Upstream and Downstream Regulation of Asexual Development in Aspergillus fumigatus
Full text
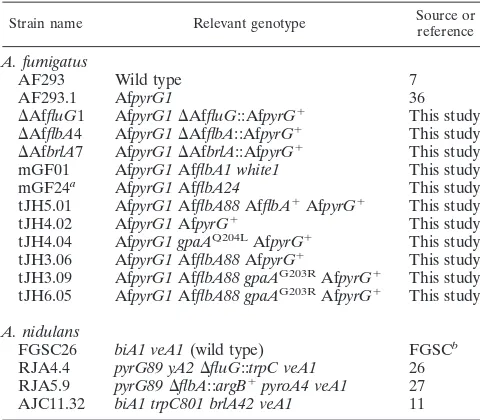
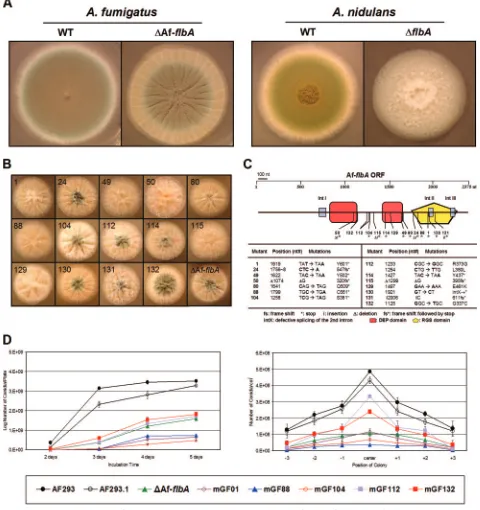
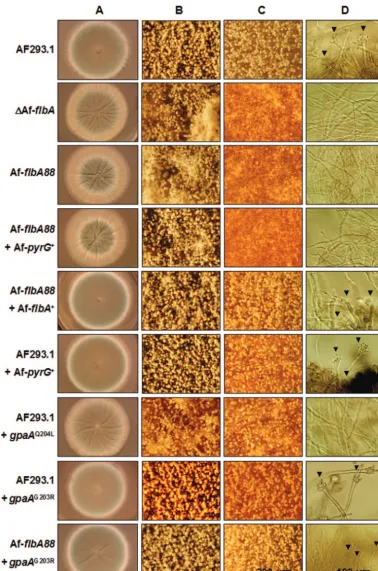
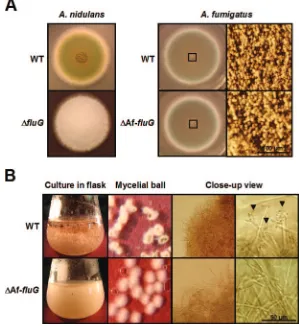
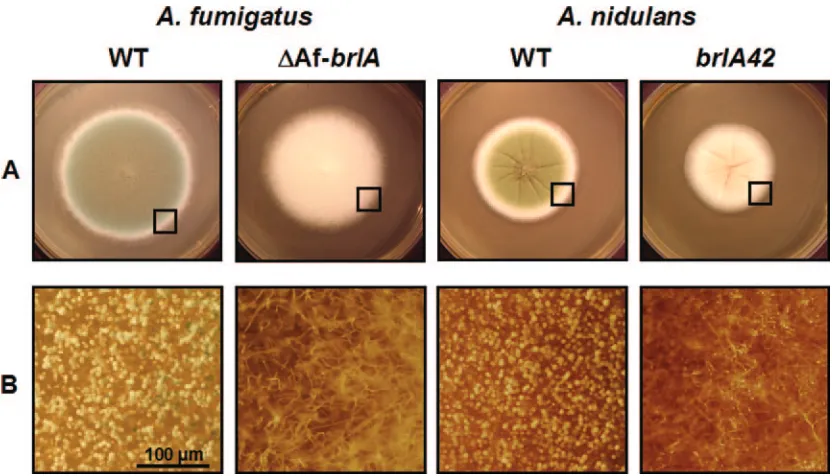
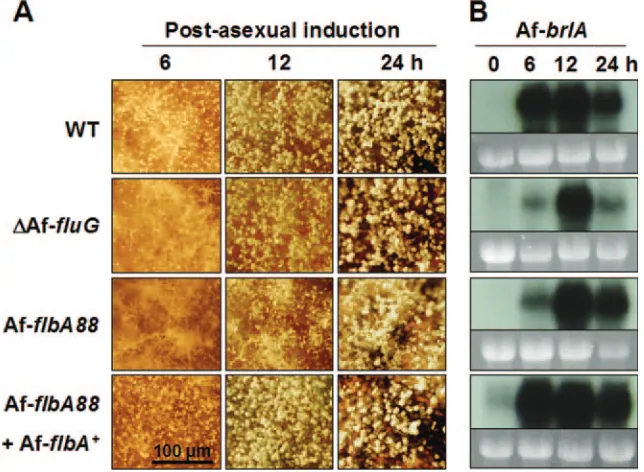
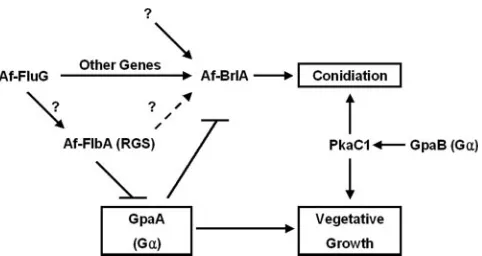
Figure
Related documents
Timberset, containing high-modulus ceramic particles, has the highest storage modulus E’, peak loss modulus E‖ and glass transition temperature Tg, followed by Albipox,
Whenever user Logs into system using portal, application layer transparently passes request to transaction manager which first authenticates and authorizes user, then
Correlation between age and different resistant organisms revealed that proteus is statistically significant present in pressure sores of younger age group 60-80 y on
An alcohol sensor is connected to the Arduino board sense alcohol usage of driver.MATLAB mobile an android application gives the location details, speed of vehicle to MATLAB in
Schematic view of the BM@N experimental setup: SiMS – Silicon Micro-strip Sensors, GEM – Gas Electron Multipliers, CSC – Cathode Strip Chamber (CSC), mRPC – Multi-gap Resistive
In the present study, the abdominal cavity ESTBS allowed various types of training and implementation of progressive complexity tasks, since the medical students, surgery residents
Connected health devices In addition to these educa- tion software applications, connected health devices form part of the telemonitoring system: connected blood glu- cose monitor
ABM-MNCs: autologous bone marrow-derived mononuclear cells; T2DM: type 2 diabetes mellitus; HbA1c: glycated hemoglobin; AUC: area under curve; HOMA-IR: homeostatic model assessment
