PDF hosted at the Radboud Repository of the Radboud University
Nijmegen
The following full text is a publisher's version.
For additional information about this publication click this link.
http://hdl.handle.net/2066/85871
Please be advised that this information was generated on 2017-12-06 and may be subject to
change.
ISBN 9 7 8 - 9 0 - 8 8 9 1 2 3 4 , - 4
9 7 8 9 0 8 8 9 1 2 3 4 4 >
IS
O
FO
R
M
S
IN
M
U
SC
LE
AN
D
BR
AIN
CE
LL
S
lo
c
a
li
z
a
ti
o
n
an
d
fu
n
ct
io
n
•
ra
lp
h
j.
a.
ou
de
op
h
ui
s
•
20
11
DMPK ISOFORMS IN MUSCLE AND BRAIN CELLS
L O C A L I Z A T I O N A N D F U N C T I O N
Voor het bijwonen van de
openbare verdediging van het
proefschrift van
RALPH J.A. OUDE OPHUIS
DMPK ISOFORMS IN MUSCLE AND
BRAIN CELLS
LOCALIZATION AND FUNCTION
op
vrijdag 1 a p ril 2011 om 13:00u
precies in de Aula van de
Radboud U niversiteit Nijmegen
aan de Comeniuslaan 2 te
Nijmegen
Na afloop van de verdediging is
er een receptie ter plaatse
PARANIMFEN
Susan Mulders
[email protected]
Rinske van de Vorstenbosch
r.vandevorstenbosch(§) ncmls.ru.nl
DMPK ISOFORMS IN MUSCLE AND BRAIN CELLS
L O C A L I Z A T I O N A N D F U N C T I O N
ISBN-13
978-90-8891234-4
ISBN-10 90-8891-234-3
Printed by
Proefsohriftm aken.nl || Printyourthesis.com
Published by
DMPK ISOFORMS IN MUSCLE AND BRAIN CELLS
L O C A L I Z A T I O N A N D F U N C T I ON
Een wetenschappelijke proeve op het gebied van de Medische Wetenschappen
Proefschrift
ter ve rk rijg in g van de graad van doctor aan de Radboud U niversiteit Nijmegen op gezag van de rector m agnificus prof. mr. S.C.J.J. Kortmann, volgens besluit van het college van decanen in het openbaar te verdedigen op vrijdag 1 a p ril 2011 om 13:00 uur precies
door
Raphaël Johannes Antonius Oude Ophuis geboren op 24 oktober 1978
Promotor Prof. dr. B. W ieringa Copromotores Dr. D.G. W ansink Dr. J.A.M. Fransen Manuscriptcommissie Prof. dr. H.G. Brunner Prof. dr. J.A.M. Sm eitink Dr. F.J.M. van Kuppeveld
The studies presented in this thesis w ere performed at the Department of C ell Biology, Nijmegen Centre for M ole cular Life Sciences, Radboud U n ive rsity Nijmegen Medical Centre, The Netherlands. Financial support was obtained from the Prinses B ea trix Fonds, the Stichting Spieren voor Spieren, the M u scula r Dystrophy Association (M DA) and the Association Française contre les Myopathies (AFM).
TABLE OF CONTENTS
11 12 16 18 22 25 28DMPK protein isoforms are d iffe re n tia lly expressed in myogenic and neu ra l c e ll lineages
Chapter 3 49
Topology and specificity of membrane insertion of tail-anchored DMPK isoforms
Chapter 4 77
A tail-anchored myotonic dystrophy protein kinase isoform induces p e ri nuclear clustering of mitochondria, autophagy and apoptosis
Chapter 5 97
Myotonic dystrophy protein kinase isoforms A and C exert differen tia l control over Ca2+ m obilization in myotubes
Chapter 6 121
Sum m arizing Discussion
References 132 Nederlandse samenvatting 146 Abbreviations 150 Dankwoord 152 Curriculum vitae 156 Chapter 1 9 G eneral introduction
• 1 Myotonic Dystophy type 1 • 2 DM1 m olecular pathogenesis • 3 DMPK protein
• 4 DMPK splice isoforms • 5 Tail-anchored proteins • 6 DMPK substrates and function • 7 Aim and o utline of th is thesis
Chapter 2 31
i
CHAPTER 1
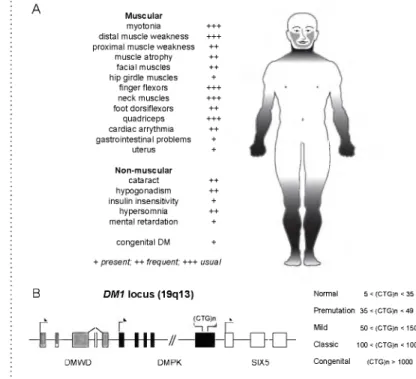
GENERAL INTRODUCTIONFigure 1 • DM1 molecular pathogenesis: symptoms and locus.
(A) C h a ra c te ristic sym ptom s of DM1. The fre q u e n c y of sym ptom s is in dic ate d by a sc a le from + to +++. Typ ical s k e le ta l m us c le fe a tu re s are illu s tra te d in the d ra w in g d a rk e s t shad in g in dic ate s m ost s e v e re ly affected s k e l e ta l m us c le areas. (B) DM1 locus org anization. The (CTG)n repeat tra c t is located in a gene-dense re gion on hum an chrom osom e 1 9 q 1 3 in exon 15 of the DMPK gene, w h ic h o ve rla p s w ith th e p ro m o te r region of the SIX5 gene. R ectan g les in d ic a te e xo n s (grey, DMWD;
black, DMPK; white, SIX5). The s tra ig h t lin e re p re s en ts in tro n s and in te rg e n ic s e quences. A rro w s denote s tart and d ire c tio n of tra n s c rip tio n . D isease c la s s ific a tio n anc c o rre sp o n d in g n u m b e r of (CTG)n repeats is indicated to the right.
GENERAL INTRODUCTION
Í
• 1. Myotonic Dystophy type 1 (DM1)DM is the most common genetic determ ined m uscula r dystrophy in a dults at a prevalence of 1:8000 (101) and was firs t described by S teinert (266), and Batten and Gibb (10) about one century ago. We now know that this m ultisystem ic neurom uscular disorder can be divided into two subtypes w ith d ifferent prevalence: DM type 1 (DM1, also known as S teinert dis ease; MIM 160900) and DM type 2 (DM2, also known as proxim al myo tonic myopathy, MIM 602668). DM symptoms va ry in s e ve rity and mostly m anifest them selves as m uscle related. Prom inent sym ptoms include myotonia, dista l and proxim al muscle weakness and atrophy, g a stro in te stin a l tra n s it problems, and cardiac a berra tions lik e conduction de fects and a rrhythm ias. Cardiac problem s are g e ne ra lly considered the leading cause of death in DM. Furtherm ore, the c e n tra l nervous system can be affected, and cataract and endocrine problem s can be involved, illu s t r ating the m ultisystem ic character of the disease (sum marized in Fig. 1A). This plethora of symptoms together w ith long asymptom atic
A
M uscular myotonia distal muscle weakness proximal muscle weakness
muscle atrophy facial muscles hip girdle muscles
finger flexors neck muscles foot dorsiflexors quadriceps cardiac arrythmia gastrointestinal problems uterus Non-m uscular cataract hypogonadism insulin insensitivity hypersomnia mental retardation +++ +++ ++ ++ ++ + +++ +++ ++ +++ ++ + + ++ ++ + ++ + congenital DM +
+ present; ++ frequent; +++ usual
DM1 locus (19q13) l L (CTG )n ^
-faotifr] i i n //—tf-E—i
-o o
N o rm a l 5 < (CTG )n < 35 P re m u ta tio n 35 < (CTG )n < 49 M ild 50 < (CTG )n < 150 C la s s ic 100 < (CTG )n < 1000 C o n g e n ita l (CTG )n > 1000 B DMWD DM PK SIX5 1 • G e n e ra l In tr o d u c ti o n1 • G e n e ra l In tr o d u c ti o n
periods and a va ria ble age of onset make DM a d iffic u lt to diagnose dis order (94). C ontrary to DM2, DM1 can also occur as a severe congenital form characterized by high neonatal m ortality, hypotonia, m ental re ta r dation and re sp ira to ry distress (101).
In the ea rly nineties of the tw en tieth century the mutation u n d e rly ing DM1 was discovered by se vera l groups on chromosome 19q13 in the 3 '-UTR of the DMPK gene (Fig. 1B) (26, 86, 175). S e vera l years later, a (CCTG)n mutation in intron 1 of the zinc finger protein 9 (ZNF9) mapping to chromosome 3q21 was identified as the cause of DM2 (167, 225).
For DM1, it has been found that se v e rity of symptoms and length of the repeat are correlated (Fig. 1B). An unaffected person ca rries a short repeat composed of 5-35 CTG triplets, w h ile m ildly affected patients ca rry 35-150 CTG units in th e ir affected allele. Patients w ith the cla ssic a l DM1 phenotype have repeats of 100-1000 CTGs and the most severely affected in divid uals c a rry a repeat of over 1000 CTGs (101, 116). Once the (CTG)n repeat crosses a threshold of 35 CTG triplets, it becomes un stable, resulting in expansion in somatic tissues (187) and in successive generations. Repeat expansion causes anticipation, meaning that se ve rity of disease increases between generations and symptoms appear e a rlie r in life (31, 102, 187).
In order to better understand the m ultisystem ic character of DM1 it is of param ount importance to gain extensive knowledge about DMPK mRNA and protein functions and expression patterns.
• 2. DM1 molecular pathogenesis
DM1 is now classified as an unstable mini- or m ic ro sa tellite or DNA expansion disorder, a heterogeneous fam ily of disorders to which also Huntington's disease, FRAXA, various types of spino cereb e lla r ataxia (94) and s e ve ra l other neurodegenerative disorders belong. Upon discovery that the mutation leading to DM1 is localized in the 3'-UTR of the DMPK gene, it was realized that special mechanisms of pathogenesis must be operational because form ation of abnorm al protein stru c tu re cannot be involved. Hence, se ve ra l hypotheses were postulated to explain the mul- tisystem ic nature of DM1 pathology.
• 2.1. RNA pathogenesis
The most p revalent and best supported explanation for the m olecular pathology of DM1 is an RNA-based toxic gain-of-function mechanism, in which newly formed tran scrip tio n products containing large (CUG)n repeats are trapped in the nucleus, w here they aggregate w ith nuclear proteins to form ribonuclear protein complexes of abnorm al nature (75, 111, 129, 181). The end re s u lt is global m isregulation of gene exp res sion (43, 65, 225). J u s t lately, it was speculated that soluble tra n scrip ts containing long CUG-repats are also able to bind tran scrip tio n and splice factors, in this way contributing to gene m isregulation (132). Length of the repeat is correlated w ith binding of n uclear proteins, nuclear reten tion and form ation of n uclear aggregates (54, 58). A n a lysis of ribonuclear foci in muscle and brain tissue from DM1 patients revealed presence of splice factors of the m uscleblind fam ily (MBNL 1, 2 and 3) w ith in the
ribonucleoprotein (RNP) aggregates. It is therefore commonly believed that the amount of MBNL a va ila b le for proper regulation of splicing is reduced. Although not found in nuclear foci but believed to bind soluble, expanded transcripts, other RNA processing factors able to bind (CUG) n repeats are upregulated or are behaving abnormally, like CUG-binding protein 1 (CUG-BP1) a member of the CUG-BP and ETR3-like (CELF) fa m ilies of factors (132, 278) or HnRNP-H (139). Furtherm ore, se vera l other proteins not involved in splicing such as protein kinase R (276), tran scrip tio n factor SP1 (specific protein 1), signal tran sd uce rs and ac tiva to rs of tran scrip tio n (STAT1 and 3), the retinoic acid receptor gamma subunit (58, 65) and Nkx2.5 (306) may also be involved in DM1 via an RNA-mediated mechanism.
In DM1 tissues, m ultiple splice a bnorm alities have been described (58), including in muscle: splicing of the mRNAs for chloride channel 1 (ClC-1 ) (40, 180), sarcoplasm ic/endoplasm ic reticulum Ca2+ ATPase 2 (143), cardiac troponin T (110, 219), insulin receptor (IR) (248), and the ryanodine receptor (143). In brain, processing of the mRNAs for Tau (164, 253), and amyloid precursor protein (129) is m isregulated. Two of these a berra nt processing events have been associated w ith typ ica l muscle symptoms: myotonia and insulin resistance are associated w ith mis- splicing of the ClC-1 and the IR respectively (58). A lte ra tio ns in splicing m ostly involve a sh ift in expression from an adult to an em bryonic mode of splicing, w ith production of isoform products typ ic a l for the imm ature state. Both reduction of a va ila ble MBNL and increase in CUG-BP1 expres sion support this sh ift in splice mode, since MBNL induces adult-type splicing w hereas CUG-BP1 is known to promote em bryonic splice modes (58).
• 2.2. Sm all noncoding RNAs
In recent years it has become cle ar that the non-coding DNA sequences in the genome are not ju s t noncoding or "stu ff er” DNA, but that they contain essentia l sequences encoding e.g. microRNAs (309), w hich have been found to be im portant in the regulation of em bryogenesis (308), protection against c e ll stress and pathogenic in sult and in gene re g u la tion in general (309).
In a paper by M alinina it was hypothesized that the hairpin struc tu re of the CTG-repeat in the m utant DMPK gene transcript, when processed by the RNAi machinery, could give ris e to CUG containing sm a ll noncod ing RNAs (178). In turn, these sm all noncoding RNAs could serve in breakdown or blocking of tran sla tio n of CUG- or CAG-containing mRNAs. Although the CUG containing mRNAs are not fu lly com plem entary to the CUG containing sm a ll noncoding RNAs, each U-U mismatch is flanked at each side by two C-G pairs providing sufficient stab ilization for the formation of near com plem entary double stranded RNA molecules. P re dicted candidates that could be affected are MBNL1 (CUG-repeat) or the myosin phosphatase-Rho interacting protein (CAG-repeat), w here the silencing effect of the CUG containing sm a ll noncoding RNAs would be most effective against the CAG sequence (178). Concrete evidence to support that this hypothetical mechanism is in fact active in DM1 was provided by Krol et al. (153) who showed that tra n scrip ts containing long CUG repeats, like the DMPK transcript, can be a targ et for Dicer. The
1 • G e n e ra l In tr o d u c ti o n 13
1 • G e n e ra l In tr o d u c ti o n
products generated by Dicer are sh o rt CUG repeats that act as siRNAs which are used in the RNA interference pathway to dow nregulate repeat containing mRNA. These re su lts provide evidence that sm a ll noncoding RNAs could play a role in the development of DM1 pathogenesis.
• 2.3. Neighboring gene effects
The (CTG)n repeat in the DMPK gene is located w ith in a CpG island, which extends to over 3.5 kb (22). When the CpG island is interrupted by an expanded repeat this increases nucleosome binding across the area, re sulting in a more condensed chrom atin stru c tu re (22, 83, 147, 152). This observation, together w ith the finding of a disruption of a DNase hyper sen sitive site located ju s t downstream of the repeat led to the hypothesis that repeat expansion may be involved in a ltering tran scrip tio n of DMPK and neighboring genes DMWD and SIX5 (77, 125, 147, 153, 247) (Fig. 1B). Products of the DMWD gene (form erly known as DMR-N9 in mouse or gene 59 in human), located ju s t upstream of DMPK, have been found in brain (127, 299) and testis (127), tissues implicated in DM1 pathology. Com parative expression profiling of DMWD in DM1 patients and unaf fected individuals, however, did not re ve a l a consistent reduction in ex pression in the DM1 patient group. Furtherm ore, no co rrelation between repeat length and DMWD expression le ve ls has been found, dismissing DMWD's candidacy for involvem ent in DM1 disease etiology (4, 71, 83).
The gene encoding homeodomain tran scrip tio n factor SIX5 is located ju s t downstream of DMPK. In fact, the (CTG)n repeat overlaps w ith the SIX5 prom otor/enhancer region (103, 147). Its expression in eye, heart and s k e le ta l muscle and testis, a ll tissues affected in DM1 pathology, im plicates SIX5 as an im portant candidate gene for involvem ent in DM1 (107, 213, 245). This idea has been fu rth e r strengthened by the finding that mRNA expression of SIX5 in DM1 patients is reduced by about 50% (147, 275). To study the role of SIX5 in DM1 pathology in more detail Six5- deficient mice have been generated (146, 243). Homozygous knockout mice and, interestingly, also mice that lack only one Six5 allele, devel oped cataract at an ea rly age (146, 245, 243). Together, these findings seem ingly indicate that a decrease in SIX5 expression may be involved in cataract formation in DM1. Since the cataracts observed in Six5- deficient mice w ere not of the same type as found in DM1 patients (224), however, there is s t ill doubt about the contribution of SIX5-deficiency to the ocular pathology in DM1. Moreover, the fact that also patients with DM2 develop cataracts on the basis of a com pletely unrelated genome mutation argues against sim ple causal involvem ent of abnorm al SIX5 ex pression (224). Finally, Six5 KO mice showed also impaired male fe rtility, suggesting that SIX5 plays a role in sperm atogenesis (144, 245), which could be linked to hypogonadism in DM1 patients. Much of this w ork is s t ill in the speculative phase, however, and the true role of neighboring genes of DMPK in DM1 disease pathology is therefore s t ill under debate.
• 2.4. Aberrant expression of DMPK protein
Ever since the discovery of the DMPK gene mutation, studies were un dertaken to identify DMPK expression levels in patient tissues. U ntil now, co ntroversy on this topic remains. W here some studies report increased DMPK expression, others show a decrease (35, 73, 85, 176, 206, 298).
Figure 2 • DMPK and NDR fam ilies of serine/threonine protein kinases.
Ph ylo g e n e tic tre e of the DMPK (DMPK, MRCKa, -ß and -y, ROCK-I and II, and CRIK) and NDR fa m ilie s (NDR1 and -2 and L a ts l and -2) of protein kinases. The s tru c tu re of the tre e is basec on h om ology com parison across the s e rin e /th re o n in e protein kin a se dom ain, using C lu sta lW (1 84, 274). Mouse seq ue n c e s w e re used and pe rcen tag e s eq uen ce id e n tity re la tiv e to the DMPK kinase dom ain is given. The tree is d ra w n to scale. Adapted from W a n s in k et al. (297).
Reduced expression is consistent w ith a haploinsufficiency model, where mRNA derived from the DM1 a lle le is retained in the nucleus in nuclear foci and unavailable for tran sla tio n into DMPK protein products (54, 182, 272).
To investigate the effect of ha ploinsufficiency and the function of DMPK, DMPK KO mouse lines were generated, independently by two labs, including ours (126, 227). Replacement of the firs t seven exons of the DMPK gene by a neomycin resistance cassette resulted in complete ab sence of DMPK protein (298). KO mice were via ble and only showed mild myopathy in head and neck m uscles at an advanced age (126). Skeletal muscle cells derived from these mice demonstrated impaired calcium hom eostasis (13). The DMPK KO mouse generated by Reddy et al. via a sim ila r strategy displayed a late onset progressive sk e le ta l myopathy, characterized by a decrease in force generation and increased fiber degeneration (227). Upon closer inspection, these mice w ere shown to display cardiac conduction defects including first, second and third de gree a trio v e n tric u la r block. The development of third degree he art block closely resem bles the condition found in DM1 patients, suggesting a potential ro le for DMPK haploinsufficiency in DM1 etiology (16). F u rth e r more, DMPK KO mice showed increased insulin resistance (170) resulting in a sig nifican tly higher body weight when they w ere fed a high-fat diet (169). Loss of DMPK expression might therefore be involved in the de vel opment of type II diabetes in DM1 patients (170).
To examine c e ll biological and physiological consequences of DMPK overexpression, a transgenic overexpressor mouse model (Tg26) has been generated. Approxim ately 25 copies of the entire human DMPK gene including reg ula to ry flanking segments w ere tandem ly integrated into the genome of this model. This resulted in a continuous surplus of DMPK transcripts, resu lting in 5-10 fold excess of human DMPK pro tein over endogenous mouse DMPK protein le v e ls (126). Tg26 transgenic mice demonstrated hypertrophic cardiomyopathy, rem iniscent of human cardiac hypertrophy, and neonatal m o rta lity indicating a d e trim enta l ef fect of DMPK overexpression during pregnancy (126). A dditional studies revealed cardiac dysrhythm ia, myopathy in s k e le ta l muscle and hypoten sion of smooth muscle suggesting a role for DMPK in a dose- or activity- dependent m anner safeguarding the physiological functioning of sk eleta l muscle (211). Since DMPK was found to be involved in myogenesis, hav ing eithe r too much or too little of this protein could induce aberrant
% ---DMPK...(100) MRCKa... (71) — MRCKß... (69) — MRCKy...(70) --- ROCK I...(52) --- ROCK II... (51) --- CRIK... (47) --- NDR1...(46) --- NDR2... (45) --- LATS1.... (42) --- LATS2...(43) 15 1 • G e n e ra l In tr o d u c ti o n
1 • G e n e ra l In tr o d u c ti o n
m u s c l e m a t u r a t i o n (36). Tak en toge ther , s e v e r a l s y m p t o m s found in DM1 p a t ie n t s c o u ld s o m e h o w be c o r r e l a t e d to a b e r r a n t l e v e l s of D M P K p r o tein. On the o t h e r hand, the f u l l c o m p l e x i t y of th e m u l t i s y s t e m i c f e a t u r e s o b s e r v e d in DM1 p a t ie n t s c a n c e r t a i n l y not be e x p l a i n e d by e f f e c t s at the prot ein l e v e l only.
• 3. D M P K protein
O v e r t im e it h a s b e c o m e c l e a r t h a t the m u l t i s y s t e m i c n a t u r e of DM1 p a t h o l o g y is not e a s i l y e x p l a i n e d by j u s t one m e c h a n i s t i c m o d e l. It r e m a i n s i m p o r t a n t to i n v e s t i g a t e p r o p e r t i e s of prot ein an d R N A p r o d u c t s of the D M P K gene si d e by side.
• 3.1. D M P K fa m i l y m e m be rs
D M P K , an e v o l u t i o n a r y you ng prot ein only found in m a m m a l s , is c l a s sifi ed a s a s e r i n e / t h r e o n i n e protein k i n a s e b el o ng in g to the group of AGC ( c A M P - d e p e n d e n t prot ein k i n a s e / p r o t e in k i n a s e G /pr ot ein k i n a s e C) k i n a s e s (183). W i t h i n the group of AG C k i n a s e s , D M P K t o g e t h e r w i t h its c l o s e s t h o m o l o g s , m y o t o n ic d y s t r o p h y k i n a s e - r e l a t e d Cd c4 2 - b in d i n g k i n a s e ( M R C K ) a , ß an d y (166, 208), r h o - a s s o c i a t e d c o i l e d - c o i l co n t a in i n g k i n a s e ( R O C K ) I an d II (2 3 2 ) an d c it ro n k i n a s e ( C R I K ) ( 1 7 3 ) m a k e up the D M P K s u b f a m il y , for w h i c h D M P K i t s e lf is the a r c h e t y p e m e m b e r (Fi g . 2) (184). D M P K f a m i l y m e m b e r s s h a r e a c o m p a r a b l e s t r u c t u r a l o r g an iz at io n w i t h a l e u c i n e - r i c h N - t e r m i n u s an d a s e r i n e / t h r e o n i n e k i n a s e d o m a in f o l l o w e d by a c o i l e d - c o i l re g io n of v a r i a b l e l e ng t h (2 32 , 313). A l t h o u g h s i m i l a r in d o m a in o rg an iz at io n , D M P K h o m o l o g s d if fe r c o n s i d e r a b l y in length. The m a i n d i f f e r e n c e s a r e the v a r i a b l e c o i l e d - c o i l d o m a in and p r e s e n c e of a d d i t i o n a l d o m a i n s lik e a G T P a s e - b i n d i n g d o m a in , ple x tr in h o m o l o g y d o m a in , c y s t e i n e - r i c h d o m a in an d c it ro n h o m o l o g y d o m a in (2 9 7 ). M o re d i s t a n t r e l a t i v e s of D M P K a r e m e m b e r s of the NDR s u b f a m i l y of k i n a s e s NDR ( n u c l e a r D b f 2 - r e l a t e d ) 1 and 2 and L A T S ( l a r g e t u m o r s u p p r e s s o r ) 1 an d 2 (F ig . 2) (33, 183). NDR1 and 2 r e s e m b l e D M P K both in d o m a in s t r u c t u r e an d le ngth. LATS1 and 2, on the o t h e r hand, ar e c h a r a c t e r i z e d by an N - t e r m i n a l u b i q u i t i n - a s s o c i a t e d d om a in , c o n n e c t e d to the s e r i n e / t h e r o n i n e k i n a s e d o m a in v i a a long lin k e r (33, 183).
M U S C L E N O N - M U S C U L A R s k e l e t a l m u s c l e t e s t is d i a p h r a g m e y e s to ng ue brain h e a r t e s o p h a g u s s t o m a c h i nt e st in e b la d d e r Table 1 • DMPK tissue distribution. L is tin g of t is s u e s or o rg a n s th a t s h o w e x p r e s s io n of D M PK. O b ta in e d fro m s t u d ie s in m ic e a n d h u m an s , a d a p te d fro m W a n s in k et al. (2 9 7 ).
Figure 3 • The DMPK gene gives rise to different, altern atively spliced, serine/ threonine protein kinases.
(A ) T he D M P K g e n e c o n s is t s of 15 e x o n s a n d is s tro n g ly c o n s e r v e d b e tw e e n m a n and m o u s e (h u m a n D M P K gen e, d ra w n to s c a le ). R e c t a n g le s in d ic a t e exo ns; th e s tra ig h t lin e r e p re s e n t s in tro ns. E x o n ic p a rts t h a t a re s u b je c t to a lt e r n a t iv e s p lic in g -i.e., a lt e r n a t iv e u s e of 5' a n d 3' s p lic e s ite s - in ex o n s 8 and 14 a re in d ic a te d in b la c k . ( B ) D e ta ile d illu s tr a t io n of a lt e r n a t iv e s p lic e m o d e s ir th e D M P K g en e. In c lu s io n of 15 n u c le o t id e s in exo n 8 r e s u lt s in p re s e n c e of the V S G G G m o tif (le f t p a n e l). A lt e r n a t iv e u s e of fou r n u c le o t id e s in exon 14 is r e s p o n s ib le fo r tw o d iffe re n t o p en re a d in g f r a m e s (O R F ), d e fin in g C - te r m in a l t a il 1 a n d t a il 2 (m id d le p a n e l). A s t e r is k s in d ic a t e stop c o d o n s in th e O R Fs. S k ip p in g of e x o n s 13 a n d 14 is a s m o o t h - m u s c le s p e c ific e v e n t, w h ic h r e s u lts in a stop c o d o n at th e b e g in n in g of exo n 15 and e x p re s s io n of a s h o rt tw o - a m in o - a c id C - te rm in u s (t a il 3 ) (r ig h t p a n e l). (C ) D o m a in o r g a n iz a tio n of th e six m a jo r D M PK is o fo rm s A th ro u g h F (d r a w n to s c a le ). A ll is o fo rm s h a ve a le u c in e - r ic h N - te rm in u s f o llo w e d by a s e r in e / t h re o n in e p ro te in k in a s e d o m a in , in c lu d in g an e x ten s io n to th e k in a s e d o m a in , an a - h e lic a l c o ile d - c o il d o m a in a n d d iffe re n t C - te rm in a l t a ils . A lt e r n a t iv e s p lic e m o d e s s h o w n u n d e r ( B ) d e fin e th e p r e s e n c e or a b s e n c e of th e V SG G G m o tif a n d the n a tu re of th e C - te r m in a l tail. A d a p te d fro m W a n s in k et al. (2 9 7).
A
ATGi>
2 3 4 5 6 7 8 (CTG)nI
10 11121314 15<
with VSGGG motif w/o VSGGG motif 12 V 13 14K Î H Î Ï
>
1213 14 15C
extension Leu-rich to kinase dom ain S er/Th r kinase dom ain domainVSGGG m o tif
coiled C-term inal co il tail
DMPK B
DMPK C
DMPK D
DMPK E
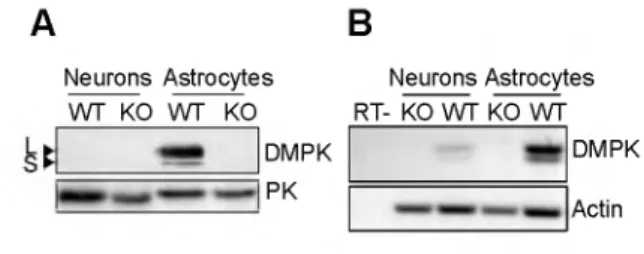
• 3.2. Cell type and t i sc u e distrib ution of D M P K exp ression
D M P K pr ote in is r x p r e s s ed ir a wid e r a n g e of t i s s ues, w i t h hi gd est e x p r e s s ¡0 n in s m o o th m u s o le, S k e s t o m a c h an d hl ad de r (Fi g . 3) (95,
2 11, 114 4). In o t h e r m u s c l e ty pe s like hea rt, s k e l e t a l m u s c l e, to ng ue and d i a p h r a g m , D M P K is a l s o found ( T a b le 1) (95, 126, 174, 244). F u r t h e r s p e c i f i c a t i o n ot the e x p t e s s i o n p n t t e r t in sl<eldtal m u s c l e s h o w A td a t D M P K is p r e s e n t ie both s l o w a n t feist t w i t c h m u s c l e f i b e r s (64, 73, 244). In ti so u e s lik e t e s tis a nd tir ain D M P 4 h a s a l s 0 been f o und, a lb e .t in m in o r a m o u n t s (211, 244). A l t h o u g h D M P K p ro t e in w a s fo und in n e u r o n s , the a n t i b o d i e s u s e d in th is s t u d y re co g n i z e d a 50 kDa prot ein k n o w n to be a c r o s s r e a c t i n g prot ei n ( s e e b e l o w ) (8, 68). Us ing in situ hyb rid iz a tio n S a r k a r an d c o w o r k e r s d e t e r m i n e d D M P K m R N A in n e u r o n s in the c e r e b r a l cor te x, a lt h o u g h th e d at a p r e s e n t e d only s h o w m i n o r e x p r e s s i o n j us t a b o v e b a c k g r o u n d l e v e l (244). S o the q u e st i o n w h e t h e r D M P K e x p r e s s i o n in b rai n is co n fi n e d to n e u r o n s or a l s o o c c u r s in g l i a l c e l l s , h a s s t i l l not been a d e q u a t e l y a n s w e r e d and f u r t h e r s t u d y into th i s c o n t r o v e r s i a l i ss u e is n e e d e d . D M P K m R N A h a s a l s o been ide n tif ie d in l i v e r (2 4 4 ), a l t h o u g h no prot ei n e x p r e s s i o n w a s found using W e s t e r n blot (95). A l s o for le ns s o m e s t u d i e s s h o w p r e s e n c e of D M P K protein an d m R N A (63, 244), w h i le o t h e r s f a i l e d to d e t e c t D M P K m R N A (3 0 3 ) . M u l t i p le g ro up s h a v e tri ed to r a i s e a n t i b o d i e s a g a i n s t v a r i o u s D M P K e p i t o p e s , but p r o t e i n s i de n ti fi e d ha d s e e m i n g l y d if f e r e n t m o l e c u l a r 17 9 tail 1 tail 3 tail 2 DMPK A tail 1 tail 2 DMPK F 1 • G e n e ra l In tr o d u c ti o n
1 • G e n e ra l In tr o d u c ti o n w e i g h t s , ra n g in g fro m 42 to 84 kD a (159, 218, 283, 285). C r o s s - r e a c t i o n to D M P K - r e l a t e d p ro t e i n s e x p l a i n e d t h e s e fi ndings. P r o d u c t s of th e D M P K gene w e r e p r e d i c t e d to f a l l in th e 6 5 - 8 0 kDa size r an g e , so the 42-55 kDa p ro t e i n s r e c o g n i z e d by the f i r s t - g e n e r a t i o n a n t ib o d i e s so o n w e r e no l o n g e r c o n s i d e r e d tr u e p r o d u c t s (95, 159, 218). R e a c t i v i t y t o w a r d s D M P K h o m o l o g u e s like M R C K a an d ß ( 1 5 9 ) m a d e the p ro d u c t io n of s p e c i f i c h i g h - a f f i n it y a n t i b o d i e s a g a i n s t D M P K a t e d i o u s and d if f i c u l t job. Not u n t i l r e c e n t ly , thi s si t u a t i o n h a s been i m p r o v e d w i t h th e d e v e l o p m e n t of m o n o c l o n a l a n t i b o d i e s w i t h t ru e D M P K s p e c i f i c i t y (159, 218).
B e c a u s e m a n y e a r l y s t u d i e s m a d e u s e of the f i r s t - g e n e r a t i o n a n t i bod ies, co n fu s i o n r e m a i n s a b o u t th e s u b c e l l u l a r l o c a l i z a t i o n of D M P K, an d c a r e sh o u l d be t a k e n in in t e rp re t i n g i m m u n e - h i s t o c h e m i c a l fi n d ings in e a r l y re p o rt s . S e v e r a l g r o u p s (11, 174, 301) found D M P K in the n e u r o m u s c u l a r j u n c t io n of s k e l e t a l m u s c l e , n e a r th e a c e t y l c h o l i n e r e ce p t o r i m p l i c a t i n g a ro l e in s i g n a l t r a n s d u c t i o n . H o w e ve r, using a p a n e l of m o n o c l o n a l a n t i b o d i e s a g a i n s t D M P K no e x p r e s s i o n of the prot ein at the n e u r o m u s c u l a r j u n c t io n w a s found by P h a m et al. (218). D M P K w a s d e t e c t e d at th e s a r c o p l a s m i c r e t i c u l u m (64, 240, 256), n e a r th e S e r C a A T P a s e and in the T - t u b u le s of ty pe I s k e l e t a l m u s c l e f i b r e s (2 8 3 ). In c a r - d ia c m u s c l e , D M P K w a s found in th e s a r c o p l a s m i c r e t i c u l u m (205, 284), next to the i n t e r c a l a t e d d i s c s (174, 218, 240, 3 01 ) w h e r e D M P K w a s a s s o c i a t e d to gap j u n c t i o n s (2 05 , 283). In o t h e r c e l l t y p e s like a d u l t s p i n a l m o t o r n e u r o n s in bra in, D M P K l o c a l i z e d in t h e c y t o s o l (6 8 ) , a t th e E R or n e a r d e n d r i t i c m i c r o t u b u l e s (8). O v e r a l l it h a s bee n d if f i c u l t to in t e rp re t t h e s e r e s u l t s , a l s o b e c a u s e h y d r o p h o b ic i t y p re d i c t i o n c l a s s i f i e d D M P K as a m e m b r a n e bound (64, 205, 240, 256, 2 84 ) or c y t o s o l i c (68, 218, 284) protein. N o w a d a y s , the v a r i a t i o n in l o c a l i z a t i o n d e s c r i b e d for D M P K ca n be b es t e x p l a i n e d by the e x i s t e n c e of m u l t i p l e s p l i c e i s o f o r m s a s w i l l be d i s c u s s e d b e l o w (95, 286, 296). I n d e p e n d e n t e v i d e n c e for v a r i a b l e s u b c e l l u l a r l o c a l i z a t i o n of D M P K c a m e fr o m s t u d i e s w i t h f l u o r e s c e n t protein ta g g e d i s o fo r m s , e x p r e s s e d fr o m c D N A v e c t o r s in t r a n s i e n t l y or p e r m a n e n t l y t r a n s f e c t e d c e l l line s. In t h e s e s t u d i e s E R , m i t o c h o n d r i a l o u t e r m e m b r a n e ( M O M ) an d c y t o s o l w e r e iden tif ie d a s the m ai n l o c a t i o n s for D M P K r e s i d e n c e a s o u t lin e d f u r t h e r b e l o w (2 86 , 296).
• 4. D M P K s p l i c e isoforms
• 4.1. A lt e r n a t i v e s p li c in g of D M P K isofor ms
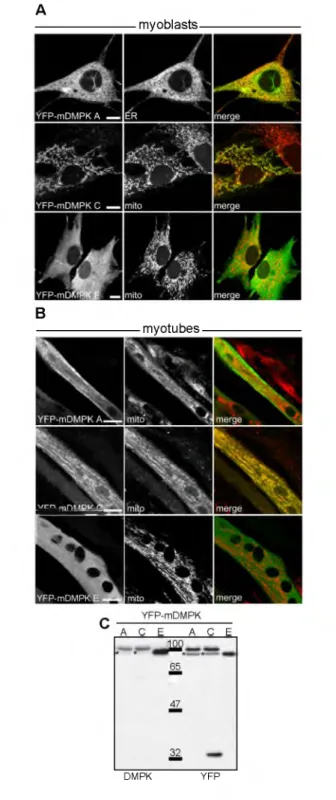
T r a n s c r i p t s fr o m the D M P K gene a r e s u b j e c t to e x t e n s i v e a l t e r n a t i v e s p li c in g , in a p r o c e s s t h a t is l a r g e l y c o n s e r v e d b e t w e e n m o u s e and man. A l t e r n a t i v e s p li c in g r e s u l t s in six m a j o r i s o f o r m s an d s e v e r a l mino r s p l i c e p r o d u c t s (95 ). The 15 ex o n s of the D M P K g e ne e n c o d e a protein w i t h a l e u c i n e - r i c h N - t e r m i n u s , a s e r i n e / t h r e o n i n e prot ein k i n a s e d o m a in (e x o n s 2-8), an a - h e l i c a l c o i l e d - c o i l d o m a in ( e x o n s 10-12) an d s e v e r a l d if f e r e n t C -t e rm in i (Fi g . 3) ( e x o n s 13-15). In one stu dy, a l t e r n a t i v e s p li c in g of D M P K t r a n s c r i p t s w a s i n v e s t i g a t e d in a t r a n s g e n i c h D M P K o v e r e x p r e s s o r m o u s e m o d e l, a l l o w i n g d ir e c t c o m p a r i s o n of h u m a n D M P K t r a n s g e n i c an d e n d o g e n o u s m o u s e D M P K ge ne p ro d u c t s . In thi s s t u d y it w a s found t h a t exon 8 c o n t a in e d an a l t e r n a t i v e 5' s p l i c e site, r e s u lt i n g in p ro d u c t io n of D M P K m R N A w i t h ( i s o f o r m s A, C an d E ) or w i t h o u t ( i s o
f o r m s B, D and F ) a 1 5 - n u c le o t id e s t r e t c h e n c o d i n g a V S G G G a m i n o ac id m o t if (Fi g . 3 B and C). A s e c o n d a l t e r n a t i v e l y s p l i c e d re g io n e n c o m p a s s e s ex o n s 12-15, r e s u lt i n g in the u s e of d if f e r e n t O R F s for the p ro d u c t io n of D M P K p r o t e i n s w i t h d if f e r e n t C-ter mi ni. I n c l u s i o n of ex o n s 12-15 r e s u l t s in D M P K i s o f o r m s A a n d B c o n t a in i n g a h y d r o p h o b i c C - t e m i n u s ( t a i l 1), w i t h a m o l e c u l a r size of ~70 kD a (Fig . 3 B and C). W h e n an a l t e r n a t i v e s p l i c e a c c e p t o r si te in exon 14 is us e d i n s t e a d of the n o r m a l 3' s p l i c e si te of intron 13, th e fi r s t four n u c l e o t i d e s of exon 14 a r e e x c l u d e d fro m the m R N A . T hi s use r e s u l t s in a f r a m e s h if t in the ORF, w h i c h t r a n s l a t e s into a l e s s h y d r o p h o b ic t a i l ( t a i l 2) of n e a r l y the s a m e size (F ig . 3 B and C). Fu si o n of ex o n s 12 an d 15, i.e., w i t h sk ip p in g of ex o n s 13-14, l e a d s to a f r a m e s h if t in th e O R F w i t h a stop cod on a p p e a r i n g at the beg inn ing of exon 15. At the prot ein p r o d u c t l e v e l th i s g i v e s ri se to a t r u n c a t e d C-ter- m i n u s c o m p o s e d of only t w o a m i n o a c i d s ( t a i l 3), w i t h a m o l e c u l a r size of a p p r o x i m a t e l y 60 kD a (fo r both D M P K E an d F, s e e Fig. 3 B and C) (95). A l t e r n a t i v e s p li c in g is use d d i f f e r e n t i a l l y in d if f e r e n t t i s s u e s . Long D M P K i s o f o r m s A-D a r e p r e d o m i n a n t l y e x p r e s s e d in s k e l e t a l m u s c l e , h e a r t and brain, w h e r e a s s h o r t i s o f o r m s E - F a r e m a i n l y found in s m o o t h m u s c l e (95, 211). A l l the a b o v e m e n t i o n e d m a t u r e t r a n s c r i p t s c o n t a in the (C U G ) n r e p e a t in the 3'- UT R, e i t h e r of n o r m a l l e ng t h w h e n t r a n s c r i b e d fr o m the h e a l t h y a l l e l e or of i n c r e a s e d l e n g t h w h e n o r ig in a t in g fr o m the m u t a t e d a l l e l e . • 4.2. Protein dom ai ns • 4.2.1. L e uc in e -r ic h N -t e rm in al domain T he t y p i c a l m o d u l a r s t r u c t u r e of D M P K an d the d if f e r e n t s e q u e n c e m o tif s in th e v a r i o u s m o d u l e s is w e l l p r e s e r v e d b e t w e e n m e m b e r s of th e AGC s u b f a m i l y of k i n a s e s . A l e u c i n e - r i c h N - t e r m i n a l reg ion of a b o u t 70 a m i n o a c i d s is c o m m o n l y found in M R C K s , RO CK s, C R IK an d D M P K (F ig . 3C). T hi s d o m a in m a y s e r v e in c o n t r o l l i n g the a c t i v i t y of the a d j a c e n t s e r i n e / t h r e o n i n e k i n a s e d o m a in in t h e s e p ro te in s, e i t h e r by f a c i l i t a t i n g linking to o t h e r s i g n a l i n g pr ot ei ns , by being a s c a f f o l d for in t e r a c t in g prot eins , or by i nd uc ing o l i g o m e r iz a t i o n of the protein. Dir ect ly, or in d ire ct ly, this d o m a in m a y a l s o be in v o l v e d in d ir e c t in g the s u b c e l l u l a r l o c a l i z a t i o n of the k i n a s e (184). S p e c i f i c e v i d e n c e for a ro l e of the l e u c i n e - r i c h d o m a in in m o d u la t in g D M P K a c t i v i t y h a s been p ro v i d e d by W a n s i n k et al. (2 9 7 ). The e l u c i d a t i o n of the c r y s t a l s t r u c t u r e of the D M P K k i n a s e d o m a in ( i n c l u d i n g th e N - t e r m i n u s ) r e v e a l e d t h a t t h e l e u c i n e - r i c h N - t e r m i n a l end c o u ld a l s o be i n v o l v e d in D M P K d im e r i z a t i o n (67, 89).
• 4.2.2. S er in e / th re o ni ne protein kina se domain
The k i n a s e d o m a in in D M P K (Fi g . 3C) h a s a l l the c h a r a c t e r i s t i c f e a t u r e s of c a t a l y t i c d o m a i n s in o t h e r k i n a s e s , e s p e c i a l l y th e hi ghl y a b u n d a n t s u b c l a s s of s e r i n e - t h r e o n i n e s p e c i f i c k i n a s e s . T y p ic a lly , a k i n a s e d o m a in c o n s i s t s of 11 m a j o r c o n s e r v e d s u b d o m a i n s s e p a r a t e d by i n s e r t i o n s and d e l e t i o n s (100, 297). In an a c t i v e c o n f o r m a t i o n ( " o n ” st a t e ), the k i n a s e d o m a in is fo ld e d into t w o l o b e s fo rm i n g the a c t i v e cl e ft . A g l y c i n e - r i c h s e q u e n c e ( G X G X 0 G , 0 is u s u a l l y p h e n y l a l a n i n e or t y r o s i n e ) in th e c l e f t is a c o m p o n e n t of th e h i g hl y c o n s e r v e d p h o s p h a t e - b i n d i n g loop, c a l l e d 1 • G e n e ra l In tr o d u c ti o n 19
1 • G e n e ra l In tr o d u c ti o n P - lo o p, c a p a b l e of b ind ing A TP (67, 100, 117, 27 3) . P r o p e r a l i g n m e n t of A TP for c a t a l y s i s is c o n t r o l l e d by the m ai n ch a i n n i t r o g e n s of the P loop.
Fo r D M P K th i s fu n c ti o n is s e r v e d by the hi ghl y c o n s e r v e d l ys in e r e s i d u e at p os it io n 100, w h i c h m a k e s c o n t a c t w i t h the a an d ß p h o s p h a t e s of A TP (117).
U n li k e in th e " o n ” s t a t e in w h i c h k i n a s e s a d o p t a def in e d s t r u c t u r e , protein k i n a s e s in the " o f f ” s t a t e a r e not in a c h e m i c a l l y c o n s t r a i n t s t a t e an d t h e r e f o r e a b l e to ad o p t v a r i o u s s t r u c t u r e s . The m o s t f r e q u e n t l y d i s c u s s e d m e c h a n i s m s of i n a c t iv a t io n in v o l v e e i t h e r m a j o r r e a r r a n g e m e n t s or d i s o r d e r of the a c t i v a t i o n loop or a C helix in th e N - t e r m i n a l lobe, w h i c h r e s u l t s in d i s p l a c e m e n t of a c t i v e si te r e s i d u e s or in b l o c k a g e of the A T P s u b s t r a t e binding si te (117, 210, 273). I m m e d i a t e l y C - t e r m i n a l of the k i n a s e d om a in , D M P K c o n t a i n s an a d d it i o n a l d o m a in , w h i c h is p r e s u m e d to be in v o l v e d in k i n a s e a c t iv a t io n . A s i m i l a r a r r a n g e m e n t is found in a p p r o x i m a t e l y 40 o t h e r AG C k i n a s e s an d in s e v e r a l of t h e s e k i n a s e s th e ro l e of thi s a d d i t i o n a l d o m a in h a s been s t u d i e d e x t e n s i v e l y . T hi s s o - c a l l e d C - t e r m i n a l e x t e n si o n of th e k i n a s e d o m a in c o n t a i n s a c o n s e r v e d h y d r o p h o b ic p h o s p h o r y l a t i o n motif, F X X [ F / Y ] [ S / T ] [ Y / F ] , w h i c h c a n be a c t i v a t e d by p h o s p h o r y l a t i o n (17, 84, 124, 296). For s o m e k i n a s e s it h a s been s h o w n t h a t w h e n the hy d ro p h o b ic
p h o s p h o r y l a t i o n m o t if is p h o s p h o r y l a t e d it r e c r u i t s p h o p h o i n o s i t i d e - d e - p en d e n t ki na se- 1, w h i c h ca n a c t i v a t e th e k i n a s e by p h o s p h o r y l a t i n g the a c t i v a t i o n loop (17, 117, 210). The VS G G G m o t if l o c a t e d in the C - t e r m i n a l e xt e n si o n of th e k i n a s e d o m a in w a s found to be in v o l v e d in a c t i v a t i o n of D M P K in both t r a n s - an d a u t o p h o s p h o r y l a t i o n (2 9 6 ). A c t i v a t i o n of the k i n a s e ca n be inh ibited by s t e r i c h i n d r a n c e of th e C - t e r m i n a l end of the d if f e r e n t D M P K i s o fo r m s , a m e c h a n i s m w h i c h is a l s o u s e d by o t h e r D M P K f a m i l y m e m b e r s l ik e R O C K II a n d M R C K a (165, 270, 29 6) . F i n a ll y , the C - t e r m i n a l e x t e n s i o n of the k i n a s e d o m a in is i nv o lv e d in d im e ri z a t i o n of D M P K by b ind ing to t h e l e u c i n e - r i c h N - t e r m i n a l r e g io n (6 7 ). D M P K is a s e r i n e / t h e o n i n e prot ein k i n a s e t h a t f a v o r s t h r e o n i n e o ve r s e r i n e and p r e f e r s p r e s e n c e of at l e a s t t h r e e a r g i n i n e s or l y s i n e s a m o n g t he f i v e r e s i d u e s N - t e r m i n a l to th e p h o s p h o a c c e p t o r s i t e (2 9 6 ). A lt h o u g h a u t o p h o s p h o r y l a t i o n of D M P K w a s o b s e r v e d , no si t e in the prot ein i t s e lf m a t c h e s the c o n s e n s u s s e q u e n c e . B e s i d e s a l l p u t a t i v e s u b s t r a t e s only a f e w n a t u r a l D M P K s u b s t r a t e s h a v e been found ( s e e l a t e r s e c t i o n s ) , s h o w ing l im it e d p r e d i c t i v e p o w e r of the c o n s e n s u s s e q u e n c e .
• 4.2.3. Coile d- coi l region
The s e r i n e / t h r e o n i n e prot ein k i n a s e d o m a in of D M P K is f o l l o w e d by an a - h e l i c a l c o i l e d - c o i l reg ion (F ig . 3C) (26, 95). The c o i l e d - c o i l s t r u c t u r e w a s fi r s t d e s c r i b e d in 1953 a s the m a i n s t r u c t u r a l e l e m e n t in a - k e r a t i n (49, 50), an d w a s l a t e r found in o t h e r p r o t e i n s lik e t r o p o m y o s in , m y os in and fi br in og e n (28, 96, 188). C o i l e d - c o i l d o m a i n s a r e of ten p r e s e n t in e u k a r y o t i c p ro te in s, and m o re i m p o rt a n t ly , in a l l m e m b e r s of th e D M P K f a m i l y (166, 173, 297). M o s t m e m b e r s of th e D M P K f a m i l y c o n t a in long c o i l e d - c o i l re g io n s of at l e a s t 600 a m i n o a c i d s (2 9 7 ) . T he c o i l e d - c o i l re g io n of D M P K i t s e lf is m u c h s m a l l e r a n d e n c o m p a s s e s o nl y ~ 6 5 r e s i d ue s (95). T h e c o i l e d c o i l r e g io n of D M P K is r e s p o n s i b l e for th e f o r m a t i o n of m u l t i m e r i c c o m p l e x e s (2 8 7 ) . M u l t i m e r i z a t i o n of D M P K i n f l u e n c e s s u b s t r a t e
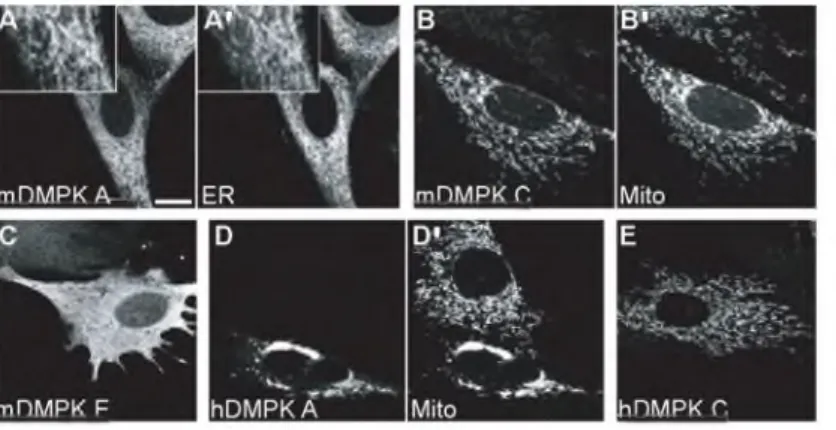
Figure 4 • Su bcellular localization of DMPK splice isoforms. N - te r m in a l Y F P - D M P K fu s io n p ro te in s w e r e e x p r e s s e d in D M P K KO m y o b la s ts and v is u a liz e d by c o n fo c a l la s e r s c a n n in g m ic ro s c o p y . ER w a s c o u n te r - s ta in e d u s in g a c a lr e t ic u lin a n tib o d y ; m i t o c h o n d r ia w e r e v is u a liz e c u s in g M ito t r a c k e rR e d . (A, A ') Y F P - m D M P K A is a s s o c ia te d to th e e n d o p la s m ic r e t ic u lu m . (B , B ') Y F P - m D M P K C is lo c a te d a t m it o c h o n d ria. (C ) Y F P - m D M P K E is lo c a t e d in th e c y to s o l. (D, D ') Y F P - h D M P K A a n d ( E ) Y F P - h D M P K C a re b oth a s s o c ia te d to m ito c h o n d r ia . O nly Y F P - h D M P K A in d u c e s s tro n g m it o c h o n d r ia l a g g re g a tio n a ro u n d th e n u c le u s . B a r = 10 pm.
binding an d i n c r e a s e s prot ein k i n a s e a c t i v i t y (199, 287, 289). L i k e w i s e , the a c o i l e d - c o i l r e g io n s in R O CK II and M R C K i n f l u e n c e k i n a s e a c t iv it y , a lt h o u g h in t h e s e t w o f a m i l y m e m b e r s a c t i v i t y is inh ibi ted by c o i l e d - c o i l i n t e r a c t i o n s (5, 270). A d d i ti o n a ll y , c o i l e d - c o i l m e d i a t e d m u lt i m e r iz i n g is in v o l v e d in a n c h o r i n g of i so fo r m D M P K C to its s p e c i f i c l o c a l i z a t i o n at the m i t o c h o n d r i a l o u t e r m e m b r a n e (2 86 , 287).
• 4.2.4. C -te rm ina l an ch or
The six m a j o r D M P K i s o f o r m s found in m o u s e and h u m a n s c a r r y t h re e
d is t i n c t C - t e r m i n a l t a i l s (Fi g . 3C) (95, 296). T a i l s 1 ( D M P K A an d B ) and 2 ( D M P K C an d n) a r e c o m p r i s e d of 96 an d £3 7 a m i n o a c i d s r o s c e c t i v e l y , an d a r e c o n s i d e r a b l y l o n g e r th a n t a i l 3 ( D M P P E and F), w h i c h c o n s i s t of onty t w a a m i n o ac id rd ai du es . A c m e n t i o n e d e a rl i e r , s e v a o a l s t u d i e s ha v e s h o w n ttiat t a i l d o m a i n s of th e d if fe re nt D M P K ^ o f o r m s i n f l u e n c e ac ti v- ity of thn photein D ina se d o m r i n , or its s d b s t r a t d e p e c i f i c i t y (1S 9, ° 9 6 ) . F u r t h e r m o r e , C - t e r m i n a l t a i l s a r e in v o l v e d t a r g e t i n g of D M P K i s o f o r m s to a s p e c i f i c s u b c e l l u l a r l o c a l i z a ti o n .
In m o u s e , D M P K t a i l 1 i n d u c e s E R l o c a l i z a t i o n , p r e s e n c e of t a i l 2 r e s u l t s in t a r g e t i n g to the MOM an d a c y t o s o l i c l o c a l i z a t i o n is o b s e r v e d for t a i l 3 (Fi g . 4 A - C ) (2 86 , 296). It h a s been found t h a t a l l i n fo rm a t i o n n e c e s s a r y for t a r g e t i n g to the E R is p r e s e n t in th e t a i l 1, w h e r e a s for t a i l 2, a d d i t i o n a l p r e s e n c e of the c o i l e d - c o i l reg io n is a r e q u i s i t e for p ro p e r MOM t a r g e t i n g (2 8 6 ). U n li k e t a i l 3, w h i c h d o e s not c o n t a in t a r g e t i n g in f o r m a t i o n (F ig . 4C), t a i l 1 an d 2 a n c h o r f i r m l y at t h e i r t a r g e t m e m b r a n e s . A l t h o u g h t h e r e is s t ro n g h o m o l o g y b e t w e e n m o u s e an d h u m a n D M P K g e n e s, a s p e c i e s d i f f e r e n c e w a s o b s e r v e d for i s o f o r m s A and B: m o u s e D M P K A and B t a r g e t to the ER , the h u m a n o r t h o l o g u e s a r e l o c a l i z e d at th e MO M ( F i g . 4D a n d E). F u r t h e r m o r e , t r a n s i e n t e x p r e s s i o n of h D M P K A i n d u c e s m i t o c h o n d r i a l c l u s t e r i n g ar o u n d the n u c l e u s , u l t i m a t e l y lea din g to c e l l dea th. E x p r e s s i o n of m o u s e an d h u m a n D M P K C and D a l s o r e s u l t s in l o c a l i z a t i o n to the MOM, but g i v e s not ri se to p e r i n u c l e a r m i t o c h o n d r i a l c l u s t e r i n g (2 8 6 ). C o m p u t e r a n a l y s i s r e v e a l e d h y d r o p h o b ic r e g io n s in tai l 1 an d 2, c l a s s i f y i n g t h e m a s m e m b r a n e a n c h o r s . T hi s s p e c i f i e s long i s o f o r m s A-D a s t a i l - a n c h o r e d (T A ) pr ot e in s. It is s t i l l u n c e r t a i n in w h a t w a y D M P K i s o f o r m s a r e i n t e g ra t e d in the lipid b i l a y e r (t h i s t h e s i s ) and if the y re q u i r e a d d i t i o n a l f a c t o r s to f a c i l i t a t e m e m b r a n e t a rg e ti ng . 1 • G e n e ra l In tr o d u c ti o n 21
1 • G e n e ra l In tr o d u c ti o n TA p r o t e i n s a r e def in e d by a o y t o s o l io N - t e r m i n a l d o m a in and a s t r e t c h of h y d r o p h o b ic r e s i d u e s in the C - t e r m i n a l region, w h i c h a l l o w s a n c h o r i n g in th e p h o s p h o li p i d b i l a y e r of m e m b r a n e s (18, 19, 22 2) . TA p r o t e i n s o c c u r at a l l m e m b r a n e s of the c e l l and p la y key r o l e s in n u m e r o u s c e l l u l a r p r o c e s s e s in c lu d in g v e s i c u l a r t r a n s p o r t (119, 122), prot ei n t r a n s l o c a t i o n (69, 2 54 ) an d a p o p t o s i s (137).
W i t h th e N - t e r m i n a l b ul k of th e prot ein l o c a t e d in th e c y t o s o l , TA p r o t e i n s a r e bound to the m e m b r a n e by an a n c h o r t h a t is u s u a l l y lo c at e d w i t h in 30 a m i n o a c i d s fro m th e N - t e rm i n u s . TA p r o t e i n s do not co n t ai n a s i g n a l s e q u e n c e , and s i n c e the h y d r o p h o b ic reg ion e m e r g e s fr o m the r i b o s o m e upon t e r m i n a t i o n of t r a n s l a t i o n , TA p r o t e i n s a r e not s u b j e c t to c o - t r a n s l a t i o n a l t r a n s l o c a t i o n (18). S o m e of t h e s e f e a t u r e s d is t i n g u is h t h e m fr o m t y pe II t r a n s m e m b r a n e p ro te in s, w h i c h a l s o e x p o s e th e i r N- t e r m i n u s to the c y t o s o l but c o n t a in a m e m b r a n e a n c h o r c l o s e r to th e ir • 5. Tail-anchored proteins N - t e rm i n u s . F u r t h e r m o r e , ty pe II t r a n s m e m b r a n e p r o t e i n s a r e c o t r a n s - l a t i o n a l l y t r a n s l o c a t e d w i t h o u t c o n t a in i n g a c l e a v a b l e s i g n a l s e q u e n c e . R e c e n t pi si p e r s h a g e sh o w n t h a g Sh e m a x i m a I n u mb er of hy d ro p h i li e a m i n o a c i d s t h a t ca n lie a p p e n d e d to the m e m b r a n e a n c h o r w i t h o u t h a m p g ri n g m s m b r a e e t r a n s l o c a t i o n ca n r e a c h up to 85 r e s i d u e s , a l t h o u g h one m u s t b e a r in mind t h a t ex te n di ng the C - t e r m i n a l p o l a r d o m a in w i l l e v e n t u a l l y r e s u l t in t y pe II t r a n s l o c a t i o n (18, 24, 47). Us ing t r u n c a t i o n a n a l y s i s it h a s been s h o w n t h a t t a r g e t i n g i n f o r m a tion r e q u i re d for t r a n s l o c a t i o n of TA prot ein t h r o u g h th e i r t a r g e t m e m b ra n e is c o n t a in e d w i t h in the C - t e r m i n a l re g io n (24, 207, 209). A lt h o u g h TA p r o t e i n s end up in m a n y c e l l u l a r m e m b r a n e s t h e y i n i t ia ll y only t a r g e t to a lim it e d n u m b e r of m e m b r a n e s . A f e w TA p r o t e i n s a r e a b l e to a n c h o r in m u l t i p l e m e m b r a n e s ; m i t o fu s i n 2 ( 5 5 ) and B c l 2 (141), for e x a m p l e , can t a r g e t to E R or MOM. To r e a c h d if f e r e n t c o m p a r t m e n t s of th e s e c r e t o r y
Figure 5 • Routing of TA proteins in anim al cells.
W h e n r e le a s e d fro m the rib o s o m e a TA p ro te in (s t r u c tu r e w ith b la c k N- end a n d g re y C - e n d ) c a n in s e rt in to th e E R (1 ). T r a n s lo c a tio n to o th e r c o m p a r tm e n t s of th e e n d o - e x o c y tic p a t h w a y is a c h ie v e d by v e s i c u l a r t r a n s port (d o tte d b la c k a rro w ). T a rg e tin g to th e MOM (2 ) or th e p e r o x is o m a l m e m b r a n e ( 4 ) is a ls o d ire c t, a lth o u g h an a lt e r n a t iv e w a y fo r TA p ro te in s to r e a c h th e p e ro x i s o m e is v ia in s e rtio n in to a s p e c ia li z e d E R s u b d o m a in (3 ), f o llo w e d by m a tu ra tio n of t h is s u b d o m a in in to a f u lly d e v e lo p e d p e ro x is o m e (d o tte d g re y a r ro w ). In p la n ts , TA p ro te in s a ls o d i r e c t ly ta r g e t to c h lo r o p la s ts . A d a p te d fr o m B o r g e s e et a l (1 8 ). ~
Figure 6 • TA features determine targeting of TA- proteins to the ER, MOM or peroxisomes. O r g a n e lle - s p e c if ic ta rg e tin g of T A - p ro te in s, w h ic h d e p e n d s m a in ly on th e le n g th of th e h y d ro p h o b ic s tre tc h a n d p re s e n c e of fla n k in g c h a rg e d a m in o a c id s . TA p ro te in s a re d e p ic te d w ith a fo ld e d c y t o s o lic d o m a ir f o llo w e d by a v a r ia b ly - s ize d h y d ro p h o b ic d o m a in (r o u n d e d g re y r e c ta n g le ). C h a rg e d r e s id u e s fla n k in g t h e h y d ro p h o b ic d o m a in a re in d ic a te d a s + a n d -. (A ) A s h o rt h y d ro p h o b ic d o m a in fla n k e d by p o s itiv e ly c h a rg e d r e s id u e s d r iv e s TA p ro te in s to th e M OM . (B / C ) L o s s of e it h e r of t h e s e f e a t u r e s - a s h o rt h y d ro p h o b ic d o m a in or fla n k in g c h a r g e c r e s id u e s - r e s u lt s in t a r g e t ing to th e ER . (D ) C - te rm in i w ith in t e r m e d ia t e f e a tu re s - s lig h t ly le n g th e n e d h y d ro p h o b ic d o m a in and/ or re d u c e d p o s itiv e c h a r g e - in d u c e ta r g e tin g to e ith e r th e MOM or ER. ( E ) For t r a n s lo c a t io n to p e ro x is o m e s , a s h o rt h y d ro p h o b ic d o m a in is fla n k e d by a m o d e ra te p o s itiv e c h a rg e . S e q u e n c e - s p e c if ic f e a t u r e s lik e a re c o g n itio n s e q u e n c e (b la c k o v a l ) f a c ilit a t e d ire c t in s e r tio n in to th e p e ro x is o m e (E ). A d a p te d fro m B o r g e s e et a l (2 0 ). p a t h w a y , TA p r o t e i n s fi r s t i n s e r t into the E R an d v i a v e s i c u l a r t r a n s p o r t t r a n s l o c a t e to t h e i r f i n a l d e s t i n a t io n (Fi g . 5). H o w th i s j o u r n e y is e f f e c t u a t e d is d e t e r m i n e d by t h e lipid c o m p o s i t i o n of th e d if f e r e n t m e m b r a n e s in de s e c r e t o r y p a t h w a y i.e., by th e c h o l e s t e r o l c o n c e n t r a t i o n (18, 24) E R t o g e t h e r w i t h the: l e ng t h an d h y d r o p h o b i c i t y of the m e m b r a n e a n c h o r (20, 27). O v e r a l l in a n i m a l c e l l s , TA p r o t e i n s e i th e r i n s e r t int o the ER m e m b r a n e , MO M, a n d in s o m a c a s e s d i r e c t l y into I h e p e r o x i s o m a l m e m b ra n e ( F i g . 5 ) (18, 20). In tine fo l lo w i n g s e c t i o n p o s s i b l e m e c h a n i s m s and a n c h o r i n g r e q u i r e m e n t s w i l l be f u r t h e r d i s c u s s e d . • 5.1. Targeting to the E R m e m br an e A lt h o u g h E R t a r g e t i n g r e q u i r e m e n t s a r e v a r i a b l e , t a r g e t i n g to the E R is c o n t r o l l e d by a r e l a t i v e l y long (>20 a m i n o a c i d s ) s t r e t c h of hy d ro p h o b ic r e s i d u e s w i t h high h y d r o p h o b ic s t r e n g t h , f l a n k e d by a v a r i a b l e a m o u n t of c h a r g e d r e s i d u e s (Fi g . 6) (2 0). I n s e r t i o n of TA p r o t e i n s into th e ER m e m b r a n e is d ir e c t an d an A T P - d e p e n d e n t p r o c e s s , a lt h o u g h th e a m o u n t of A T P r e q u i re d for m e m b r a n e i n s e rt i o n d i f f e r s b e t w e e n d if f e r e n t TA p r o teins. C y t o c h r o m e b5 is a b l e to i n s e r t into p r o t e a s e - t r e a t e d m i c r o s o m e s in the n e a r a b s e n c e of ATP, w h i l e ad d it io n of A T P d o e s not i n c r e a s e in t e g ra t i o n into the E R m e m b r a n e s i g n i f i c a n t l y (24, 305). It is h y p ot h e si z e d t h a t th i s a b i l i t y to i n d e p e n d e n t l y i n s e r t into the E R m e m b r a n e r e q u i r e s an E R m e m b r a n e a n c h o r w i t h a m o re m o d e r a t e hy d ro p h o b ic it y . S u c h an a r r a n g e m e n t w i l l give th e prot ein h ig he r s o l u b i l i t y in the c y t o s o l (18). TA p r o t e i n s w i t h a hig hly h y d r o p h o b ic tail, like s y n a p t o b r e v i n 2 ( S y b 2 ) , ar e m o r e pr on e to fo rm a g g r e g a t e s in th e c y t o s o l an d t h e r e f o r e need ATP c o n s u m i n g c h a p e r o n e s to r e m a i n s o l u b l e and to i n s e r t p r o p e r l y into the m e m b r a n e (24, 155). In t e re s t in g ly , the E R m e m b r a n e h a s been s h o w n to be th e m o s t p e r m i s s i v e m e m b r a n e for a c c e p t i n g TA pr ot ei ns . TA pr ot ei ns , in vi v o t a r g e t e d to th e MOM, a r e a b l e to i n s e r t into m i c r o s o m e s in v i tro, d e m o n s t r a t i n g t h a t E R m e m b r a n e is c a p a b l e of t a k i n g up m a n y TA p r o t e i n s w h e n it d oe s not need to c o m p e t e w i t h o t he r m e m b r a n e s (18). TA p r o t e i n s t h a t t a r g e t to the p e r o x is o m e c a n a l s o t r a v e l v i a the ER , in th e s o - c a l l e d p E R m e d i a t e d ro ut e (Fi g . 5), a s h a s bee n s h o w n
<
1 • G e n e ra l In tr o d u c ti o n 231 • G e n e ra l In tr o d u c ti o n for Pex15p an d a s c o r b a t e p e r o x i d a s e (66, 142, 203). A l t e r n a t i v e l y , d ir e c t p e r o x i s o m a l t a r g e t i n g w a s found for Pe x 2 6 an d h F i s v i a binding to the Pex19 r e c e p t o r (Fi g . 5) (99, 108). S u r p r i s i n g l y , the t a i l d o m a in in p e r o x i s o m e - t a r g e t e d TA p r o t e i n s h a s only mild h y d r o p h o b ic i t y an d c o n t a i n s a p o s i t i v e l y c h a r g e d C - t e r m i n u s (F ig . 6) (18).
The d e p e n d e n c e on A T P for E R t a r g e t i n g u s u a l l y s u g g e s t s a need for m o l e c u l a r c h a p e r o n e s , a lt h o u g h for the m a j o r i t y of c a s e s it is not k n ow n w h i c h c h a p e r o n e s a r e inv o lv e d. S e v e r a l w a y s of i n s e rt i o n ha v e been i n v e s t i g a t e d and it w a s s h o w n by High's group, t h a t t o g e t h e r w i t h A T P or GTP the s i g n a l re c o g n i t i o n p a r t i c l e ( S R P ) co u ld i n t e r a c t w i t h S y b 2 and S e c 6 1 ß , an d in th i s w a y w a s in v o l v e d in i n s e rt i o n into the E R m e m b r a n e (1). T h e s e r e s u l t s a r e c o n t r o v e r s i a l , h o w e v e r , and not s u p p o r t e d by other g ro u p s w h o s h o w e d d e p e n d e n c e on A T P but not GTP or the S R P (155, 264, 265). A n o t h e r i n t e r a c t io n t h a t f a c i l i t a t e s A T P - d e p e n d e n t in te g ra t i o n of TA p r o t e i n s i n v o l v e s m o l e c u l a r c h a p e r o n e s H sp 4 0 and H sc 7 0 (2). Fi na lly , the m o s t w e l l - d e f i n e d t a r g e t i n g f a c t o r is the 40 kD a c y t o s o l i c protein A s n a - 1 / T R C 4 0 , a prot ein w i t h A T P a s e a c t iv i t y . Asna -1 ca n i n t e r a c t post- t r a n s l a t i o n a l l y w i t h th e m e m b r a n e - a n c h o r i n g reg io n of TA p r o t e i n s and t h e r e b y d e l i v e r the prot ei n to a p r o t e i n a c e o u s r e c e p t o r at the E R m e m b r a n e (2 6 5 ). E x p r e s s i o n of A T P a s e - d e f e c t i v e m u t a n t s of A s n a r e s u lt e d in a n in h ib i to ry e f f e c t on TA pr ot e in in se rt io n , but did not i n f l u e n c e o the r t r a n s l o c a t i o n p a t h w a y s . T hi s d e m o n s t r a t e s t h a t i n v o l v e m e n t of A TP h y d r o l y s i s is e s s e n t i a l . B a s e d on t h e s e fi n d in g s a m e c h a n i s t i c m o d e l w a s pr o p o se d , w h e r e b y a r e c e p t o r on the E R m e m b r a n e s t i m u l a t e s Asna-1 ac t iv i t y , a l l o w i n g r e l e a s e of the TA s u b s t r a t e an d i n s e rt i o n of the m e m b r a n e a n c h o r into the m e m b r a n e (2 6 5 ) . R e c e n tl y , a y e a s t h o m o l o g u e for A sna -1, Get3 w a s ide nti fi ed w h i c h s h o w e d s i m i l a r c h a r a c t e r i s t i c s and w a s found to f a c i l i t a t e TA protein i n s e rt i o n v i a binding to th e E R r e c e p t o r c o m p l e x G e t1 / G e t2 (114, 189, 251, 268).
I n c r e a s i n g k n o w l e d g e of m e m b r a n e a n c h o r i n g and u n d e rl y i n g t a r g e t ing m e c h a n i s m s of TA p r o t e i n s is of g r e a t v a l u e in u n d e r s t a n d i n g the d is t ri b u t i o n an d c h a r a c t e r i s t i c s of D M P K i s o fo r m s . Not m u c h w a s kn ow n re g a r d in g D M P K m e m b r a n e a n c h o r i n g at th e s t a r t of th i s t h e s i s period.
• 5.2. Targeting to the MOM
The g e n e r a l c o n s e n s u s for MOM t a r g e t i n g is d e p e n d e n t on a m e m b r a n e a n c h o r of m o d e r a t e hy d ro p h o b ic i ty , fl a n k e d by b a s i c a m i n o a c i d s (Fi g . 6). A lt h o u g h e x c e p t i o n s e x is t (7) , a B x 0-9B x 0-2-TM D-x 0-1B x 0-6B ( w h e r e B s t a n d s for b a s i c r e s i d u e s and x for a n y a m i n o a c i d ) c o n s e n s u s s e q u e n c e h a s been p ro p o s e d (137). C o r r e c t p o si ti on in g of e a c h r e s i d u e in th i s s e q u e n c e is e s s e n t i a l a s it w a s found t h a t a l t e r in g e i t h e r one r e s i d u e - f.e. by d e l e tion of p o s i t i v e c h a r g e s or i n c r e a s i n g TMD l e ng t h - l e a d s to d e c r e a s e d MO M bin ding or e v e n r e r o u t i n g to th e E R (18, 61, 113, 119). S t i l l , p r e d i c ti o n s s o l e l y b a s e d on h y d r o p h o b ic i t y and c h a r g e of r e l e v a n t s e q u e n c e s m a y not be a c c u r a t e and a d d i t i o n a l s i g n a l s , p r o b a b l y in the N - t e r m i n u s of th e protein, c o n t r ib u t e to t a r g e t i n g (3, 118, 257).
S i n c e t r a n s l o c a t i o n to the MO M is d i r e c t and d o e s not o c c u r v i a the ER (18) (Fi g . 5), the q u e st io n h a s been r a i s e d w h e t h e r the c l a s s i c a l m i t o c h o n d r i a l t r a n s l o c a t i o n m a c h i n e r y (i.e., TOM c o m p l e x ) is in vo lv ed in i n s e r t i n g TA protein into th e MOM. The a n s w e r c a m e fr o m t w o r e c e n t s t u d i e s in m a m m a l i a n and y e a s t c e l l s , w h i c h r e v e a l e d t ha t t a r g e t i n g of