data presented here will contribute to future work aiming to describe the
455
mechanisms underlying these regulatory pathways.
456
Thus we identified many specific genes response to BR at different stage
457
and integrated the BR transcriptional network. From the RNA-seq data, we
458
identified 110 transcriptional factors specifically respond to BR at different
459
stage and moreover, we identified ABI4 participates the BR response and
460
showed BR hypersensitive and BRZ insensitive phenotype. ChIP-Seq assay
461
showed ABI4 directly binds BAK1 promoter and inhibit its expression. Our
462
work has emphasized the importance of investigating the functional roles of
463
BR responsive genes and the feasibility of RNA-seq to identify genes and
464
proteins involved in BR-mediated regulation of plant growth.
465 466
Materials and Methods 467
Plant Materials and Growth Conditions 468
Arabidopsis (Arabidopsis thaliana) plants cpd (salk_023532) and bri1-701 brl1
469
brl3, abi4-102 (cs3738) were used in this study. Seeds were grown on
470
half-strength Murashige and Skoog (½ × MS) medium (pH 5.8) with 0.8% (w/v)
471
agar and 1% (w/v) sucrose for 14 days at 22°C. The plants were grown at
472
22°C under a 16 h light/8 h dark photoperiod.
473
BRZ and BL treatment 474
For the BRZ (Cayman Chemical), BL (Wako Chemicals) treatment assays, the
475
plants were treated with either 1 μM BL or mock control for 2 h. Wild-type
476
seeds were grown on ½ x MS medium with 1 μM BRZ, 1 μM BL for 7 days in
477
light and 5 days in dark, followed by observation of the seedlings.
478
Total RNA was extracted from 7-day-old and 24-day-old seedlings using the
480
QIAGEN RNAprep plant kit. Oligo (dT) magnetic beads were used to enrich
481
the mRNA. mRNA was fragmented using a fragment buffer treatment. The
482
first-strand cDNA was synthesized by random hexamer-primers using the
483
mRNA fragments the template. Buffer, dNTPs, RNase H, and DNA polymerase
484
I were used to synthesize the second strand. The double-strand cDNAs,
485
purified with QiaQuick PCR extraction kit, were used for end repair and base A
486
addition. Finally, sequencing adaptors were ligated to the fragments. All
487
low-quality reads (FASTq value < 13) were removed, and 3p and 5p adapter
488
sequences were trimmed using Genome Analyzer Pipeline (Fasteris). The
489
remaining low-quality reads with ‘n’ were removed using a Python script. The
490
fragments were purified by agarose gel electrophoresis and PCR-amplified to
491
produce the sequencing library. All reads were pair-end sequenced with an
492
average insert size of 160 bp, and typical read-length of 90 bp. The
493
RNA-sequencing was performed using an Illumina HiSeq 2000 platform.
494
Differential Expression Analysis 495
The read count data for all genes and samples were imported to edgeR once
496
to perform global normalization, calculation of the common dispersion factor,
497
and then the estimation of gene-specific dispersion parameters (i.e., tag-wise
498
dispersion). Differential gene expression between each pair of BR-treated
499
plants and controls was evaluated by calling the exact test.Raw p-values were
500
adjusted to form multiple comparison effects using the q-value (false discovery
501
rate) method. The cutoff for significant differential expression was set as >1.5
502
absolute fold change (FC) and q-value < 0.01. We used the software package
503
topology-based GO scoring (topGO, version 2.26.0) of the R package to
504
conduct gene ontology enrichment analysis, with the gene-to-GO association
505
data obtained from the GO database (submitted by TAIR at 4/1/2016).
506
Gene Expression Analysis by RT-qPCR 507
Total RNA was extracted from seedlings using the QIAGEN RNAprep plant kit.
508
First-strand cDNA was synthesized using the iScriptTM gDNA Clear cDNA
Synthesis Kit (Bio-Rad). PCR primers were designed using Perlprimer soft.
510
PCRs were performed with a Bio-Rad CFX96 Real-Time System. PCR
511
reactions were performed in a total volume of 20 μl, with 1 μl of first-strand
512
cDNAs and 0.4 μl of each primer. The amplification program was: 95°C for 2
513
min, and 39 cycles of 95°C for 15 s, 55°C for the 30 s. The UBOX gene was
514
used as a control to normalize the level of total RNA. Primers for real-time
515
PCR are listed in Supplemental Table S9. The 2−ΔΔCT values from three
516
technical replicates from each biological replicate were used for the statistical
517
analysis (Schefe et al. 2006).
518
Chromatin Immunoprecipitation (ChIP) and ChIP-Seq Libraries 519
For ChIP-Seq, 12-day-old seedlings of p35S:ABI4-cFLAG plants were
520
selected for the ChIP experiment. Anti-FLAG (Abmart) was used to
521
precipiptate the DNA. ChIP assays were performed as described (Hansen et al.
522
2017), with minor modifications.
523
ChIP-Seq Assays and CHIP-qPCR 524
Chromatin immunoprecipitation (ChIP) sequencing library preparation and
525
data analysis were conducted by LC-Bio (Hangzhou, Zhejiang 310018, China).
526
FastQC (v0.11.5) was used for quality control analysis of the sequencing reads,
527
to generate an NGS quality control report. Trimmomatic (v0.36) was used to
528
clean the raw reads and filter out the adaptor and low-quality reads and
529
alignment of the reads to the genome. The reference genome for Arabidopsis
530
thaliana (TAIR10) was downloaded from
531
ftp://ftp.arabidopsis.org/home/tair/Sequences/whole_chromosomes/
532
(reference genome). For each sample, we mapped the clean sequence reads
533
to the reference genome using the STAR (v2.5.3a) program. RSeQC (v2.6)
534
was used to evaluate mapped reads distribution, coverage uniformity, and
535
strand specificity. MACS2 (v2.1.1) was used to call peaks, giving robust and
536
high-resolution ChIP-Seq peak predictions. Peaks were annotated as related
537
genes using Homer (v4.10). deepTools (v2.4.1) was used to plot gene
538
coverage of the reads near TSS and TES. ChIPseeker (v1.5.1) was used to
depict the reads distribution on chromosomes. Motif and analyze transcription
540
factors were searched by Homer (v4.10).
541
3–5 μL Immunoprecipitated products were used for ChIP-qPCR. Each
542
immunoprecipitation was performed three times independently, with the input
543
being used as the control. The primers for ChIP-qPCR are listed in
544
Supplemental Table S9.
545
Accession numbers 546
Sequence data from this article can be found in the GenBank/EMBL data
547
libraries under accession numbers: BRI1: AAC49810.1; CPD, NM120651; DWF4, 548
AF044216; SAUR-AC1, S70188.1; ABI4, AF040959.1.
549
550
Supplemental Data 551
Supplemental Figure S1. Comparison among BR-regulated genes with BES1
552
target genes, and BZR1 target genes.
553
Supplemental Figure S2. Genome-Wide Binding Profiles from ChIP-seq
554
Analysis.
555
Supplemental Figure S3. Direct Targets of ABI4.
556
Supplemental Table S1. Genes differentially expressed in cpd vs BL treated
557
cpd.
558
Supplemental Table S21. Genes differentially expressed in 7-day-old cpd vs
559
BL treated 7-day-old cpd.
560
Supplemental Table S3. Genes differentially expressed in 24-day-old cpd vs
561
BL treated 24-day-old cpd.
562
Supplemental Table S4. Genes specifically up-regulated by BR at different
563
stage.
564
Supplemental Table S5. Genes specifically down-regulated by BR at different
565
stage.
566
Supplemental Table S6. Comparison of BES1 and BZR1 target genes with
567
BR responsive genes.
568
Supplemental Table S7. Transcription factors responsive to BR.
Supplemental Table S8. Putative target genes of ABI4 in Arabidopsis
570
seedlings.
571
Supplemental Table S9. Primers used in this study.
572 573
ACKNOWLEDGMENTS 574
This work was partially supported by a grant from Natural Science Foundation
575
of China (NSFC31870253), and the research budget of Shanghai Center for
576
Plant Stress Biology to J.L.
577 578
Tables 579
580
Table 1. Transcription factors regulated by BR at different stages.
581
(Bracket means numbers of this kind of genes detected in our RNAseq result)
582
Transcription Factor
7-day-old seedlings
24-day-old seedlings
ERF(28) DOWN DOWN
ERF(6) UP
bHLH(14) UP
HDZIP(9) DOWN
MYB(26) DOWN
C2H2(14) DOWN
GRF(4) UP
GATA(4) UP
ZFHD(3) UP
BES1(2) DOWN DOWN
583
Table 2. Direct binding peaks of ABI4
584
Gene Peak Localization Fold
Change
P-Value
BAK1 Chr4:16090492-16091002 2.47047 2.17E-06
SERK1 Chr1:
27017836-27018111
2.08315 3.29E-05
SERK2 Chr1:12458754-12459029 1.99309 0.00039852
ABI5 Chr2:
15207369-15207633
2.74754 4.57E-08
RD26 Chr4:13707566-13707853 2.73776 7.32E-08
Figure Legends 586
Figure 1.cpd and bir1-701 brl1 brl3 showed the same significantly 587
retarded phenotype. 588
(A) The phenotype of 7-day-old wild-type, bri1-701 brl1 brl3, and cpd grown in
589
half-strength Murashige and Skoog medium (½ MS) and 1/2MS+1 μM
590
brassinolide (BL). Bar=1cm
591
(B) The phenotype of 24-day-old wild-type, bri1-701 brl1 brl3, and cpd grown in
592
soil.
593 594
Figure 2. BR responsiveness in brassinolide (BL)-treated 7- and 595
24-day-old cpd and bri1-701 brl1 brl3 seedlings determined by RNA-Seq 596
analysis. 597
(A, B) Venn diagrams show the numbers of genes upregulated (A) or
598
downregulated (B) in BL-treated 7-day-old cpd seedlings.
599
(C, D) The top 15 enriched Gene Ontology (GO) terms in the category of
600
biological process among genes upregulated in 7-day-old cpd seedlings (C) ,
601
downregulated in 7-day-old cpd seedlings (D). In each barplot, the x-axis
602
represents the number of regulated genes annotated to each respective GO
603
term, and color shading shows the statistical significance (negative
604
log10-transformed p-values) of the enrichment of the respective GO term
605
obtained by performing Fisher’s exact test.
606
(E, F) Venn diagrams show the numbers of genes upregulated (E) or
607
downregulated (F) in BL-treated 24-day-old cpd seedlings.
608
(G, H) The top 15 enriched Gene Ontology (GO) terms in the category of
609
biological process among genes upregulated in 24-day-old cpd seedlings (G) ,
610
and downregulated in 24-day-old cpd seedlings (H). In each barplot, the x-axis
611
represents the number of regulated genes annotated to each respective GO
612
term, and color shading shows the statistical significance (negative
613
log10-transformed p-values) of the enrichment of the respective GO term
614
obtained by performing Fisher’s exact test.
616
Figure 3. Specific differentially expressed genes (DEGs) in 617
brassinosteroid (BR)-treated cpd and bri1-701 brl1 brl3. 618
(A, B) Comparison of genes upregulated (A) or downregulated (B) in 7-day-old
619
seedlings (S1), 24-day-old seedlings (S2) of cpd, and previously identified
620
BR-responsive genes.
621
(C–H) Top 15 (or 14 for F) significantly enriched Gene Ontology (GO) terms in
622
the category of biological process for genes (C) genes upregulated or (D)
623
downregulated by BR in both 7-day-old and 24-day-old seedlings. (E)
624
Specifically upregulated or (F) specifically downregulated by BR in 7-day-old
625
seedlings, (G) genes specifically upregulated or (H) specifically downregulated
626
by BR in 24-day-old seedlings. For each barplot, the x-axis represents the
627
number of BR-regulated genes annotated to each respective GO term, with
628
color shading shows the statistical significance (negative log10-transformed
629
p-values) of the enrichment of the respective GO term obtained by performing
630
Fisher’s exact test. DSR, double-strand break; DR, defense response.
631 632
Figure 4. Validation of the RNA-seq results. 633
(A) RT-qPCR analysis of BR6OX2, BEH2, IBH1, KIDAR1, AT3G07010, BEE1,
634
BZR1 and BES1in the 7-day-old cpd and bri1-701 brl1 brl3 seedlings treated
635
with or without 1 µM brassinolide (BL) for 2 h. For each sample, the RT-qPCR
636
assays were repeated 3 times, and the error bars denote ±SD.
637
(B) RT-qPCR analysis of BR6OX2, BEH2, IBH1, KIDAR1, AT3G07010, BEE1,
638
BZR1 and BES1 in the 24-day-old cpd and bri1-701 brl1 brl3 seedlings treated
639
with or without 1 µM BL for 2 h. For each sample, the RT-qPCR assays were
640
repeated 3 times, and the error bars denote ±SD.
641
(C) Effects of brassinosteroid (BR) on the expression of pKIDAR1::GUS
642
transgene plants. The top row showed the GUS staining of wild-type (WT) and
643
10 pKIDAR1::GUS T1 plants. The bottom row showed the 1 μM BL treatment
644
increased the GUS signal of pKIDAR1::GUS transgene plants.
646
Figure 5. abi4-102 was hypersensitive to BR and hyposensitive to BRZ. 647
(A-D) Phenotype of abi4-102 grown in½ MS in light (A), grown in ½ MS in dark
648
(B), grown in ½ MS+1μM BRZ in light (C), grown in ½ MS+1μM BRZ in dark
649
(D).
650
(E-H) Phenotype of ABI4-cFLAG grown in½ MS in light (E), grown in½ MS in
651
dark (F), grown in ½ MS+1μM BRZ in light (G), grown in ½ MS+1μM BRZ in
652
dark (H).
653
(I) Quantification of root length of Col-0, abi4-102 and ABI4-cFLAG grown in
654
½ MS containing different concentration of BL.Each data point represents the
655
average of 25 seedlings of duplicated experiments, and the error bars denote
656
±SD.
657
(J) Gene expression level of CPD, DWF4 and SAUR-AC1 in treated with or
658
without 1 µM BL of Col-0, abi4-102 and ABI4-cFLAG. For each sample, the
659
RT-qPCR assays were repeated 3 times, and the error bars denote ±SD.
660 661
Figure 6. ABI4 binds BAK1 promoter and inhibit transcription. 662
(A) Sequence logo for ABI4 binding motif in the promoters of ABI4-targeting
663
genes. The height of each letter represents the frequency of the base at that
664
position.
665
(B) Gene expression level of BAK1 in abi4-102. For each sample, the
666
RT-qPCR assays were repeated 3 times, and the error bars denote ±SD.
667
(C) ChIP-qPCR results indicating that the promoter fragments of BAK1, can be
668
amplified from the immunoprecipitates pulled down by the anti-FLAG antibody.
669
For each sample, the ChIP-qPCR assays were repeated 3 times, and the error
670
bars denote ±SD.
676 677
Literature Cited 678
Cai, Z., J. Liu, H. Wang, C. Yang, Y. Chen, Y. Li, S. Pan, R. Dong, G. Tang, D.
679
Barajas-Lopez Jde, H. Fujii & X. Wang (2014) GSK3-like kinases
680
positively modulate abscisic acid signaling through phosphorylating
681
subgroup III SnRK2s in Arabidopsis. Proc Natl Acad Sci U S A, 111,
682
9651-6.
683
Cano-Delgado, A., Y. Yin, C. Yu, D. Vafeados, S. Mora-Garcia, J. C. Cheng, K.
684
H. Nam, J. Li & J. Chory (2004) BRL1 and BRL3 are novel
685
brassinosteroid receptors that function in vascular differentiation in
686
Arabidopsis. Development, 131, 5341-51.
687
Chandler, J. W. (2018) Class VIIIb APETALA2 Ethylene Response Factors in
688
Plant Development. Trends Plant Sci, 23, 151-162.
689
Friedrichsen, D. M., J. Nemhauser, T. Muramitsu, J. N. Maloof, J. Alonso, J. R.
690
Ecker, M. Furuya & J. Chory (2002) Three redundant brassinosteroid
691
early response genes encode putative bHLH transcription factors
692
required for normal growth. Genetics, 162, 1445-56.
693
Fujimoto, S. Y., M. Ohta, A. Usui, H. Shinshi & M. Ohme-Takagi (2000)
694
Arabidopsis ethylene-responsive element binding factors act as
695
transcriptional activators or repressors of GCC box-mediated gene
696
expression. Plant Cell, 12, 393-404.
697
Goda, H., S. Sawa, T. Asami, S. Fujioka, Y. Shimada & S. Yoshida (2004)
698
Comprehensive comparison of auxin-regulated and
699
brassinosteroid-regulated genes in Arabidopsis. Plant Physiol, 134,
700
1555-73.
701
Goda, H., Y. Shimada, T. Asami, S. Fujioka & S. Yoshida (2002) Microarray
702
analysis of brassinosteroid-regulated genes in Arabidopsis. Plant
703
Physiol, 130, 1319-34.
704
Guo, H., L. Li, H. Ye, X. Yu, A. Algreen & Y. Yin (2009) Three related
705
receptor-like kinases are required for optimal cell elongation in
706
Arabidopsis thaliana. Proc Natl Acad Sci U S A, 106, 7648-53.
707
Hansen, A. S., I. Pustova, C. Cattoglio, R. Tjian & X. Darzacq (2017) CTCF
708
and cohesin regulate chromatin loop stability with distinct dynamics.
709
Elife, 6.
710
He, J. X., J. M. Gendron, Y. Sun, S. S. Gampala, N. Gendron, C. Q. Sun & Z. Y.
711
Wang (2005) BZR1 is a transcriptional repressor with dual roles in
712
brassinosteroid homeostasis and growth responses. Science, 307,
713
1634-8.
714
He, J. X., J. M. Gendron, Y. Yang, J. Li & Z. Y. Wang (2002) The GSK3-like
715
kinase BIN2 phosphorylates and destabilizes BZR1, a positive regulator
716
of the brassinosteroid signaling pathway in Arabidopsis. Proc Natl Acad
717
Sci U S A, 99, 10185-90.
Hu, Y. & D. Yu (2014) BRASSINOSTEROID INSENSITIVE2 interacts with
719
ABSCISIC ACID INSENSITIVE5 to mediate the antagonism of
720
brassinosteroids to abscisic acid during seed germination in
721
Arabidopsis. Plant Cell, 26, 4394-408.
722
Li, J. & J. Chory (1997) A putative leucine-rich repeat receptor kinase involved
723
in brassinosteroid signal transduction. Cell, 90, 929-38.
724
Li, J. & K. H. Nam (2002) Regulation of brassinosteroid signaling by a
725
GSK3/SHAGGY-like kinase. Science, 295, 1299-301.
726
Li, J., K. H. Nam, D. Vafeados & J. Chory (2001) BIN2, a new
727
brassinosteroid-insensitive locus in Arabidopsis. Plant Physiol, 127,
728
14-22.
729
Li, L., X. Yu, A. Thompson, M. Guo, S. Yoshida, T. Asami, J. Chory & Y. Yin
730
(2009) Arabidopsis MYB30 is a direct target of BES1 and cooperates
731
with BES1 to regulate brassinosteroid-induced gene expression. Plant J,
732
58, 275-86.
733
Mussig, C., S. Fischer & T. Altmann (2002) Brassinosteroid-regulated gene
734
expression. Plant Physiol, 129, 1241-51.
735
Nemhauser, J. L., T. C. Mockler & J. Chory (2004) Interdependency of
736
brassinosteroid and auxin signaling in Arabidopsis. PLoS Biol, 2, E258.
737
Schefe, J. H., K. E. Lehmann, I. R. Buschmann, T. Unger & H. Funke-Kaiser
738
(2006) Quantitative real-time RT-PCR data analysis: current concepts
739
and the novel "gene expression's CT difference" formula. J Mol Med
740
(Berl), 84, 901-10.
741
Steber, C. M. & P. McCourt (2001) A role for brassinosteroids in germination in
742
Arabidopsis. Plant Physiol, 125, 763-9.
743
Sun, Y., X. Y. Fan, D. M. Cao, W. Tang, K. He, J. Y. Zhu, J. X. He, M. Y. Bai, S.
744
Zhu, E. Oh, S. Patil, T. W. Kim, H. Ji, W. H. Wong, S. Y. Rhee & Z. Y.
745
Wang (2010) Integration of brassinosteroid signal transduction with the
746
transcription network for plant growth regulation in Arabidopsis. Dev
747
Cell, 19, 765-77.
748
Wang, H., J. Tang, J. Liu, J. Hu, J. Liu, Y. Chen, Z. Cai & X. Wang (2018)
749
Abscisic Acid Signaling Inhibits Brassinosteroid Signaling through
750
Dampening the Dephosphorylation of BIN2 by ABI1 and ABI2. Mol Plant,
751
11, 315-325.
752
Wang, Z. Y., T. Nakano, J. Gendron, J. He, M. Chen, D. Vafeados, Y. Yang, S.
753
Fujioka, S. Yoshida, T. Asami & J. Chory (2002) Nuclear-localized BZR1
754
mediates brassinosteroid-induced growth and feedback suppression of
755
brassinosteroid biosynthesis. Dev Cell, 2, 505-13.
756
Xie, Z., T. Nolan, H. Jiang, B. Tang, M. Zhang, Z. Li & Y. Yin (2019) The
757
AP2/ERF Transcription Factor TINY Modulates
758
Brassinosteroid-Regulated Plant Growth and Drought Responses in
759
Arabidopsis. Plant Cell, 31, 1788-1806.
760
Ye, H., L. Li, H. Guo & Y. Yin (2012) MYBL2 is a substrate of GSK3-like kinase
761
BIN2 and acts as a corepressor of BES1 in brassinosteroid signaling
pathway in Arabidopsis. Proc Natl Acad Sci U S A, 109, 20142-7.
763
Yin, Y., D. Vafeados, Y. Tao, S. Yoshida, T. Asami & J. Chory (2005) A new
764
class of transcription factors mediates brassinosteroid-regulated gene
765
expression in Arabidopsis. Cell, 120, 249-59.
766
Yin, Y., Z. Y. Wang, S. Mora-Garcia, J. Li, S. Yoshida, T. Asami & J. Chory
767
(2002) BES1 accumulates in the nucleus in response to
768
brassinosteroids to regulate gene expression and promote stem
769
elongation. Cell, 109, 181-91.
770
Yu, X., L. Li, J. Zola, M. Aluru, H. Ye, A. Foudree, H. Guo, S. Anderson, S.
771
Aluru, P. Liu, S. Rodermel & Y. Yin (2011) A brassinosteroid
772
transcriptional network revealed by genome-wide identification of BESI
773
target genes in Arabidopsis thaliana. Plant J, 65, 634-46.
774
Zhang, D., H. Ye, H. Guo, A. Johnson, M. Zhang, H. Lin & Y. Yin (2014)
775
Transcription factor HAT1 is phosphorylated by BIN2 kinase and
776
mediates brassinosteroid repressed gene expression in Arabidopsis.
777
Plant J, 77, 59-70.
778
Zhang, S., Z. Cai & X. Wang (2009) The primary signaling outputs of
779
brassinosteroids are regulated by abscisic acid signaling. Proc Natl
780
Acad Sci U S A, 106, 4543-8.
781
Zhang, S. S., H. Yang, L. Ding, Z. T. Song, H. Ma, F. Chang & J. X. Liu (2017)
782
Tissue-Specific Transcriptomics Reveals an Important Role of the
783
Unfolded Protein Response in Maintaining Fertility upon Heat Stress in
784
Arabidopsis. Plant Cell, 29, 1007-1023.
A
B
Col-0 bri1-701 brl1 brl3 cpd
Col-0+BL bri1-701 brl1 brl3+BL cpd+BL
Col-0 bri1-701 brl1 brl3 cpd
Figure 1.cpd and bir1-701 brl1 brl3 showed same significantly retarded phenotype.
(A). The phenotype of 7-day-old wild-type, bri1-701 brl1 brl3, andcpdgrown in ½ MS and ½ MS+1 μM BL. Bar=1cm
Figure 2. BR responsiveness in BL treated 7- and 24-day-old cpd and bri1-701 brl1 brl3seedlings determined by RNA-Seq analysis.
(A) to (B) Venn diagrams show the numbers of genes upregulated (A) or downregulated (B) in BL treated 7-day-old cpdseedlings.
(C) to (D) Venn diagrams show the numbers of genes upregulated (A) or downregulated (B) in BL treated 24-day-old cpdseedlings.
(E) to (H) The top 15 enriched GO terms in the category of biological process among genes (E) upregulated in 7-day-old cpdseedlings, (F) downregulated in 7-day-old cpd seedlings, (G) upregulated in 24-day-old cpdseedlings, and (H) downregulated in 24-day-old cpdseedlings. In each barplot, the x-axis represents the number of regulated genes annotated to each respective GO term, and color shading shows the statistical significance (negative log10-transformed p-values) of the enrichment of the
respective GO term obtained by performing Fisher’s exact test.
A
B
E
G
H
C
D
A
B
C
D
E
F
G
H
Figure 3. Specific DEGs in BR treated cpdand bri1-701 brl1 brl3.
(A) and (B) Comparison of genes upregulated (A) or downregulated (B) in 7-day-old seedlings (S1), 24-day-old seedlings (S2) of cpd, and previously identified BR-responsive genes.
Figure 4. Validation of the RNA-seq results.
(A). qRT-PCR analysis of BR6OX2, BEH2, IBH1, KIDAR1,AT3G07010, BEE1, BZR1and BES1in the 7-day-old cpdand bri1-701 brl1 brl3 seedlings treated with or without 1 µM BL for 2 h. For each sample, the RT-qPCR assays were repeated 3 times, and the error bars denote
±SD.
(B). qRT-PCR analysis of BR6OX2, BEH2, IBH1, KIDAR1,AT3G07010, BEE1, BZR1and BES1in the 24-day-old cpdand bri1-701 brl1 brl3seedlings treated with or without 1 µM BL for 2 h. For each sample, the RT-qPCR assays were repeated 3 times, and the error bars denote
±SD.
(C). Effects of BR on the expression of pKIDAR1::GUS transgene plants. The top row showed
A
B
C
WT
pKIDARI::GUS T1
1 2 3 4 5 6 7 8 9 10
+BL 0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5
R
e
la
ti
v
e
G
e
n
e
Ex
p
re
s
s
io
n
L
e
v
e
l cpd cpd+BL bri1-701 brl1 brl3 bri1-701 brl1 brl3+BL 0
1 2 3 4 5 6 7
R
e
la
ti
v
e
G
e
n
e
Ex
p
re
s
s
io
n
L
e
v
e
l
Figure 5. abi4-102 was hypersensitive to BR and hyposensitive to BRZ.
(A-D). Phenotype of abi4-102 grown in½ MS in light (A), grown in ½ MS in dark (B), grown in ½ MS+1μM BRZ in light (C), grown in ½ MS+1μM BRZ in dark (D).
(E-H). Phenotype of ABI4-cFLAG grown in½ MS in light (E), grown in½ MS in dark (F), grown in ½ MS+1μM BRZ in light (G), grown in ½ MS+1μM BRZ in dark (H).
(I). Quantification of root length of Col-0, abi4-102and ABI4-cFLAGgrown in ½ MS containing different concentration of BL. Each data point represents the average of 25 seedlings of duplicated experiments, and the error bars denote ±SD.
(J). Gene expression level of CPD, DWF4and SAUR-AC1in treated with or without 1 µM BL of Col-0, abi4-102 and ABI4-cFLAG. For each sample, the RT-qPCR assays were repeated 3 times, and the error bars denote ±SD.
Col-0
MS
abi4-102
MS+1uM BRZ
Col-0 abi4-102
Col-0 ABI4-cFLAG
MS
Col-0 ABI4-cFLAG
MS+1uM BRZ Col-0 abi4-102 MS
A
C
B
Col-0 abi4-102MS+1uM BRZ
G
E
D
F
H
Col-0 ABI4-cFLAG MS Col-0 ABI4-cFLAGMS+1uM BRZ
I
J
0 0.2 0.4 0.6 0.8 1 1.20 1 10 100 1000
R a ti o o f R o o t L e n g th (% )
Concentration of BL(nM) Col-0 abi4-102 ABI4-cFLAG 0 2 4 6 8 10 12
CPD DWF4 SAUR-AC1
R e la ti v e G e n e Ex p re s s io n L e ve