Which trees should be removed in thinning treatments?
12
0
0
Full text
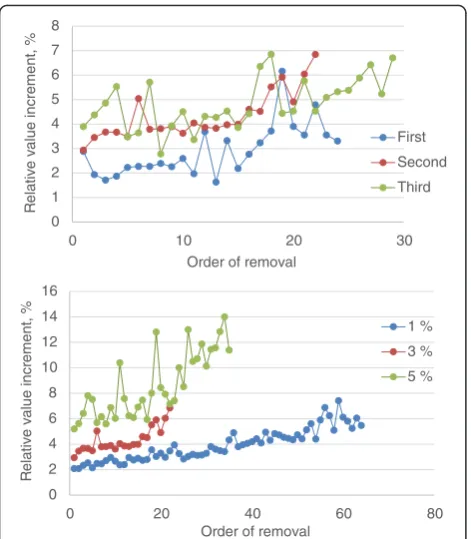
Figure
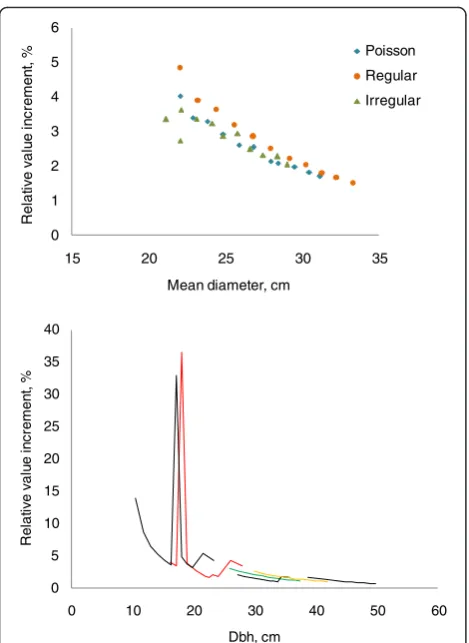
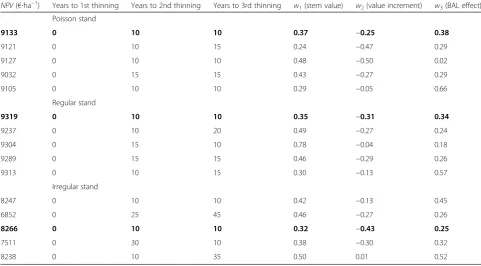
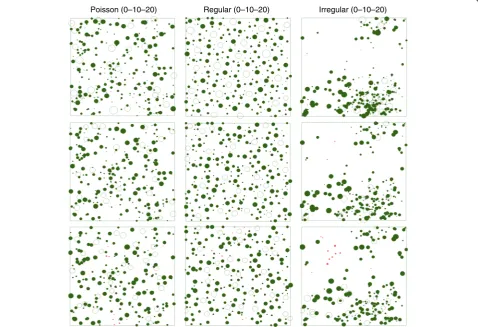
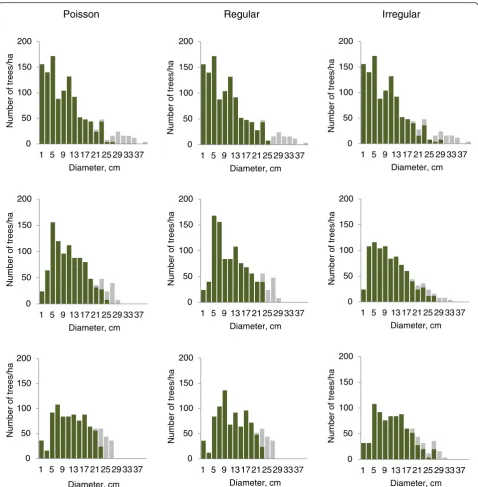
+7

Related documents