Zebrafish as a model for leukemia and other hematopoietic disorders
10
0
0
Full text
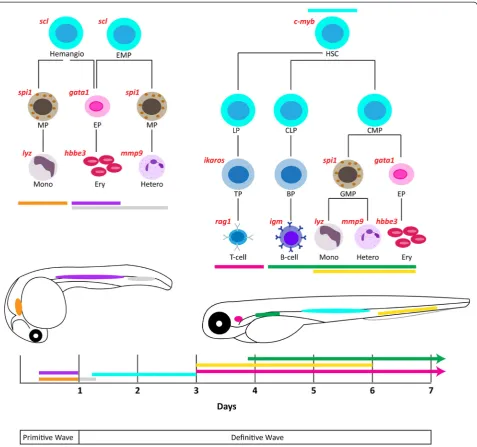
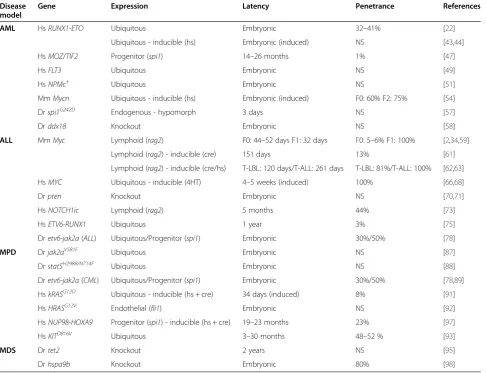
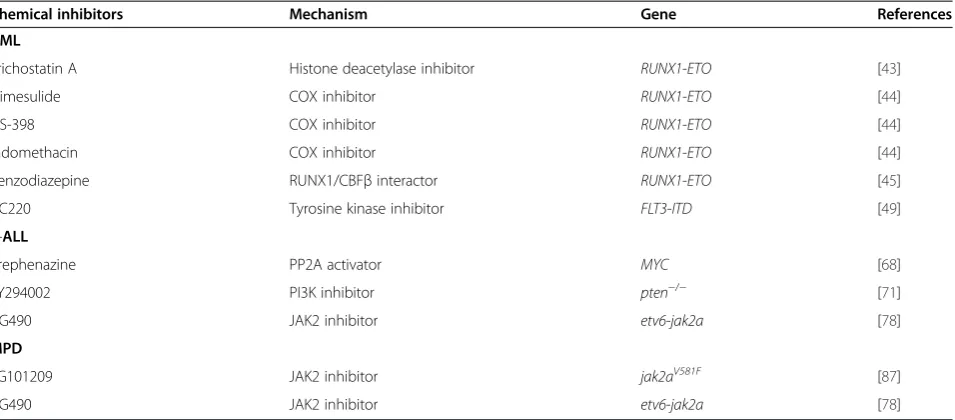
Figure
Related documents