0095-1137/04/$08.00⫹0 DOI: 10.1128/JCM.42.11.5022–5028.2004
Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Use of Multilocus Variable-Number Tandem-Repeat Analysis for
Typing
Mycobacterium avium
subsp.
paratuberculosis
Pieter Overduin,
1Leo Schouls,
2Paul Roholl,
3Adri van der Zanden,
4Nofel Mahmmod,
4Arnold Herrewegh,
1and Dick van Soolingen
1*
Mycobacteria Reference Laboratory,1Laboratory of Toxicology, Pathology, and Genetics,3and Laboratory for
National Vaccination Program Control,2National Institute of Public Health and the Environment,
Bilthoven, and Medical Microbiology and Infectious Diseases and Gastroenterology Department,
Gelre Hospital, Apeldoorn,4The Netherlands
Received 13 February 2004/Returned for modification 29 March 2004/Accepted 2 August 2004
The etiology of Crohn’s disease in humans is largely unknown. Clinical signs of Crohn’s disease partly
resemble the clinical picture of Johne’s disease in ruminants caused byMycobacterium aviumsubsp.
paratu-berculosis. Because of the high prevalence of these bacteria in (products of) ruminants and their remarkable thermostability, concern has been raised about the possible role of these bacteria in the pathogenesis of Crohn’s disease. In an attempt to develop a molecular typing method to facilitate meaningful comparative DNA
fingerprinting ofM. aviumsubsp.paratuberculosisisolates from the human and animal reservoirs, multilocus
variable-number tandem-repeat analysis (MLVA) was explored and compared to IS900restriction fragment
length polymorphism (RFLP) typing. MLVA typing subdivided the most predominant RFLP type, R01, into six
subtypes and thus provides a promising molecular subtyping approach to study the diversity ofM. aviumsubsp.
paratuberculosis.
Paratuberculosis, also named Johne’s disease in ruminants, is characterized by a chronic inflammation of the ileum and is caused byMycobacterium aviumsubsp.paratuberculosis.
In humans, the symptoms of Crohn’s disease partly resemble those of Johne’s disease in ruminants (7). However, the etiol-ogy of Crohn’s disease is much more complex and appears to be multifactorial (16, 27).
The finding that M. avium subsp.paratuberculosis has re-markable thermostability during pasteurization of milk (12) and the high prevalence of these bacteria both dead and alive in milk for consumers (13, 20) imply a regular exposure of humans to these bacteria. The role microbial agents, and es-peciallyM. aviumsubsp.paratuberculosis, may play in the in-duction of the immunological disorder characteristic of Crohn’s disease has not yet been established (16, 27). In some studies, M. avium subsp. paratuberculosis was isolated from intestinal biopsies or feces of Crohn’s disease patients (6, 10, 11, 19, 23). Furthermore, a recent study in the United King-dom revealed a significant correlation between the presence of
M. aviumsubsp.paratuberculosisin intestinal biopsies and
ac-tive Crohn’s disease (2). Others did not find evidence for a relation between Crohn’s disease and the presence ofM. avium subsp.paratuberculosis(4, 5). Unfortunately,M. aviumsubsp.
paratuberculosisis very difficult to cultivate from human
biop-sies. Culture in liquid medium followed by PCR of presumed growth ofM. aviumsubsp.paratuberculosishas increased the number of biopsies in whichM. aviumsubsp.paratuberculosisis suspected (23). These cultures, however, seldom yield suffi-cient DNA for restriction fragment length polymorphism
(RFLP) typing experiments, and many isolates fail to grow in sufficient quantities for RFLP typing.
Recently, pulsed-field gel electrophoresis of pigmented and nonpigmented M. avium subsp. paratuberculosis isolates re-sulted in a subdivision of a part of the IS900 RFLP types, providing more refined data for epidemiological interpretation (24). Unfortunately, the latter technique also requires large quantities of DNA.
Therefore, the question of whether these hardly cultivatable
M. aviumsubsp.paratuberculosisstrains are of the same
geno-type as animal M. avium subsp. paratuberculosis isolates re-mains unanswered. In view of this, a PCR-based typing tech-nique for low quantities of bacterial DNA, as present in human intestinal biopsies, would be a valuable extension of typing methods for the molecular epidemiology of Crohn’s disease.
Several PCR-based techniques for identification and typing
ofM. aviumsubsp.paratuberculosisstrains have been described
in the last few years. Whittington et al. (30, 31) and Marsh et al. (18) have described IS1311 PCR followed by restriction enzyme analysis to differentiate between cattle and sheep type strains and otherM. aviumcomplex isolates. However, these techniques have so far failed to distinguish betweenM. avium subsp.paratuberculosisisolates.
Another promising PCR-based method to study the epide-miology of tuberculosis is mycobacterial interspersed repetitive unit (MIRU) typing. This method is based on variation in the number of MIRUs of 40 to 100 bp in length. MIRUs are arranged mostly in tandem repeats and are dispersed in inter-genic regions ofM. tuberculosis complex bacteria (9, 25, 26). Not all genomic loci with MIRUs have been fully explored, but in several studies, variable-number tandem-repeat (VNTR) typing ofM. tuberculosiscomplex isolates has revealed a higher resolution than that achieved with IS6110RFLP typing (17).
Bull et al. (3) found four MIRU loci which can be used to
* Corresponding author. Mailing address: National Institute of Pub-lic Health and the Environment (RIVM), P.O. Box 1, 3720 BA Bilthoven, The Netherlands. Phone: 31 30 2742363. Fax: 31 30 2744418. E-mail: [email protected].
5022
on May 15, 2020 by guest
http://jcm.asm.org/
discriminate betweenM. aviumsubspeciesparatuberculosis,M.
aviumsubspeciesavium, andM. intracellularestrains.
Further-more, cattle and sheep strains were divided into four different groups on the basis of MIRU typing.
The aim of our study was to investigate the usefulness of multilocus VNTR typing analysis (MLVA) as a tool for typing
M. avium subsp. paratuberculosis strains and to investigate
whether a higher degree of discrimination can be reached amongM. aviumsubsp.paratuberculosisisolates.
MATERIALS AND METHODS
Strains.M. aviumsubsp.paratuberculosisstrains were cultured on Herrold’s egg yolk medium according to the method of Whipple et al. (29). The host species included cattle, sheep, red deer, fallow deer, roe deer, and humans.
Strains from the Czech Republic, Sweden, and the United States were ob-tained from the Veterinary Research Institute, Brno, Czech Republic. Human isolates of Dutch origin were obtained from the Institute for Animal Science and Health, Lelystad, The Netherlands. The isolates were identified asM. avium
subsp.paratuberculosisby cultural and biochemical properties, and this identifi-cation was confirmed by DNA-DNA hybridization (14). The cattle isolates from The Netherlands were obtained from the Animal Health Service, Deventer, The Netherlands. All isolates from Argentina were obtained from the Instituto Na-cional de Tecnologia Agropecuaria, Buenos Aires, Argentina, and the isolate from Venezuela was obtained from the Biolac United Nations University Insti-tute, Baruta, Venezuala. DNA samples of theM. aviumsubsp.paratuberculosis
strains SN 1 to SN 8, K05, K16, and K18 were kindly donated by the St. George Hospital Medical School Department of Surgery, London, United Kingdom.
IS900RFLP typing.IS900RFLP typing is currently the standard method for typingM. aviumsubsp.paratuberculosis.This method has the highest discrimi-natory power and serves as the “gold standard” for MLVA analysis. IS900RFLP type, geographic region, and host were used for the selection of strains to build a heterogeneous strain panel. IS900RFLP typing was performed as described previously by van Soolingen et al. (28), with some modifications. Lysozyme incubation was performed overnight and proteinase K-sodium dodecyl sulfate treatment was done for 20 min at 65°C.
Digestion was performed with 3g of DNA and 7 U of BstEII (Roche) at 37°C for at least 4 h. Separation of the DNA restriction fragments was done on a 0.8% MP agarose (Roche) gel measuring 20 by 24 cm at 1.7 V/cm (40 V) overnight until the 1,150-bp fragment ofMwMarker IV (Roche) had reached a distance of 17 cm from the slots. Subsequently, the gel was blotted onto a Hybond N⫹nylon (Amersham Biosciences) membrane followed by overnight incubation of the membrane at 42°C in the presence of an IS900probe.
Preparation of the IS900probe.The IS900DNA probe was prepared by PCR amplification of a 707-bp fragment of the IS900insertion sequence specific forM. aviumsubsp.paratuberculosis.
Primers used were IS900A (5⬘ACG CCG CGG GTA GTT A) and IS900B (5⬘GGG GCG TTT GAG GTT TC).
PCRs were performed with 10 ng of chromosomal DNA of strain ATCC 19698 by using Ready to Go PCR beads (Amersham Biosciences). PCR conditions were as follows: 1 cycle of 5 min at 94°C; 30 cycles of 1 min at 94°C, 1 min at 60°C, and 1 min at 72°C; and 1 cycle of 4 min at 72°C. PCR products were purified on Qiaquick spin columns (QIAGEN) according to the manufacturer’s instructions. The probe was labeled with peroxidase (ECL DNA labeling kit; Amersham Biosciences) according to the manufacturer’s instructions.
Analysis of the RFLP patterns.Analysis of the RFLP patterns was performed by using Bionumerics version 2.0 software (Applied Maths, Sint-Martens-Latem, Belgium). Conserved bands in the IS900RFLP pattern of 8.8, 5.2, 3.0, 2.4, 2.1, and 1.6 kb were used as the internal standard for the normalization of RFLP patterns.
Selection of VNTR loci.In view of the close genetic relatedness ofM. avium
subsp.paratuberculosisand otherM. aviumcomplex strains, the recently pub-lished genomic sequence ofM. aviumstrain 104 (GenBank accession number NC_002943 [http://www.ncbi.nlm.nih.gov/sutils/genom_table.cgi]) was used to identify VNTR sequences to be tested in MLVA typing. Tandem repeats were identified by using the Tandem Repeats Finder software (1) under the default settings of the program. Tandem repeats present in more than two copies and with a sequence match of 85% or higher were compared to the nucleotide sequence ofM. aviumsubsp.paratuberculosisstrain K10 (GenBank accession number NC_002944 [http://www.ncbi.nlm.nih.gov/sutils/genom_table.cgi]).
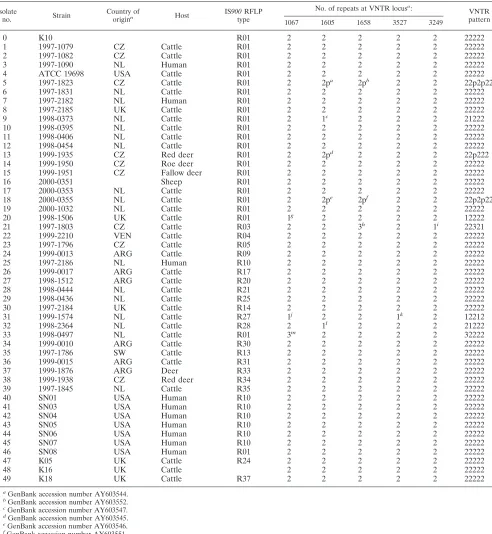
[image:2.585.133.448.84.328.2]From the total of IS900RFLP typed strains, a selection of 50 strains with variable features was made; different IS900RFLP patterns (Fig. 1) isolated from various hosts in different countries (Table 1). Primers were designed to target
FIG. 1. IS900RFLP types of the strains used for MLVA. The lanes show the patterns after normalization and analysis with Bionumerics version 2.0 (see Materials and Methods). R types are designated according to the nomenclature of the National Institute of Public Health and the Environment, Bilthoven, The Netherlands; C types are designated according to the nomenclature of Collins et al. (8) and Pavlik et al. (21)
on May 15, 2020 by guest
http://jcm.asm.org/
TABLE 1. Strains used for MLVA and results
Isolate
no. Strain
Country of
originn Host
IS900RFLP type
No. of repeats at VNTR locuso:
VNTR pattern
1067 1605 1658 3527 3249
0 K10 R01 2 2 2 2 2 22222
1 1997-1079 CZ Cattle R01 2 2 2 2 2 22222
2 1997-1082 CZ Cattle R01 2 2 2 2 2 22222
3 1997-1090 NL Human R01 2 2 2 2 2 22222
4 ATCC 19698 USA Cattle R01 2 2 2 2 2 22222
5 1997-1823 CZ Cattle R01 2 2pa 2pb 2 2 22p2p22
6 1997-1831 NL Cattle R01 2 2 2 2 2 22222
7 1997-2182 NL Human R01 2 2 2 2 2 22222
8 1997-2185 UK Cattle R01 2 2 2 2 2 22222
9 1998-0373 NL Cattle R01 2 1c 2 2 2 21222
10 1998-0395 NL Cattle R01 2 2 2 2 2 22222
11 1998-0406 NL Cattle R01 2 2 2 2 2 22222
12 1998-0454 NL Cattle R01 2 2 2 2 2 22222
13 1999-1935 CZ Red deer R01 2 2pd 2 2 2 22p222
14 1999-1950 CZ Roe deer R01 2 2 2 2 2 22222
15 1999-1951 CZ Fallow deer R01 2 2 2 2 2 22222
16 2000-0351 Sheep R01 2 2 2 2 2 22222
17 2000-0353 NL Cattle R01 2 2 2 2 2 22222
18 2000-0355 NL Cattle R01 2 2pe 2pf 2 2 22p2p22
19 2000-1032 NL Cattle R01 2 2 2 2 2 22222
20 1998-1506 UK Cattle R01 1g 2 2 2 2 12222
21 1997-1803 CZ Cattle R03 2 2 3h 2 1i 22321
22 1999-2210 VEN Cattle R04 2 2 2 2 2 22222
23 1997-1796 CZ Cattle R05 2 2 2 2 2 22222
24 1999-0013 ARG Cattle R09 2 2 2 2 2 22222
25 1997-2186 NL Human R10 2 2 2 2 2 22222
26 1999-0017 ARG Cattle R17 2 2 2 2 2 22222
27 1998-1512 ARG Cattle R20 2 2 2 2 2 22222
28 1998-0444 NL Cattle R21 2 2 2 2 2 22222
29 1998-0436 NL Cattle R25 2 2 2 2 2 22222
30 1997-2184 UK Cattle R14 2 2 2 2 2 22222
31 1999-1574 NL Cattle R27 1j 2 2 1k 2 12212
32 1998-2364 NL Cattle R28 2 1l 2 2 2 21222
33 1998-0497 NL Cattle R01 3m 2 2 2 2 32222
34 1999-0010 ARG Cattle R30 2 2 2 2 2 22222
35 1997-1786 SW Cattle R13 2 2 2 2 2 22222
36 1999-0015 ARG Cattle R31 2 2 2 2 2 22222
37 1999-1876 ARG Deer R33 2 2 2 2 2 22222
38 1999-1938 CZ Red deer R34 2 2 2 2 2 22222
39 1997-1845 NL Cattle R35 2 2 2 2 2 22222
40 SN01 USA Human R10 2 2 2 2 2 22222
41 SN03 USA Human R10 2 2 2 2 2 22222
42 SN04 USA Human R10 2 2 2 2 2 22222
43 SN05 USA Human R10 2 2 2 2 2 22222
44 SN06 USA Human R10 2 2 2 2 2 22222
45 SN07 USA Human R10 2 2 2 2 2 22222
46 SN08 USA Human R01 2 2 2 2 2 22222
47 K05 UK Cattle R24 2 2 2 2 2 22222
48 K16 UK Cattle 2 2 2 2 2 22222
49 K18 UK Cattle R37 2 2 2 2 2 22222
aGenBank accession number AY603544. bGenBank accession number AY603552. cGenBank accession number AY603547. dGenBank accession number AY603545. eGenBank accession number AY603546. fGenBank accession number AY603551. gGenBank accession number AY603549. hGenBank accession number AY603541. iGenBank accession number AY603543. jGenBank accession number AY603550. kGenBank accession number AY603542. lGenBank accession number AY603548. mGenBank accession number AY603540.
nCZ, Czech Republic; NL, The Netherlands; USA, United States; UK, United Kingdom; VEN, Venezuela; ARG, Argentina; SW, Sweden. op indicates a point mutation.
on May 15, 2020 by guest
http://jcm.asm.org/
flanking regions of the VNTRs and are listed in Table 2. The details of the amplification of the various VNTRs are listed in Table 3. PCR was performed with a Biometra T gradient thermocycler (Biometra, Go¨ttingen, Germany) by using either Ready to Go PCR beads or HotstarTaq(QIAGEN). VNTR 1067, 1605, 1658, 3527, 7661, and 9425 (Table 3) were amplified with Ready to Go PCR beads according to the instructions of the manufacturer. Due to poor results, VNTR 3249 was amplified with HotstarTaqin standard PCR buffer and Q buffer (QIAGEN) for GC-rich templates according to the instructions of the manufacturer. All PCRs were performed by using 10 ng of purified DNA. To detect differences in numbers of repeats, the PCR products were analyzed on a 1% MP agarose gel in 1⫻Tris-borate-EDTA containing 0.5g of ethidium bromide (BDH)/ml.
Sequence analysis.DNA sequencing reactions were performed with purified PCR products by using an ABI Prism Big Dye Terminator kit as described above for multilocus sequence typing analysis. Products were analyzed with an ABI 3700 capillary sequencer (Applied Biosystems). Raw data were analyzed with DNASTAR software.
Nucleotide sequence accession numbers.Sequences ofM. aviumsubsp. paratu-berculosisdetermined in this study have been deposited in the GenBank database under accession numbers AY603540, AY603541, AY603542, AY603543, AY603544, AY603545, AY603546, AY603547, AY603548, AY603549, AY603550, AY603551, and AY603552.
RESULTS
RFLP typing. Over 250 isolates were subjected to IS900
RFLP typing. Thirty different RFLP types were found. The majority (37%) was of type R01, followed by R09 (26%), R10 (8%), and R17 (7%). The isolates were gifts of available strains at a certain moment from the donating institutes; therefore,
the distribution of the different RFLP types may not reflect the natural distribution of these types.
In silico comparison of VNTR loci and flanking regions.The
whole genome sequence of M. avium strain 104 (GenBank accession number NC_2943 [available at http://www.ncbi.nlm .nih.gov/sutils/genom_table.cgi]) was analyzed for the presence of tandem repeats. In total, 376 different VNTR sequences with a length between 6 and 263 bp and with a mutual homol-ogy of 67 to 100% were found. The copy number of the repeats ranged from 2 to 56.
Sequences of VNTRs which met the following criteria were compared to the M. avium subsp. paratuberculosis K10 sequence: more than two copies, more than 18 nucleotides, and a mutual homology between 77 and 100%. A total of 47 VNTRs were selected. Other VNTRs were present in both strains but differed in the flanking regions.
Some of the VNTRs present inM. aviumstrain 104 were not present in theM. aviumsubsp.paratuberculosisK10 sequence. Finally, 27 VNTRs having two or more repeats of 20 to 70 bp in length and homology of 80 to 100% were used for a homol-ogy search of each VNTR and its flanking regions.
A comparison of 1-kb genomic regions containing each VNTR and its flanking region ofM. aviumstrain 104 andM.
aviumsubsp.paratuberculosisstrain K10 revealed mutations in
[image:4.585.45.541.81.254.2]20 loci. Thirteen loci had mutations in both the repeat and the
TABLE 2. VNTR primer sequences
Primer
no. Primer
a Annealing
temp (°C) Sequence Distance (nt)
b
1 1067 Fw 50 CGC CGC CCG CCG AAA AAG ⫺212
2 1067 Rv 50 CGG GAC ATC ACA AAT ACA GAA GAA 53
3 1605 Fw 50 CGA TGC CGC CGA AGG TTT TGG TG ⫺342
4 1605 Rv 50 ACG TGA GGA TCG GGT TGG CAG TCG 90
5 1658 Fw 50 CCC AAC CGT TCC CAA CGA GA ⫺31
6 1658 Rv 50 CCC GGG GAG CAT CAG GTC 166
7 3249 Fw 55 GCA GCA GGA CGA TCA GGC C ⫺84
8 3249 Rv 55 CGC GAG TTC GGT GCC GTG A 155
9 3527 Fw 53 GGC GCT CGC AGG AAA CCA AC ⫺125
10 3527 Rv 53 ACG GCC TCA GCT CCC AGT AG 33
11 2495 Fw 58 GCA CCC CCG GCA GCG ACT CCT CTT ⫺184
12 2495 Rv 58 GCA ACC CCG GCG TCG TCC CCA ATC 141
13 7661 Fw 60 TCG TGC TGT CCA TCG CGG GGT AGG ⫺33
14 7661 Rv 60 GCA TCG CCA GCA CAT AGC 124
15 9425 Fw 63 AGG TGG CCA GGC GGG TCA TCT TCC ⫺80
16 9425 Rv 63 CAG AAG GCG ACG GAG GAC TAC ATC 46
aFw, forward primer. Rv, reverse primer. bDistance is from the 5⬘end to the nearest repeat.
TABLE 3. PCR protocols for MLVA
VNTR Conditions for PCR step no.:
1 (1 cycle) 2 (30 cycles) 3 (1 cycle)
1067, 1605, 1658 95°C, 5 min 94°C, 1 min; 50°C, 1 min; 72°C, 1 min 72°C, 4 min
2495 95°C, 5 min 94°C, 1 min; 58°C, 1 min; 72°C, 1 min 72°C, 4 min
7661 95°C, 5 min 94°C, 1 min; 60°C, 1 min; 72°C, 1 min 72°C, 4 min
9425 95°C, 5 min 94°C, 1 min; 63°C, 1 min; 72°C, 1 min 72°C, 4 min
3527 95°C, 5 min 94°C, 1 min; 53°C, 1 min; 72°C, 1 min 72°C, 4 min
3249 95°C, 15 mina 94°C, 1 min; 55°C, 1 min; 72°C, 1 min 72°C, 4 min
aReactions with HotstarTaqrequire initial activation ofTaqpolymerase.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.585.42.547.624.716.2]flanking regions. Seven isolates had mutations only in the flanking regions.
The number of point mutations in the VNTR flanking re-gions varied from 2 to 13. The differences observed comprised inserts, gaps, and point mutations. The number of mutations found in repeat regions ranged from 0 to 4 nucleotides. Fur-thermore, deletions of whole repeats were observed.
Thirteen out of 20 VNTR loci inM. aviumsubsp.
paratuber-culosisstrain K10 had fewer repeats than their corresponding
locus inM. aviumstrain 104. Six loci had an equal number and one had an increased number of repeats.
Variation in repeat number amongM. aviumsubsp.
paratu-berculosisisolates. As a result of the in silico analysis, the 20 VNTR loci showing mutations compared to the M. avium strain 104 were tested. Polymorphism in these VNTR loci was determined in 49M. aviumsubsp.paratuberculosisstrains (Ta-ble 1), 22 of RFLP type R01, 7 of RFLP type R10, 19 of other RFLP types, and 1 of an unknown type.
PCR on four loci yielded either no product or multiple nonspecific products. These loci were excluded from further study. Eleven loci gave no polymorphism in the number of repeats. Four loci showed two different VNTR types, and one locus revealed three VNTR types. Based on analysis of the five VNTR loci in 49M. aviumsubsp.paratuberculosisisolates, six different MLVA types were found (Table 1).
DNA sequence analysis of variable VNTR loci inM. avium
subsp.paratuberculosisstrains.All VNTR loci with variation in
the number of repeats were subjected to DNA sequence anal-ysis. In addition, three VNTR loci (2495, 7661, and 9425) without variation in the number of repeats were analyzed to detect possible nucleotide polymorphisms.
In the loci 1605 and 1658, point mutations betweenM. avium subsp.paratuberculosisstrains were observed in the genomic regions flanking the VNTRs. In three strains, a C-T mutation at position 119 was observed in VNTR 1605. The location of this mutation is outside the repeat sequence. In two strains, a C-T mutation was observed at position 74 of VNTR 1658, and this mutation was positioned inside the second repeat. The DNA sequence of the three loci 2495, 7661, and 9425 showed no variation in the number of tandem repeats analyzed nor did they reveal any mutations at the single-nucleotide level.
In conclusion, MVLA typing of 49M. aviumsubsp.
paratu-berculosis isolates yielded six different genotypes, and DNA
sequence analysis of the VNTR loci resulted in two additional alleles.
Comparison of IS900RFLP typing and MLVA-associated
variation.The 22 strains of RFLP type R01 could be
subdi-vided into six different combined VNTR types, providing a subdivision of the largest group of IS900 RFLP types. Seven strains of RFLP type R10 have an MLVA type identical to 17 other different RFLP types. No association was found between the MLVA types of the M. avium subsp. paratuberculosis strains and the animal source or the country of isolation. All human isolates revealed the same MLVA type, 22222, as found for the majority of the cattle strains. This finding indicates that these isolates are genetically closely related to the most com-mon and widespread cattle isolates.
Diversity index.Diversity indices of RFLP and MLVA
typ-ing were calculated accordtyp-ing to the method of Hunter and Gaston (15).The index ranges between 0 and 1. The closer the
diversity index is to 1, the higher the discriminatory power of the method. The RFLP index is based on 49 strains of known IS900RFLP type (Table 1). The MLVA index is based on 50 strains of known MLVA type. The indices are 0.448 and 0.316, respectively.
DISCUSSION
The detection of four different VNTR types and two point mutations in the VNTR flanking regions among 24M. avium subsp.paratuberculosisisolates of the R01 RFLP type analyzed is promising for VNTR typing as a tool to study the epidemi-ology ofM. aviumsubsp.paratuberculosis. Further analysis of other VNTRs in the genome ofM. aviumsubsp.
paratubercu-losis may yield an epidemiological tool with an even higher
resolution, enabling a clear interpretation of comparative DNA fingerprinting of M. aviumsubsp. paratuberculosis iso-lates from animal and human sources.
The C-T mutation in VNTR 1605 causes the loss of an HaeII restriction site, which can be used for the detection of this mutation.
The search for VNTRs inM. aviumsubsp.paratuberculosis was based on the genome sequence ofM. aviumstrain 104. Since the complete nucleotide sequence of M. aviumsubsp.
paratuberculosisstrain K10 has recently become available for
BLAST searches, it is possible to compare VNTRs present in
M. avium strain 104 with the genome of M. avium subsp.
paratuberculosis. It is conceivable that other VNTRs will be
disclosed which also show polymorphism among M. avium subsp.paratuberculosisstrains and can be used in MLVA typ-ing.
Evaluation of RFLP versus MLVA shows that on the one hand, the group of R01 could be successfully divided in sub-groups, but on the other hand, 17 different RFLP types have the same MLVA type. More strains of each RFLP type need to be tested by MLVA to establish the absolute value of MLVA typing compared to the different RFLP types. Evaluation of RFLP versus MLVA typing by calculating the diversity index gives an idea about the discriminative value of these methods. The results suggest that RFLP is more discriminative than MLVA (0.448 and 0.316, respectively), but one has to keep in mind that the strain panel was based on RFLP types and as a result of that favors RFLP over MLVA in the number of different types assigned to it. Once the calculation was based upon the group R01 RFLP type isolates, among which six different MLVA types were found, the MLVA method is much more discriminative. The index is more reliable for large rep-resentative collections of strains (15). In this study, there is neither a representative collection nor a large number of strains, so limiting conditions are not applicable. Therefore, the reliability of the indices is low.
So far, only a limited number ofM. aviumsubsp.
paratuber-culosis isolates from humans is available for typing. In this
study, 10 M. aviumsubsp. paratuberculosisisolates from hu-mans have been subjected to IS900RFLP typing. This typing revealed only cattle types: three isolates belong to type R01 and seven belong to type R10. However, in The Netherlands, only 10% of the cattle isolates are of the R10 (C5) type (P. Overduin and D. van Soolingen, unpublished data), and in an international study, only 0.5% of the cattle isolates were
on May 15, 2020 by guest
http://jcm.asm.org/
signed to the R10 type (22). In view of this finding, it is re-markable that six out of sevenM. aviumsubsp.paratuberculosis isolates originating from the United States have RFLP type R10. One could hypothesize that the high percentage of R10 (C5) strains found in humans indicates that theseM. avium subsp.paratuberculosis strains are more capable of infecting humans or that there is a (unknown) reservoir ofM. avium subsp.paratuberculosisdifferent from cattle. More isolates are needed for further study of this question.
All isolates from humans were of the most common MLVA type (22222). In addition to IS900RFLP typing, this provides evidence that the human M. avium subsp. paratuberculosis strains are identical to a subpopulation of the strains found in cattle. This finding suggests that humans can be infected with
M. aviumsubsp.paratuberculosisstrains from cattle.
Unfortu-nately, cultivation of M. avium subsp. paratuberculosis from human tissue is still very difficult. Although modern liquid cultures produce higher yields than solid media (23), the num-ber of human isolates available for typing is still very limited (2). Therefore, until more suitable cultivation methods have become available and the nature of noncultivatableM. avium subsp.paratuberculosisbacteria has been disclosed, large epi-demiological studies on human isolates are future prospects.
Spoligotyping is frequently used as an alternative for, or in addition to, RFLP typing ofM. tuberculosis. We have explored the use of spoligotyping for discrimination ofM. aviumsubsp.
paratuberculosisstrains (P. Overduin, A. Herrewegh, and D.
van Soolingen, unpublished data). As a model, we have used the genome sequence ofM. aviumstrain 104 which has a direct repeat region containing a 27-bp repeat and 12 spacers. De-tailed investigation of other M. avium strains with a DNA probe covering a large part of the direct repeat region ofM.
aviumstrain 104 revealed that only 2 out of 20M. aviumstrains
had anM. aviumstrain 104 homologous direct repeat region. Furthermore, study of the complete genome sequence ofM.
aviumsubsp.paratuberculosisstrain K10 revealed that neither
this direct repeat region nor the spacers found inM. avium strain 104 are present in M. avium subsp. paratuberculosis strain K10. These results indicate that the direct repeat region found inM. aviumstrain 104 is not common in otherM. avium complex strains. It remains unclear whether there is a direct repeat region present inM. aviumsubsp.paratuberculosisat all. In this study, we tested a number of VNTRs for the presence of polymorphisms in the conserved mycobacterial subspecies
M. avium subsp. paratuberculosis. Five out of eight studied
VNTRs showed polymorphism in the number of repeats. Se-quencing revealed point mutations in two VNTRs. Both poly-morphisms divided the major R01 RFLP group into several subgroups, demonstrating that VNTR typing could be a useful (additional) tool for typingM. aviumsubsp.paratuberculosis.
ACKNOWLEDGMENTS
We thank Ivo Pavlik, Douwe Bakker, Cees Kalis, Mariso Romano, Jacobus de Waard, and John Hermon-Taylor for kindly donatingM. aviumsubsp.paratuberculosisisolates and DNA.
M. aviumstrain 104 was sequenced by The Institute for Genomic Research, Rockville, Md.M. aviumsubsp.paratuberculosisstrain K10 was sequenced at the Department of Microbiology and Biomedical Genomics Center, University of Minnesota, St. Paul, Minn., by L. Li, J. Bannantine, Q. Zhang, A. Amonsin, D. Alt, and V. Kapur.
REFERENCES
1.Benson, G.1999. Tandem repeats finder: a program to analyze DNA se-quences. Nucleic Acids Res.27:573–580.
2.Bull, T. J., E. J. McMinn, K. Sidi-Boumedine, A. Skull, D. Durkin, P. Neild, G. Rhodes, R. Pickup, and J. Hermon-Taylor.2003. Detection and verifica-tion of Mycobacterium aviumsubsp.paratuberculosisin fresh ileocolonic mucosal biopsy specimens from individuals with and without Crohn’s dis-ease. J. Clin. Microbiol.41:2915–2923.
3.Bull, T. J., K. Sidi-Boumedine, E. J. McMinn, K. Stevenson, R. Pickup, and J. Hermon-Taylor. 2003. Mycobacterial interspersed repetitive units (MIRU) differentiateMycobacterium aviumsubspeciesparatuberculosisfrom other species of theMycobacterium aviumcomplex. Mol. Cell. Probes17:
157–164.
4.Cellier, C., H. De Beenhouwer, A. Berger, C. Penna, F. Carbonnel, R. Parc, P. H. Cugnenc, Y. Le Quintrec, J. P. Gendre, J. P. Barbier, and F. Portaels.
1998.Mycobacterium paratuberculosisandMycobacterium aviumsubsp. sil-vaticumDNA cannot be detected by PCR in Crohn’s disease tissue. Gastro-enterol. Clin. Biol.22:675–678.
5.Chiba, M., T. Fukushima, Y. Horie, M. Iizuka, and O. Masamune.1998. No
Mycobacterium paratuberculosisdetected in intestinal tissue, including Pey-er’s patches and lymph follicles, of Crohn’s disease. J. Gastroenterol.33:
482–487.
6.Chiodini, R. J., H. J. van Kruiningen, W. R. Thayer, R. S. Merkal, and J. A. Coutu.1984. Possible role of mycobacteria in inflammatory bowel disease. I. An unclassified Mycobacterium species isolated from patients with Crohn’s disease. Dig. Dis. Sci.29:1073–1079.
7.Clarke, C. J.1997. The pathology and pathogenesis of paratuberculosis in ruminants and other species. J. Comp. Pathol.116:217–261.
8.Collins, D. M., D. M. Gabric, and G. W. de Lisle.1990. Identification of two groups ofMycobacterium paratuberculosisstrains by restriction endonuclease analysis and DNA hybridization. J. Clin. Microbiol.28:1591–1596. 9.Cowan, L. S., L. Mosher, L. Diem, J. P. Massey, and J. T. Crawford.2002.
Variable-number tandem repeat typing ofMycobacterium tuberculosis iso-lates with low copy numbers of IS6110by using mycobacterial interspersed repetitive units. J. Clin. Microbiol.40:1592–1602.
10.DelPrete, R., M. Quaranta, A. Lippolis, V. Giannuzzi, A. Mosca, E. Jirillo, and G. Miragliotta.1998. Detection ofMycobacterium paratuberculosisin stool samples of patients with inflammatory bowel disease by IS900-based PCR and colorimetric detection of amplified DNA. J. Microbiol. Methods
33:105–114.
11.Gitnick, G., J. Collins, B. Beaman, D. Brooks, M. Arthur, T. Imaeda, and M. Palieschesky.1989. Preliminary report on isolation of mycobacteria from patients with Crohn’s disease. Dig. Dis. Sci.34:925–932.
12.Grant, I. R., H. J. Ball, and M. T. Rowe.1999. Effect of higher pasteurization temperatures, and longer holding times at 72°C, on the inactivation of My-cobacterium paratuberculosisin milk. Lett. Appl. Microbiol.28:461–465. 13.Grant, I. R., H. J. Ball, and M. T. Rowe.2002. Incidence ofMycobacterium
paratuberculosisin bulk raw and commercially pasteurized cows’ milk from approved dairy processing establishments in the United Kingdom. Appl. Environ. Microbiol.68:2428–2435.
14.Haagsma, J., C. J. J. Mulder, A. Eger, and G. N. J. Tytgat.1992. Mycobac-terial isolates from Crohn’s disease in The Netherlands, p. 16–21.InC. J. J. Mulder and G. N. J. Tytgat (ed.), Is Crohn’s disease a mycobacterial disease. Kluwer Academic Publishers, Dordrecht, The Netherlands.
15.Hunter, P. R., and M. A. Gaston.1988. Numerical index of the discrimina-tory ability of typing systems: an application of Simpson’s index of diversity. J. Clin. Microbiol.26:2465–2466.
16.Karlinger, K., T. Gyorke, E. Mako, A. Mester, and Z. Tarjan.2000. The epidemiology and the pathogenesis of inflammatory bowel disease. Eur. J. Radiol.35:154–167.
17.Lee, A. S. G., L. L. H. Tang, I. H. K. Lim, R. Bellamy, and S.-Y. Wong.2002. Discrimination of single-copy IS6110DNA fingerprints ofMycobacterium tuberculosisisolates by high-resolution minisatellite-based typing. J. Clin. Microbiol.40:657–659.
18.Marsh, I., R. Whittington, and D. Cousins.1999. PCR-restriction endonu-clease analysis for identification and strain typing ofMycobacterium avium
subsp.paratuberculosisandMycobacterium avium subsp.avium based on polymorphisms in IS1311. Mol. Cell. Probes13:115–126.
19.McFadden, J. J., P. D. Butcher, R. Chiodini, and T. J. Hermon.1987. Crohn’s disease-isolated mycobacteria are identical toMycobacterium para-tuberculosis, as determined by DNA probes that distinguish between myco-bacterial species. J. Clin. Microbiol.25:796–801.
20.Millar, D., J. Ford, J. Sanderson, S. Withey, M. Tizard, T. Doran, and J. Hermon-Taylor.1996. IS900PCR to detectMycobacterium paratubercu-losisin retail supplies of whole pasteurized cows’ milk in England and Wales. Appl. Environ. Microbiol.62:3446–3452.
21.Pavlik, I., L. Bejckova, M. Pavlas, Z. Rozsypalova, and S. Koskova.1995. Characterization by restriction endonuclease analysis and DNA hybridiza-tion using IS900of bovine, ovine, caprine and human dependent strains of
Mycobacterium paratuberculosisisolated in various localities. Vet. Microbiol.
45:311–318.
on May 15, 2020 by guest
http://jcm.asm.org/
22.Pavlik, I., A. Horvathova, L. Dvorska, J. Bartl, P. Svastova, R. du Maine, and I. Rychlik.1999. Standardisation of restriction fragment length polymor-phism analysis forMycobacterium aviumsubspeciesparatuberculosis. J. Mi-crobiol. Methods38:155–167.
23.Schwartz, D., I. Shafran, C. Romero, C. Piromalli, J. Biggerstaff, N. Naser, W. Chamberlain, and S. A. Naser. 2000. Use of short-term culture for identification ofMycobacterium aviumsubsp.paratuberculosisin tissue from Crohn’s disease patients. Clin. Microbiol. Infect.6:303–307.
24.Stevenson, K., V. M. Hughes, L. de Juan, N. F. Inglis, F. Wright, and J. M. Sharp.2002. Molecular characterization of pigmented and nonpigmented isolates ofMycobacterium aviumsubsp.paratuberculosis. J. Clin. Microbiol.
40:1798–1804.
25.Supply, P., J. Magdalena, S. Himpens, and C. Locht.1997. Identification of novel intergenic repetitive units in a mycobacterial two-component system operon. Mol. Microbiol.26:991–1003.
26.Supply, P., E. Mazars, S. Lesjean, V. Vincent, B. Gicquel, and C. Locht.2000. Variable human minisatellite-like regions in theMycobacterium tuberculosis
genome. Mol. Microbiol.36:762–771.
27.Thompson, D. E.1994. The role of mycobacteria in Crohn’s disease. J. Med. Microbiol.41:74–94.
28.van Soolingen, D., P. E. de Haas, P. W. Hermans, and J. D. van Embden.
1994. DNA fingerprinting ofMycobacterium tuberculosis. Methods Enzymol.
235:196–205.
29.Whipple, D. L., D. R. Callihan, and J. L. Jarnagin.1991. Cultivation of
Mycobacterium paratuberculosisfrom bovine fecal specimens and a suggested standardized procedure. J. Vet. Diagn. Investig.3:368–373.
30.Whittington, R. J., A. F. Hope, D. J. Marshall, C. A. Taragel, and I. Marsh.
2000. Molecular epidemiology ofMycobacterium aviumsubsp. paratubercu-losis: IS900restriction fragment length polymorphism and IS1311 polymor-phism analyses of isolates from animals and a human in Australia. J. Clin. Microbiol.38:3240–3248.
31.Whittington, R. J., C. A. Taragel, S. Ottaway, I. Marsh, J. Seaman, and V. Fridriksdottir.2001. Molecular epidemiological confirmation and circum-stances of occurrence of sheep (S) strains ofMycobacterium aviumsubsp.
paratuberculosisin cases of paratuberculosis in cattle in Australia and sheep and cattle in Iceland. Vet. Microbiol.79:311–322.