ANATOMICAL IMPLICATIONS
Nitina Ahuja1
, Daizy R. Batish2
and Harminder Pal Singh3 1,2
Department of Botany, Panjab University, Chandigarh, 3
Department of Environment Studies, Panjab University, Chandigarh – 160014, India
Abstract
The present study was conducted to evaluate the phytotoxic effect of eugenol, a monoterpenoid alcohol, on some crop species and to determine its anatomical implications. For this study, three crops viz., Zea mays, Phaseolus aureus and Oryza sativa were selected. A laboratory bioassay was carried out to assess the phytotoxicity of eugenol on the crop species in a dose-dependent manner. Eugenol inhibited the early growth of the crop species determined by its effect on the radicle and plumule length. The radicle and plumule length were found to decrease with increase in concentration of eugenol in all the three crop species. Significant inhibition was observed at higher dose of eugenol treatment. The radicle inhibition was more pronounced as compared to the plumule. Zea mays was found to be the most sensitive among the test plants. Further, significant alterations in the internal structure of roots of Zea mays as seen in the epidermal, cortical and vascular regions were observed due to eugenol treatment. Therefore, the phytotoxicity of eugenol towards crops is a significant step to determine its potential against weed species for its possible use as a bio-herbicide in the agro-ecosystems.
Keywords: Eugenol, Phytotoxicity, Bioherbicide
I. INTRODUCTION
Plants possess a vast repository of natural compounds/ essential oils which play a vital role in various ecological interactions. The aromatic essential oils released from plants function as pollinator attractants [1], are involved in plant-mediated interactions with other plants, microbes and animals [2] and even play a role in plant defence against microbes, herbivores and pathogens [3]. Of late, the essential oils/ monoterpenes have found a great significance in terms of their high phytotoxicity towards a number of other plants [4,5,6,7,8,9]. Release of essential oils and their constituent monoterpenes from aromatic plants affects the growth of other plants in their vicinity. There are a number of reports which highlight the importance of these essential oils as allelopathic agents [10,11,12,13,14,15]. In view of the above studies, it is worthwhile to assess the phytotoxicity of other monoterpenoids/constituents of essential oils so as to identify potential lead chemicals for synthesis of new herbicides. Among constituents of essential oils, eugenol, a monoterpenoid alcohol is a major constituent of clove (Eugenia caryophyllata [L.] Merrill and Perry) oil, nutmeg (Myristica fragrans
Houtt.), cinnamon (Cinnamomum zeylanicum Blume.) and bay leaf (Laurus nobilis L.). It is known
to possess phytotoxicity against a number of plants [10,16,17,18,19] and thus has potential to be used as a bioherbicide [10]. Thus, the present study was planned to assess the phytotoxicity of eugenol on the early growth of some plants (Zea mays, Oryza sativa and Phaseolus aureus) and to determine its possible effect on the root internal structure.
II. MATERIALS AND METHODS 2.1. Materials
Volume 03, Issue 7, [July- 2017] e-ISSN: 2394-5532, p-ISSN: 2394-823X
were procured from Punjab Agricultural University, Ludhiana. The seeds were surface sterilized with sodium hypochlorite (0.1%, w/v) followed by washing with distilled water three times. The seeds were then pre-imbibed in distilled water for 16-18 h prior to germination.
2.2. Methodology
2.2.1. Growth studies under laboratory conditions
Studies were undertaken to explore the effect of eugenol on early growth and development of the three test plants viz. Zea mays, Oryza sativa and Phaseolus aureus under laboratory conditions. The pre-imbibed seeds, ten of each plant were uniformly placed in Petri dishes (6 inches in diameter) lined with a thin moist layer of absorbant cotton wad overlined with Whatman no. 1 filter paper. Different concentrations of eugenol i.e. 0.50, 1.0, 2.0, 5.0, 10.0, 20.0, 40.0nl/cc were applied on the inner side of Petri dish lid. The Petri dishes were immediately sealed with double layer of adhesive tape.
The Petri dishes in similar set up but without treatment of eugenol, served as control. For each treatment, five replicates were maintained in a growth chamber at 25±2˚C temperature, 16/8 light/dark photoperiod and a relative humidity of 75±2% and photon flux density of 150 µmol m-2s-1. After five days (120h), length of the seedlings was measured with the help of a scale.
2.2.2. Root Anatomy: Staining and Mounting
For anatomical studies, the roots from seedlings of Zea mays (selected due to its maximum sensitivity towards eugenol; clear root morphology) raised in Petri dishes (as described earlier) were used. The roots were chopped on the 5th day and fixed in fixative (in the ratio ethyl alcohol: pure water: acetic acid: formalin; 45 : 45 : 5 : 5) for 24 h. After 24 h, the sections of root were cut with hand using fresh surgical blade and subjected to permanent staining and mounting. Prior to permanent staining, all sections were transferred to distilled water.
The thin fine sections were transferred to lactic acid and warmed for 2-3 min. The sections were then dipped in aniline blue stain and washed in lactic acid to remove the excess stain.
The stained sections were mounted in glycerine. Each section was transferred from lactic acid to a drop of glycerine loaded on a clean glass slide. The section was covered with a cover slip carefully to avoid any air bubble and observed under light microscope.
The finely stained sections of various concentrations were photographed using Digital Phase Contrast Microscope (Leica DM - 3000).
2.2.3. Statistical Analysis
For each experiment, five replicates were maintained in a completely randomized block design. The data of the experiments was expressed as mean values and subjected to one-way
Analysis of Variance (ANOVA) followed by separation of means applying a post hoc Tukey’s test
using software programme SPSS (version 10).
III. RESULTS AND DISCUSSION
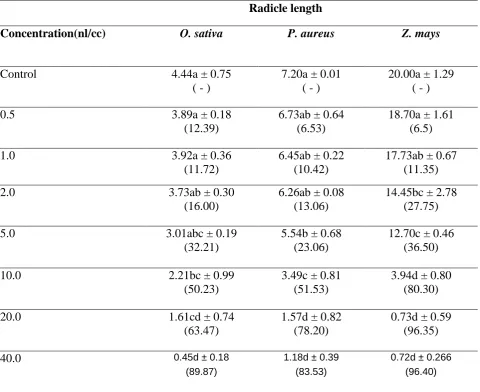
In all the three plants, the radicle length was observed to decrease gradually with an increase in the concentration of eugenol on comparison with control.
In O. sativa, a significant decrease by ~50% with respect to control was observed at 10 nl/cc concentration which further reduced to ~63% and 90%, at 20 and 40 nl/cc eugenol respectively (Table 1).
Volume 03, Issue 7, [July- 2017] e-ISSN: 2394-5532, p-ISSN: 2394-823X
Table 1. Effect of different concentrations of eugenol on the radicle length of Oryza sativa, Phaseolus aureus and Zea mays
Values within parenthesis indicate % decrease over control
Different alphabets indicate significant difference at P ≤ 0.05 applying Tukey’s test
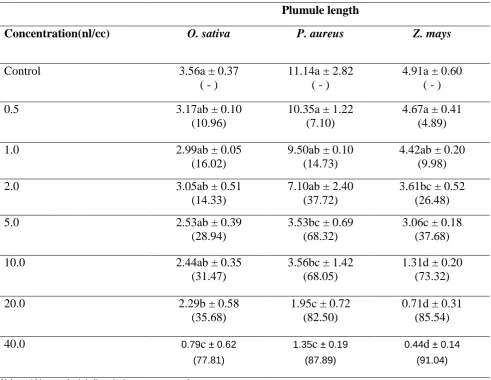
A similar trend of decrease was also observed in the plumule length. A decline in the plumule length with respect to control was observed in all the three plants in a dose-response manner (Table 2). In case of O. sativa, a gradual decrease in the plumule length was measured upto 10 nl/cc concentration. Thereafter, a significant decrease was recorded at higher concentrations of 20 nl/cc (~36%) and 40 nl/cc (~78%) (Table 2).In P. aureus, a significant decrease of ~ 68% in the plumule length was observed at 5 nl/cc concentration. At the highest concentration of 40 nl/cc the plumule length declined by ~88%.Further, in case of Z. mays, a significant inhibition in the plumule length was observed at 2 nl/cc eugenol. A further reduction of ~73%, 86% and 91% was recorded in the plumule length at 10, 20 and 40 nl/cc of eugenol (Table 2).
The results clearly indicate that eugenol inhibits the early growth of all the three test plants viz.Oryza sativa, Phaseolus aureus and Zea mays. In general, the inhibition was dose-responsive i.e. the radicle as well as plumule length declined with an increase in the concentration of eugenol.
However, maximum inhibition was observed in case of Z. mays for both radicle and plumule length.
This observation is not new as there are several reports in literature which indicate the growth retardatory effects of essential oils and their chemical constituents, especially the monoterpenes [8,15,20,21,22].
Radicle length
Concentration(nl/cc) O. sativa P. aureus Z. mays
Control 4.44a ± 0.75
( - )
7.20a ± 0.01 ( - )
20.00a ± 1.29 ( - )
0.5 3.89a ± 0.18
(12.39)
6.73ab ± 0.64 (6.53)
18.70a ± 1.61 (6.5)
1.0 3.92a ± 0.36
(11.72)
6.45ab ± 0.22 (10.42)
17.73ab ± 0.67 (11.35)
2.0 3.73ab ± 0.30
(16.00)
6.26ab ± 0.08 (13.06)
14.45bc ± 2.78 (27.75)
5.0 3.01abc ± 0.19
(32.21)
5.54b ± 0.68 (23.06)
12.70c ± 0.46 (36.50)
10.0 2.21bc ± 0.99
(50.23)
3.49c ± 0.81 (51.53)
3.94d ± 0.80 (80.30)
20.0 1.61cd ± 0.74
(63.47)
1.57d ± 0.82 (78.20)
0.73d ± 0.59 (96.35)
40.0 0.45d ± 0.18
(89.87)
1.18d ± 0.39 (83.53)
Volume 03, Issue 7, [July- 2017] e-ISSN: 2394-5532, p-ISSN: 2394-823X
Table 2. Effect of different concentrations of eugenol on the radicle length of Oryza sativa, Phaseolus aureus and Zea mays
Values within parenthesis indicate % decrease over control
Different alphabets indicate significant difference at P ≤ 0.05 applying Tukey’s test
[23] assessed the activity of twenty seven monoterpenes against seed germination and early
radicle growth of Raphanus sativus L. (radish) and Lepidium sativum L. (garden cress). They
ascertained a dose-dependent inhibitory activity of all the monoterpenes to a differential extent in both the test plants. Carvone was observed to be inhibitory to the radicle elongation of both the test
weeds at all concentrations. In the present study, it was also found that eugenol caused greater
retardation in the radicle length than the plumule length of the test plants. Among the test plants, Zea mays responded sensitively to eugenol.
The greater inhibitory potential of eugenol towards radicle of the test plants also confirms with earlier reports which state the stronger phytotoxicity of monoterpenes towards the roots. This could be attributed to inhibition in the cell proliferation at the root apical meristem or lowered root mitotic activity [22,24].
Moreover, in Z. mays where maximum growth retardatory effect was seen, even the internal
structure was altered when treated with eugenol. It was observed (as revealed through cross section of the roots) that in comparison to control, the diameter of cross-section of the root decreased visibly from control to 10 nl/cc. The root shape was distorted from a round in control to almost oval in treated tissues (Figure 1. a, b and c). In control (untreated root), the different cellular layers starting with the epidermis followed by cortex and vascular bundle were intact.
The vascular bundle in untreated control consisted of five metaxylem vessels in the shape of pentarch.
Plumule length
Concentration(nl/cc) O. sativa P. aureus Z. mays
Control 3.56a ± 0.37
( - )
11.14a ± 2.82 ( - )
4.91a ± 0.60 ( - )
0.5 3.17ab ± 0.10
(10.96)
10.35a ± 1.22 (7.10)
4.67a ± 0.41 (4.89)
1.0 2.99ab ± 0.05
(16.02)
9.50ab ± 0.10 (14.73)
4.42ab ± 0.20 (9.98)
2.0 3.05ab ± 0.51
(14.33)
7.10ab ± 2.40 (37.72)
3.61bc ± 0.52 (26.48)
5.0 2.53ab ± 0.39
(28.94)
3.53bc ± 0.69 (68.32)
3.06c ± 0.18 (37.68)
10.0 2.44ab ± 0.35
(31.47)
3.56bc ± 1.42 (68.05)
1.31d ± 0.20 (73.32)
20.0 2.29b ± 0.58
(35.68)
1.95c ± 0.72 (82.50)
0.71d ± 0.31 (85.54)
40.0 0.79c ± 0.62
(77.81)
1.35c ± 0.19
(87.89)
0.44d ± 0.14
Volume 03, Issue 7, [July- 2017] e-ISSN: 2394-5532, p-ISSN: 2394-823X
1(a) 2(a) 3(a)
1(b) 2(b) 3(b)
1(c) 2(c) 3(c)
Control 5.0 nl/cc 10.0 nl/c
Figure 1. T. S. of Zea mays roots treated with no oil i.e. control (1), 5.0 nl/cc (2) and 10.0 nl/cc (3) eugenol. (a) Full view, (b) Details of vascular bundle and (c) Details of epidermis and cortex.
Volume 03, Issue 7, [July- 2017] e-ISSN: 2394-5532, p-ISSN: 2394-823X
transport system of the plant. The decrease in the size and increase in the number of metaxylem vessels might be indicative of water retention by the plant to cope with eugenol caused stress. Development of aerenchyma in the roots may have the function in water aquisition since such tissues decrease the metabolic cost of growing roots because of the reduced presence of cells in respiration [25]. There are also other reports which support similar findings on anatomical changes caused as a result of exposure to essential oils [26,27,28]. It is thus clear that eugenol causes disruption in the internal tissue organization of the plants and leads to retardation in their growth and development. The study, therefore, aims at utilizing such plant essential oils/ monoterpenes for weed management as eco-friendly bioherbicides in agro-ecosystems.
IV. ACKNOWLEDGEMENT
Nitina Ahuja is thankful to UGC-BSR, New Delhi for providing financial assistance in the form of fellowship.
BIBLIOGRAPHY
[1] Harborne, J.B. 1988. Introduction to Ecological Biochemistry. 3rd Edition. Academic Press, London.
[2] Langenheim, J.H. 1994. Higher plant terpenoids: a phytocentric overview of their ecological roles. Journal of Chemical Ecology, 20: 1223-1280.
[3] Burt, S. 2004. Essential oils: their antibacterial properties and potential applications in foods – a review. International Journal of Food Microbiology,94: 223-253.
[4] Muller, W.H. and Muller, C.H. 1964. Volatile growth inhibitors produced by Salvia spp. Bulletin of the Torrey Botanical Club 91: 327-330.
[5] Singh, H.P., Batish, D.R., Kaur, S., Ramezani H., Kohli R.K. 2002a. Comparative phytotoxicity of four monoterpenes against Cassia occidentalis. Annals of Applied Biology, 141: 111 – 116.
[6] Batish, D.R., Lavanya, K., Singh, H.P. and Kohli, R.K. 2004. Phytotoxicity of lemon-scented eucalypt oil and its potential use as a bioherbicide. Crop Protection,23: 1209-1214.
[7] Kordali, S., Cakir, A. and Sutay, S. 2007. Inhibitory effects of monoterpenes on seed germination and seedling Growth. Verlag der Zeitschrift fur Naturforschung,62c: 207-214.
[8] Rolli, E., Marieschi, M., Maiettic, S., Sacchettic, G. and Bruni, R. 2014. Comparative phytotoxicity of 25 essential oils on pre- and post-emergence development of Solanum lycopersicum L.: A multivariate approach. Industrial Crops and Products,60:280-290.
[9] Synowiec, A., Kalemba, D., Drozdek, E. and Bocianowski, J. 2017. Phytotoxic potential of essential oils from temperate climate plants against the germination of selected weeds and crops. Journal of Pest Science,90: 407-419. [10] Tworkoski, T. 2002. Herbicide effects of essential oils. Weed Science,50: 425–431.
[11] Barney, J.N., Hay, A.G. and Weston, L.A. 2005. Isolation and characterization of allelopathic volatiles from mugwort (Artemisia vulgaris). Journal of Chemical Ecology31: 247-265.
[12] Azizi, M., Alimoradee, L. and Rashedmohassel, M.H. 2006. Allelopathic effects of Buniumpersicum and Cuminum
cyminum essential oils on seed germination of some weed species. Iranian Journal of Medicinal and Aromatic Plants22: 198-208.
[13] Setia, N., Batish, D.R., Singh, H.P. and Kohli, R.K. 2007. Phytotoxicity of volatile oil from Eucalyptus citriodora against some weedy species. Journal of Environmental Biology28: 63-66.
[14] Azirak, S. and Karaman, S. 2008. Allelopathic effect of some essential oils and components on germination of weed species. ActaAgriculturae Scandinavica58: 88-92.
[15] Hamdi A., Maiouli, K., Heyden, Y.V., Flamini, G. and Marzouk, Z. 2017. Phytotoxic activities of essential oils and hydrosols of Halophyllum tuberculatum. Industrial Crops and Products,97: 440-447.
[16] Bainard, L.D., M.B. Isman, and M.K. Upadhyaya. 2006. Phytotoxicity of clove oil and its primary constituent eugenol and the role of leaf epicuticular wax in the susceptibility to these essential oils. Weed Science,54: 833–837. [17] Boyd, N.S. and E.B. Brennan. 2006. Burning nettle, common purslane, and rye response to a clove oil herbicide.
Weed Technology, 20: 646–650.
[18] Meyer, S.L.F., Lakshman, D.K., Zasada, I.A. Vinyard, B.T., Chitwood D.J. 2008. Dose-response effects of clove oil
from Syzygium aromaticum on the rootknot nematode Meloidogyne incognita. Pest Management Science,64: 223–
229.
[19] Stokłosa, A., Matraszek, R., Isman, M.B. and Upadhyaya, M.K. 2012. Phytotoxic Activity of Clove Oil, Its Constituents, and Its Modification by Light Intensity in Broccoli and Common Lambsquarters (Chenopodium album). Weed Science,60:607-611.
Volume 03, Issue 7, [July- 2017] e-ISSN: 2394-5532, p-ISSN: 2394-823X [21] Singh, H.P., Kaur, S, Mittal S., Batish, D.R., Kohli, R.K. 2009. Essential Oil of Artemisia scoparia inhibits plant growth by generating Reactive Oxygen Species and causing oxidative damage. Journal of Chemical Ecology,35:
154–162.
[22] Nishida, N., Tamotsu, S., Nagata, N., Saito, C. and Sakai, A. 2005. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. Journal of Chemical Ecology,31: 1187-1203.
[23] De Martino, L., Mancini, E., De Almeda, L.F.R., De Feo, V. 2010. The antigerminative activity of twenty seven monoterpenes. Molecules,15: 6630-6637.
[24] Hsiung, Y.C., Chen, Y.A., Chen, S.Y., Chi, W.C., Lee, R.H., Chiang, T.Y. and Huang, H.J. 2013. Volatile myrcene
inhibits growth and activates defense responses in rice roots. Acta Physiologiae Plantarum35:2475-2482.
[25] Souza, T.C., Castro, E.M., Magalhaes, P.C., Lino, L.O., Alves, E.T. and Albuquerque, P.E.P. 2013.
Morphophysiology, morphoanatomy, and grain yield under field conditions for two maize hybrids with contrasting response to drought stress. Acta Physiologiae Plantarum,35: 3201-3211.
[26] Muller, W.H. and Hauge, R. 1967. Volatile growth inhibitors produced by Salvia leucophylla: effect on seedling anatomy. Bulletin of the Torrey Botanical Club,94: 182-191.
[27] Lorber, P. and Muller, W.H. 1976. Volatile growth inhibitors produced by Salvia leucophylla: effects on seedling root tip structure. American Journal of Botany,63: 196-200.