Invasion success of a scarab beetle within its native range: Host range expansion versus host-shift
Full text
Figure

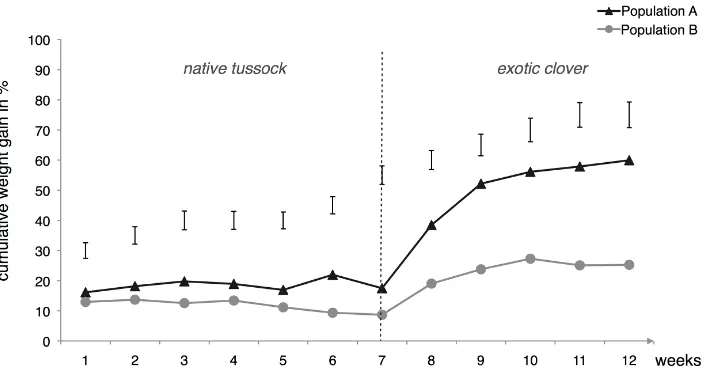

Related documents
b All cell wall-de fi cient cells were able to form mycelial colonies on MYM medium lacking high levels of osmolytes.. Unlike the majority of colonies derived from S-cells,
In China, an increasing number of universities expect to improve the quality of thesis (dissertation) by the application of blind review system and they regard it as a way to exclude
Specifically, we examine teacher education governance, policy reform, and political context from 2000 to 2010, across four Canadian provinces: Alberta, British Columbia, Manitoba,
Saks Fifth Avenue also is implementing an omni-channel approach as it builds its next-generation retail environment: Saks is using solutions from Oracle® to enable a vision of
F The African-American model also controls for child sex, marital status, family income, child health status, health insurance, source of well child care, mother’s education
Analysis of data regarding medical staff shift changes revealed that 64 % of the patients been observed experi- enced a shift change of nurses during their stay.. The aver- age
The export-sales ratios for the sub- sample of UK firms exhibit the theoretically expected relationship: The export intensity increases with firm size, but firms in the largest
At the struc- ture level, this study examined performance criteria in national higher ed- ucation system rankings (e.g., Universitas 21-U21) and university rankings (e.g., Shanghai
