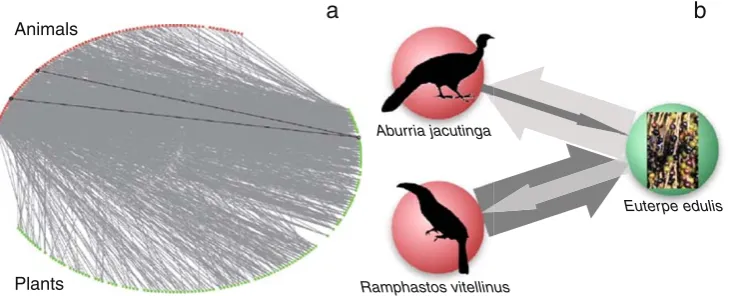
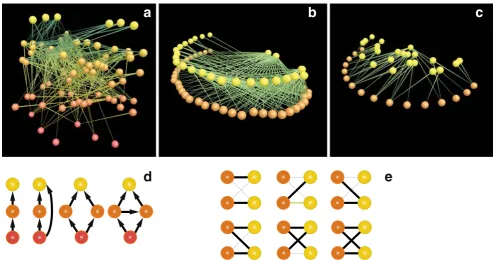
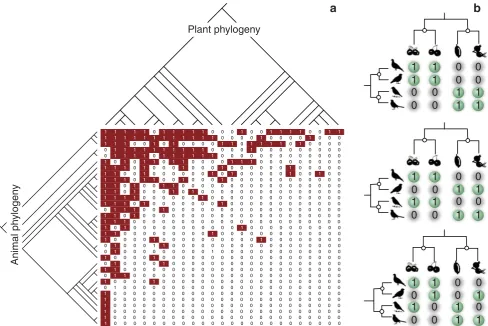
Coevolution in Multispecific Interactions among Free-Living Species
7
0
0
Full text
Figure
Related documents
