Reproductive and morphometric characteristics of wild boar (Sus scrofa) in the Czech Republic
8
0
0
Full text
Figure

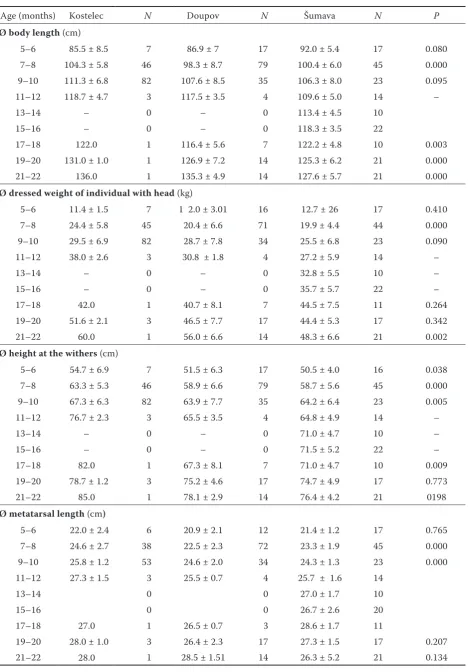
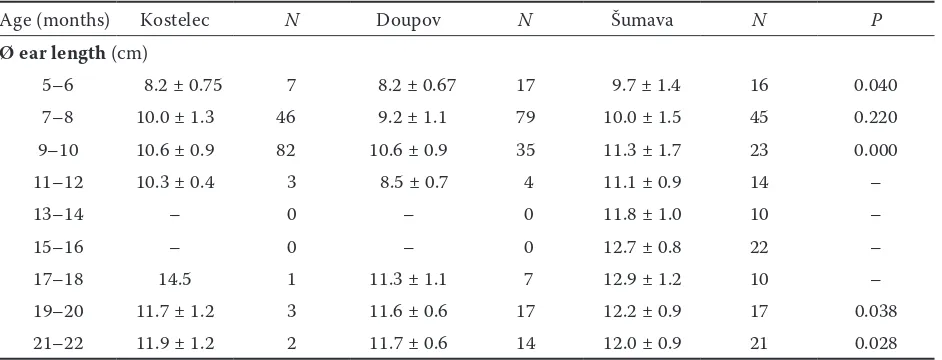

+2

Related documents