_____________________________________________________________________________________________________
*Corresponding author: E-mail: [email protected];
ISSN:2457-0125
Dietary Inclusion of Fermented
Parkia
in Feeds as
an Organic Strategy to Improve Feed Quality
and Antioxidant Parameters of African Catfish
(
Clarias gariepinus
) Fingerlings
J. Abafi
1,2*, M. Aliyu-Paiko
2, A. Aliyu-A
2, K. M. Adamu
3and M. A. King
11Department of Chemical Sciences, Biochemistry Unit, Federal Polytechnic, P.M.B. 55, Bida,
Niger State, Nigeria.
2
Department of Biochemistry, Ibrahim Badamasi Babangida University, P.M.B. 11, Lapai, Niger State, Nigeria.
3Department of Biological Sciences, Ibrahim Badamasi Babangida University, P.M.B. 11, Lapai,
Niger State, Nigeria.
Authors’ contributions
This work was carried out in collaboration of all authors. Author JA designed the study and performed the laboratory analysis. Author MAP wrote the protocol and the first draft of the manuscript. Authors AAA and KMA managed the literature searches. Author MAK performed the statistical analysis. All authors read and approved the final manuscript.
Article Information
DOI: 10.9734/AJB2T/2019/v5i330059 Editor(s): (1) Dr. Md. Shaheed Reza, Department of Fisheries Technology, Bangladesh Agricultural University, Bangladesh. Reviewers: (1) Rômulo Batista Rodrigues, Federal University of Rio Grande do Sul, Brazil. (2)Rakpong Petkam, Khon Kaen University, Thailand. (3)Vera Lúcia Viana do Nascimento, Federal Institute of Piauí- Campus Teresina Central, Brazil. Complete Peer review History:http://www.sdiarticle4.com/review-history/50430
Received 18 June 2019 Accepted 20 August 2019 Published 26 September 2019
ABSTRACT
Aim: The aim of this study is to assess fermented parkia infusion in feed as organic strategy to improve feed quality and immune system of African catfish (Clarias gariepinus) fingerlings.
Place and Duration of Study: Department of Biochemistry, Ibrahim Badamasi Babangida University, Lapai, Niger State, Nigeria, between 1st January 2017 and 25th March 2017.
Methodology: In the present study, feed ingredients were fermented for 72 hrs with probiotic starter culture from Parkia biglobosa (TF diet) in comparison to a control diet with similar
ingredients to the experimental diets but totally unfermented (CF diet) and used to make feed pellets. The experimental feeds were fed at 5% of body weight of each dietary treatment to triplicate groups of 20 fish per tank (100 Liter capacity), two times daily for 10 weeks. Proximate composition and mineral content of diets and whole fish, Feed efficiency, growth performance and Biochemical parameters of the fish were evaluated.
Results: The result of proximate compositions of experimental diets revealed a significantly higher protein and ash contents in TF compared to the CF. Carbohydrate content was significantly lower in TF diet while crude fibre, moisture and fat contents were not significantly higher in CF compared to TF. The highest values of some minerals such as sodium (Na), potassium (K), phosphorus (P) and magnesium (Mg) were also recorded in the TF while the least was recorded in CF. The result of feed efficiency and growth performance indicated similar feed acceptance and intake between fish fed the fermented diet and the control. The activities of serum antioxidant enzymes activities of superoxide dismutase and catalase were significantly (P < 0.05) higher in serum of fish fed TF compared to that in fish fed CF diet.
Conclusion: It can be deduced from this study that the fermented diet improved the immunity of fish and does not adversely affect the growth performance.
Keywords: Functional feeds; probiotics; Clarias gariepinus.
1. INTRODUCTION
Aquaculture according to Ayinla [1] is the fastest growing food producing industry in the world providing sustainable livelihood opportunities and food security for the ever-increasing world population [2]. With the global increase demand for fish over the past two decades, aquaculture production is likely to double in the next fifteen years, as a result of wild fisheries approaching biological limits [3]. Therefore, there is an immense need to intensify fish farming, especially in the developing nations like Nigeria [4]. Aquaculture has greater potential to meet the increasing demand for fish; however, sustainable supply and cost of feed ingredients for aquaculture pose a major challenge. Diets with proper nutrient balance (i.e. functional feed) are important in enhancing fish health and production. Furthermore, the intensification of aquaculture practices applied to maintain high levels of production could result in adverse environmental situations that may acutely stress the fish,
altering their biochemical functions and
suppressing their immune responses. As a result, diseases, most especially bacterial infections tend to thwart fish production thereby threatening the sustainability of aquaculture [5]. Thus,
substantial reliance on vaccines and antibiotics to combat fish diseases is inevitable. Sequential use of antibiotics to control diseases creates adverse effect that result in accumulation of antibiotics residues in the tissue, which consequently leads to immune suppression among others [6]. Suitably, the development of natural growth promoters has become important
and urgent alternative for healthier fish
production. It is suggested that the growth performance of cultured species could be enhanced through addition of probiotic to diet that will improve functional characteristics of feed and consequently increase feed efficiency [7].
In the era of diminishing feed quality, fermentation of feed ingredients through probiotic could play a role in complementing feed fortification. In addition, the ingestion of the
fermented feed along with the live
microorganisms could further maintain health-balance of fish by stimulating the immune system, aiding in nutrient digestion and increasing the
resistance against enteric opportunistic
pathogens [8]. The functionality of enteric microbiota depends on the ability of beneficial microorganisms to interact with the digestive tract, which benefits the host by the influence on
the biological functions [9]. Beneficial
microorganisms or probiotic activate the
digestion process by providing digestive
enzymes and vitamins as well as the ability of nutrients to compete the adhesion sites to enhance the feed utilization [10]. Furthermore, the antimicrobial substances produced by intestinal microorganisms enhance the immune
response and decrease the pathogenic
bacteria [11]. This present study was therefore designed to investigate the effects of dietary inclusion of fermented iru infusion as an organic strategy to improve feed quality/utilization,
growth performance and biochemical
2. MATERIALS AND METHODS
2.1 Experimental Fish and Husbandry Management
The African Catfish fingerlings were purchased from a commercial hatchery in Bida local government and transported to the fish research laboratory of the department of Biochemistry, IBB University, Lapai, Nigeria. The fish were acclimated to the experimental conditions and fed control diet for two weeks. At the end of the adaptation period, a total of 120 fish (with mean initial weight of 7.4 g) were randomly divided into 2 groups of 3 replicates each. Each replicate contained 20 fish. All cultured tanks contained bore-hole water from the University water supply. Water temperature, pH and dissolved oxygen (DO) were maintained within optimum range for the culture of African catfish. Fish were reared
under natural photoperiod and hand-fed
assigned experimental diets at 5% body weight twice daily at 9:00 and 16:00 hours for 10 weeks. Approximately half of the water in the system was changed four (4) times a week to reduce the regular accumulation of nitrogenous residue from feed.
Fish were weighed individually at the beginning and end of the experiment, whereas batch weighing per tank was carried out every week to monitor growth performance alongside adjusting feeding rate. At the end of the experiment after 10 weeks, surviving fish were pooled together and randomly grouped into Four (4) per treatment and used to determine serum biochemical parameters and carcass proximate compositions.
2.2 Production of Starter Cultures from Fermentation of Feed
To obtain “Parkia” water, Parkia biglobosa
(African locust beans)was prepared according to
the modified method of Odunfa and Oyewole [12], shown in the flow chart in Fig. 1.
Production of Starter culture involved soaking of
dadawa in approximately 3 litres of distilled water in a plastic container, which was covered for 72 hours. Thereafter, five (5) litres of distilled water was used to wash the slimily dadawa. The solid
dadawa debris was sieved out while the surface
water called dadawa or parkia water was
collected.
African locust beans seed ↓
Sorted, cleaned and washed ↓
Cooked for 14 hours ↓
Washed, sieved and parboiled for 1 hour ↓
Sieved cotyledon ↓
Allowed cool and dehulled ↓
Boiled for another 9 hours ↓
Poured into plastic basket lined with jute sack (covered tightly to prevent heat escape)
↓
Packed in a clean plastic container, Ferment while still hot in a dark, warm place for 72 hours.
↓
Dadawa
Fig. 1. Production of Dadawa from African locust beans, Parkia biglobosa Table 1. Experimental diets and feed
formulation Ingredients Control feed
(%)
Test feed (%)
Fish meal 24 24
Soybean meal 27 27
Corn starch 34 34
Vegetable oil 8 8
Vitamin premix 2 2
Mineral premix 2 2
Tapioca 3 3
Parkiawater - +
Briefly after 72 hours, the surface water (called
2.3 Proximate Analysis
Experimental diets and final fish carcass were
analyzed for moisture content, proximate
composition of crude protein, crude lipid, crude fiber, ash and carbohydrate (Nitrogen free extract; NFE) contents following standard AOAC methods [13].
2.4 Calculations of Feed Efficiency and Growth Parameters
Feed efficiency and growth parameters were calculated using the following:
Feed intake (FI) = Total feed intake (g)/number of live fish
Protein intake (PI) = Feed intake (g)
percent protein in the diet
Feed conversion ratio (FCR) = Feed intake (g) /wet weight gain (g)
Protein efficiency ratio (PER) = wet weight gain (g)/total protein intake
2.5 Growth Performance Parameters
Fish survival (%) = final number of surviving fish/initial number of fish x 100
Weight gain (WG) =
Weight gain (WG %) = × 100
Specific growth rate (SGR %) = ×
100
Where: refers to the mean final weight, is the mean initial weight and is the feeding trial period in days.
2.6 Determination of Antioxidant Enzymes Activities
Comparative study of biochemical profile was investigated by sampling blood from fish. The
experimental fish were anesthetized and
maintained in water at 5°C. The blood was obtained from ten (10) randomly selected fish in three containers of twenty (20) fish per tank. This was done as described by Argungu et al. [14].
The blood was transferred into anticoagulant free test tube to clot for serum biochemical analyses. Serum thereof was obtained by centrifugation using a bench centrifuge for 10 min at 3,000 rpm, after which the serum was collected by the use of micropipette and transferred into anticoagulant free test-tube and stored in refrigerator for analysis.
Superoxide dismutase (SOD) and catalase (CAT) activities were determined spectro-photometrically from the serum according to the modified methods of Misra et al. [15] and Beers et al. [16], respectively.
2.7 Statistical Analysis
The Results were presented as mean ± SD of two values. Mean values for all monitored parameters were analyzed by one-way analysis of variance (ANOVA). P values <0.05 were considered significant when compared by Turkey’s test. All statistical analyses were carried out using SPSS software, version 24.0.
3. RESULTS
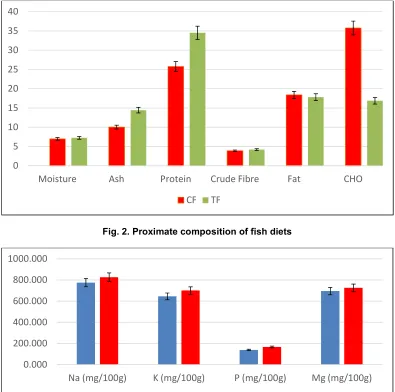
3.1 Proximate Compositions and Mineral Content of Experimental Diets
The result for proximate compositions and mineral content of experimental diets are presented in Fig. 2. The result indicated that fermented diet (TF) contained significantly higher protein and ash contents compared to the CF. Carbohydrate content was significantly lower in TF diet. Crude fibre, moisture and fat contents were not significantly higher in CF compared to TF. The highest values of analyzed minerals such as sodium (Na), potassium (K), phosphorus (P) and magnesium (Mg) were also recorded in the TF while the least was recorded in CF.
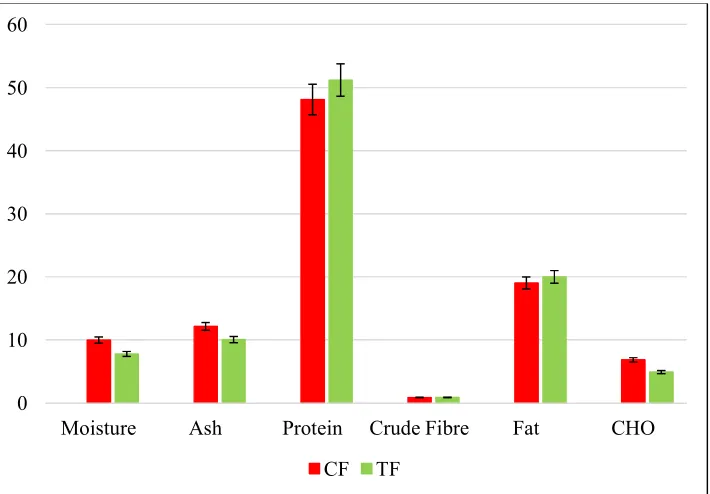
3.2 Proximate Compositions and Mineral Contents of Fish Carcass
3.3 Feed Efficiency of African Catfish Fed Fermented Diet
The result of feed efficiency of African Cat fish fed fermented diet is shown in Fig. 4. As indicated, feed acceptance and intake were similar between fish fed the fermented diet and the control.
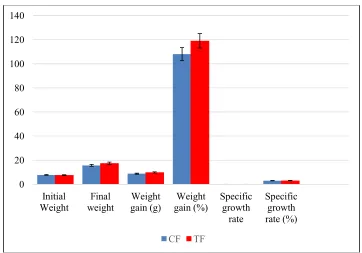
3.4 Growth Performance of African Catfish Fed Fermented and Non- Fermented Diets
The result for growth performance of African catfish fed experimental diets as shown in Fig. 5 followed a similar trend to that observed for feed
efficiency, with no significant (P > 0.05) difference between TF and CF treatment groups.
3.5 Activities of Serum Antioxidant Enzymes in African Catfish Fed Fermented and Non-Fermented Diets for 10 Weeks
The result for activities of serum antioxidant enzymes in African catfish fed fermented and non-fermented diets is presented in Fig. 6. The activities of antioxidants enzymes; superoxide dismutase and catalase were significantly (P < 0.05) higher in serum of fish fed TF compared to that in fish fed CF diet.
Fig. 2. Proximate composition of fish diets
Fig. 3. Mineral contents of TF and CF diets
0 5 10 15 20 25 30 35 40
Moisture Ash Protein Crude Fibre Fat CHO
CF TF
0.000 200.000 400.000 600.000 800.000 1000.000
Na (mg/100g) K (mg/100g) P (mg/100g) Mg (mg/100g)
Fig. 4. Proximate composition of fish carcass
Fig. 5. Proximate composition and mineral contents of fish carcass
4. DISCUSSION
Functional feeds represent one of the most intensively explored and widely promoted areas in aquaculture industry [17]. According to
Aworh et al. [18] the use of probiotics for fermentation serves as a useful strategy in complementing feed fortification. This strategy promotes efficient absorption of nutrients due to the ability of probiotics (such as Bacillus species) 0
10 20 30 40 50 60
Moisture Ash Protein Crude Fibre Fat CHO
CF TF
0.000 100.000 200.000 300.000 400.000 500.000 600.000
Na (mg/100g) K (mg/100g) P (mg/100g) Mg (mg/100g)
to produce digestive enzymes (amylases, proteases, and lipases, etc.) and provide simpler nutrients (vitamins, fatty acids and amino acids), which could enhance the digestive
process and feed utilization. This will result in improved general health condition and
growth performance of aquatic animals
[19].
Fig. 6. Nutrient utilization of African Catfish fed fermented and non-fermented diets for 10 weeks feeding trial
Fig. 7. Growth parameters of African catfish, Clarias gariepinus fed experimental diets for a period of 10 weeks feeding trial
0 20 40 60 80 100 120
Feed Intake (g)
Feed Conversion
Ratio
Protein Intake (g)
Protein Efficiency
Ratio
Fish Survival (%)
CF
TF
0 20 40 60 80 100 120 140
Initial Weight
Final weight
Weight gain (g)
Weight gain (%)
Specific growth
rate
Specific growth rate (%)
Fig. 8. Activities of serum antioxidant enzymes of African catfish fed fermented and non-fermented diets for 10 weeks
Proximate composition analysis is an important criterion in determining the nutritional quality of feed in animal husbandry [20]. In the present study, there was significant (P< 0.05) increase in protein and ash contents of the formulated diet after fermentation. The increase in crude protein content could be attributed to the ability of the suspected microbes (such as Bacillus species) present in TF diet to carry out different proteolytic enzyme activities, thereby producing free amino acids during fermentation [21].
Protein obtained from the diet provides proper ratio of amino acid necessary for growth performance, which is utilized to synthesize new proteins or maintain existing proteins in tissues while excess protein, is converted to energy [22]. Proteins and their encoding genes in feeds also regulate cellular transport of micro-minerals in fish [23].
The protein content of whole fish in terms of nutritional value depends on the physiological utilization of dietary amino acids after digestion, absorption, and a minimal rate of oxidation [24]. The notable numerical increase in protein content of whole fish in this study indicates
adequate utilization and significant impact on nutrient storage of the fermented diet. The increased crude protein content observed in whole fish is in line with the findings of Degani [25], who reported that proteins are the major macromolecules in fish tissue and could make up to 65-75% of the total organic matter on a dry matter basis. Abdel-Tawwab et al. [26] added that changes in protein contents in the fish body could be attributed to changes in deposition rate in muscle and can consequently have effect on growth performance.
The observed decrease in carbohydrate content of fermented (TF) diet may be due to the ability of the microorganisms to utilize carbohydrate as their major energy source from organic matter during microbial protein synthesis [27]. Jonathan et al. [21], Boateng et al. [28] individually reported decrease in carbohydrate content during fermentation as a result of increased production of several digestive enzymes such as amylase, glucosidase and galactanase by fermentative microbes. This observation is also similar to that reported by Nitschke and Pastore [29], Oladele and Oshodi [30], who stated that Bacillus sp produced several enzymes during 0.04
2.6
0.06
3.85
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5
Superoxide dismutase (U/L) Catalase (U/L)
fermentation which are capable of degrading carbohydrates into simple sugars that are used by the microorganism as energy source for metabolic activities.
Carbohydrate is one of the dietary ingredients that profoundly affect feed utilization. Feed Increase in carbohydrate level above 10% dietary dry matter can result in reduced feed intake and utilization in aquatic animals [31]. The carbohydrate decrease observed in the present study is similar with the findings of Hemre et al. [31] and Erfanullah, [32] who reported improved feed utilization and protein retention in rainbow trout and Atlantic salmon when fed with diets containing low levels of carbohydrate content. Consequently, the decrease in carbohydrate level as a result of fermentation is appropriate for the cultured fish.
Furthermore, the level of carbohydrate content observed in the present study is appropriate for the cultured fish as a preferred oxidative substrate for nervous tissue and blood cell. Consequently, the carbohydrate present in the diet can lower gluconeogenic activity, thus diverting amino acids away from oxidative pathways essential for growth promotion.
The result of proximate composition of TF diet
showed decrease in lipid content after
fermentation compared to CF diet. This could be as a result of microorganisms producing lipases, which converts the polymeric forms of the lipids to soluble monomers [33]. Furthermore, the loss in lipid content of TF could be attributed to it utilization as modifiers during the fermentation process. This is in line with the argument of Vakili et al. [34] who revealed that lipids (or fatty acids) serve as useful fermentation modifiers.
Different flavors and aroma were observed during the fermentation of diet. This may be as a result of metabolism of lipid components and formation of other microbial metabolites. Modupe et al. [35] reported that the degradation of fat and other available compounds produce aroma during fermentation process. This could also be the cause for the depletion of lipids after the fermentation of feed.
Lipids (fats) are high-energy nutrients that can be utilized to spare protein in aquaculture feeds. Lipids make up about 7-15 percent of fish diets, which provide twice the energy density of proteins and carbohydrates in fish [36]. Other important functions of lipids include supplying
fatty acids, improvement in absorption of fat-soluble vitamins, precursors of hormones and prostaglandins and as building blocks of cellular and membrane structures in fish [37]. Dietary lipids play an important role in the production and regulation of active modulators of bone metabolism (such as eicosanoids, cytokines, leucotriens, lipoxygenase). Dietary lipid levels are known to affect bone formation and reabsorption in fish.
The Ash content gives an idea of the inorganic content of the sample from where the mineral content could be obtained [38]. The analysis of mineral compositions of TF diet indicated significantly (P< 0.05) higher contents of sodium, potassium, phosphorus and magnesium than in the control diet. The result obtained were higher than that reported for Berhi cultivar by Kadam et al. [39], who stated that this increase may be due to contribution by fermentation micro organisms in the breakdown of the organic components of the samples during the period of fermentation [40].
The increase in mineral content may also be as a result of the fermentative microorganisms, which provide optimum pH condition for enzymatic degradation of phytates that form complexes with essential minerals [41]. Niba et al. [42] reported an improved bioavailability of minerals in liquid fermented pig feed. Similar trend was also observed on the effect of fermentation on the nutriet status of locust bean where an increase of 30% in ash content was recorded after fermentation [43]. Furthermore, Adams [44] reported an increase in the concentrations minerals after fermentation of feed ingredients.
Fish require the same minerals as terrestrial animals because of their involvement in skeletal formation, as cofactors of many metallo-enzymes, for osmoregulation and other metabolic functions [45].
At the end of the experimental feeding trial,
improved nutrient utilization and growth
utilization when turbot, catla carp and hybrid striped bass feeds were supplemented with bacteria Vibrio proteolyticus [48]. Adamu et al. [49] also reported that, African catfish fed fluted pumpkin leaf (FPL) and parboiled fermented African locust beans (PFALB) had better growth performance and nutrient utilization.
Probiotic inclusion in diet enhances growth performance and nutrient utilization in fish due to the modulatory effect on feeding, metabolism and feed digestibility [7]. The increased feed utilization and growth observed in the whole fish was similar to that of Ai et al. [50], Lamari et al. [51] who reported that Bacillus and Lactobacillus genera seem to be the most related with nutrient utilization and growth improvement, which may have influenced appetite, conversion ratio or reducing myostatin transcription [52], a protein responsible for modifying muscle growth and development in fish [50].
The relatively higher growth performance and nutrient utilization of fish fed TF diet may further be attributed to fermentation process as it has been proven to enhance the quality and palatability of fish feeds [53]. On the other hand, the relatively faster growth rate potential of those fed the fermented feed reveals that the fish accepted and consumed the fermented feed pellets better. Furthermore, due to increased acceptance and consumption of the fermented feed pellets, nutrients such as protein and lipid deposition in the muscle could significantly increase fish growth [26].
The increased growth may also be due to the ability of the suspected microbes in TF diet to adhere to the gastrointestinal tract of the catfish hence producing wide range of relevant digestive enzymes (amylase, lipase and protease) which hydrolyze the indigestible components thus synthesizing biotin and vitamin B12, which
enhances feed utilization and digestibility [53].
Similarly, the relatively higher protein efficiency ratio (PER) observed in fish fed TF diet showed improved protein utilization in African catfish. This contributed to optimization of protein used for fish growth. The improvements in the biological value of the fermented diets have shown its efficiency in converting protein to flesh in fish [54].
Oxidative stress refers to the cellular condition that indicates a physiological imbalance between the levels of antioxidants and that for oxidants
(i.e. free radicals or reactive species), such that the imbalance favours oxidants. Thus, under normal physiological conditions, aquatic animal cells maintain a balance between generation and removal of reactive oxygen species [55].
The antioxidant enzymes, superoxide dismutase
(SOD), catalase (CAT) and glutathione
peroxidase (GPx) are considered the first line of antioxidant defense mechanism against oxidative stress. Superoxide Dismutase is considered the first enzyme responsible for scavenging of free radical and thus protecting cells from been damaged [56]. In addition, SOD converts superoxide anion (O-2) to hydrogen peroxide (H
2O2) while CAT reduces H2O2 to water [57].
Activities of serum antioxidant enzymes revealed that the fish fed fermented (TF) diet suggested increased level of plasma antioxidant enzymes of SOD and CAT compared to the control fish. Similar results were observed when fish fed probiotics (Bacillus coagulans B16 and R.
palustris) elicited increased SOD and CAT
activities in tilapia [58]. Salinas et al. [59] reported that respiratory burst activity of teleost fish (Sparus aurata L.) increased in vitro by the
addition of heat-inactivated Lactobacillus
delbrueckii sp.
The significant increase in the antioxidant enzymes activities could be attributed to the
microbial production of proteins through
proteolytic enzyme activities in the fermented diet, which could serve as precursor in the production of the organic enzymes.
A properly balanced gut microbiota has the ability to positively affect the integrity of the intestinal barrier against colonization by pathogens [60], through its protective and metabolic function and can stimulate the immune system in an anti-inflammatory manner [61].
Furthermore, this elevation in the activities of the antioxidant enzymes is an indication of enhanced scavenging of free radicals in the blood of the TF fed fish group. Therefore, the interplay between free radicals, antioxidants, and diseases in cells, tissues and organisms in the experimental fish is important in maintaining good health [62].
5. CONCLUSION
Fermented diet may be successfully used as an alternative to conventional fish feed for African
antioxidant system as revealed by the results findings.
6. RECOMMENDATIONS
i. Further research should be carried out to ascertain the appropriate amount of microbial cells to be employed in feed fermentation for aquaculture.
ii. The duration of feed fermentation should be prolonged to obtain higher feed microbial pre-digestion for enhanced feed utilization.
COMPETING INTERESTS
Authors have declared that no competing interests exist.
REFERENCES
1. Ayinla OA. Aquaculture development and
appropriate enterprise combination in the BRACED States. In the High Level Meeting of Experts and the Meeting of
BRACED States Commissioners for
Agriculture. Songhai Farms, Port -Harcourt. 2012;2:1-41.
2. Wang Y, Xu N, Xi A, Ahmed Z, Zhang B, Bai X. Effects of Lactobacillus plantarum
MA2 isolated from Tibet kefir on lipid metabolism and intestinal microflora of srats fed on high-cholesterol diet. Appl Microbiol Biotechnol. 2009;84:341–347. 3. Fazio F, Marafioti S, Torre A, Sanfilippo M,
Panzera M, Faggio C. Haematological and serum protein profiles of Mugil cephalus:
effect of two different habitat.
Ichthyological Research. 2013;60:36–42.
4. EDR. Analysis of fish feed market in Delta
State. Economic Development Research No 7. PIND Foundation. 2014;63.
5. Abd El-Rhman AMA, Khattab YA, Shalaby
AM. Micrococcus luteus and
Pseudomonas species as probiotics for
promoting the growth performance and health of Nile tilapia, Oreochromis niloticus. Fish Shell Fish Immuol. 2009;27:175-180.
6. Nayak SK, Swain P, Mukherjee SC. Effect
of dietary supplementation of probiotic and vitamin C on the immune response of Indian major carp, Labeo rohita (Ham.). Fish Shellfish Immunol. 2007;23:892-896.
7. Pereira LF, Gonçalves AF, Domingues
MD, Peixoto S, Braga K, Sansuwan G, et al. Growth performance, antioxidant and
innate immune responses in European seabass fed probiotic supplemented diet at three rearing temperatures. Frontiers in Marine Science Journal; 2004.
8. Llewellyn MS, Boutin S, Hoseinifar SH,
Derome N. Teleost microbiomes: The state of the art in their characterization,
manipulation and importance in
aquaculture and fisheries. Front. Microbiol. 2014;5:207.
DOI: 10.3389/fmicb.2014.00207
9. Ramírez C, Romero J. Fine flounder
(Paralichthys adspersus) microbiome
showed important differences between wild and reared specimens. Front. Microbiol. 2017;8:271.
10. Hovda MB, Lunestad BT, Fontanillas R,
Jan Thomas Rosnes JT. Molecular
characterisation of the intestinal microbiota of farmed Atlantic salmon (Salmo salar L.). Aquaculture. 2007;272:581-588.
11. Ringø E, Sperstad S, Myklebust R, Refstie
S, Krogdahl A. Characterisation of the microbiota associated with intestine of Atlantic cod (Gadus morhua L.): The effect of fish meal, standard soybean meal and a bioprocessed soybean meal. Aquaculture. 2006;261:829-841.
12. Odunfa SA, Oyewole OB. Identification of
Bacillus species from Iru, a fermented African locust bean product. J. Basic Microbiol. 1986;26:101-108.
13. AOAC. Association of Official Analytical Chemists. Official method of analysis. 17th Edn, Gaithersburg, USA; 1990.
14. Argungu LA, Mohammad SNA, Abu-Naim
BB, Roushon A, Aminur-Rahman M, Arshad A, Aliyu-Paiko M. Biochemical and
haematological profile of Malaysian
snakehead, Channa striatus (Bloch).
Journal of Food, Agriculture and
Environment. 2017;13(1):12-17.
15. Misra HP, Fridovich I. The role of
superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. Journal of Biology Chemistrty. 1972;247(10):3170-5.
16. Beers RF, Sizer IW. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. Journal of Biology Chemistry. 1952;195(1):133- 40.
17. Hasler CM. Functional foods: Benefits,
concerns and challenges—a position
18. Aworh OC. The role of traditional food
processing technologies in national
development: The West African
experience. International Union of Food Science & Technology. 2008;1-18.
19. Ray RC, Sivakumar PS. Traditional and
novel fermented foods and beverages from tropical root and tuber crops. Int. J. Food Sci. Technol. 2009;44:1073–1087.
20. Qayyum MMN, Butt MS, Anjum FM,
Nawaz H. Composition analysis of some selected legumes for protein isolates recovery. J. Anim. Plant Sci. 2012;22: 1156–1162.
21. Jonathan SG, Fadahunsi IF, Garuba EO.
Fermentation studies during the production of “iru‟ from bambara nut (Voandzeia
subterranea l. Thours), an indigenous
condiment from South-Western Nigeria. EJEAFChe. 2011;10(1):1829-1836. 22. Luo J, Goetsch AL, Nsahlai IV, Sahlu T,
Ferrell CL, Owens FN, Galyean ML, Moore JE, Johnson ZB. Metabolizable protein requirements for maintenance and gain of growing goats. Small Ruminant Research. 2004b;53(3):309-326.
23. Bakke AM, Glover C, Krogdahl A. Feeding,
digestion and absorption of nutrients. In Multifunctional Gut of Fish. 2010;30:57-110.
24. Montero D, Basurco B, Nengas I, Alexis M,
Izquierdo M. CIHEAM, Zaragoza, Spain. 2005;73-90.
25. Degani G. The effect of different protein level and temperature and feed utilization, growth and body composition of Clarias gariepinus (Burchell 1822). Aquaculture.
1989;76:293-301.
26. Abdel-Tawwab M, Khattab YAE, Ahmed
MH, Shabby AME. Compensatory growth, feed utilization, whole body composition and haematological changes in starved juvenile Nile tilapia, Oreochromis niloticus
(L). Journal of Applied Aquaculture. 2006;18(3):17-36.
27. Jasim UM, Haque KZ, Jasimuddin KM,
Mehedi-Hasan KM. Dynamics of microbial protein synthesis in the rumen-a review. Annals of Veterinary and Animal Science. 2015;2(5).
28. Boateng M, BOkai OB, Frimpong YO,
Asabere CO. A comparative study of the nutritional and microbial profiles of the raw and processed seeds of the African Locust Bean (Parkia biglobosa). Livestock
Research for Rural Development.
2004;26:178.
29. Nitschke M, Pastore GM. Production and properties of a surfactant obtained from
Bacillus subtilis grown on cassava
wastewater. Bioresource Technology.
2006;97(2):336-341.
30. Oladele EP, Oshodi AA. Effect of
fermentation on some chemical and nutritive properties of Berlandier nettle spurge (Jatropha cathartica) and physic Nut (Jatropha curcas) seeds. Pakistan Journal of Nutrition. 2008;7:292-296.
31. Hemre GI, Sandnes K, Lie O, Torrissen O,
Waagbo R. Carbohydrate nutrition in Atlantic salmon, Salmo salar L.: Growth and feed utilization. Aquac. Res. 1995;26: 149–154.
32. Erfanullah JAK. Growth rate, feed
conversion, and body composition of Catla catla, Lbeo rohita, and Cirrhinus mrigala
fry fed diets of various carbohydrate-to-lipid ratios. J. World Aqua- cult. Soc. 1998;29: 84–91.
33. Nguyen TN. Total sulfur amino acid
requirement and its application to practical diets for juvenile tilapia (Oreochromis
spp.). PhD Dissertation. Auburn University, Auburn, AL. 2007;11(3):123-127.
34. Vakili AR, Khorrami B, Danesh MM,
Parand E. The effects of thyme and cinnamon essential oils on performance, rumen fermentation and blood metabolites
in Holstein calves consuming high
concentrate diet. Asian Australas. J. Anim. Sci. 2013;26(7):935-944.
35. Modupe EO, Abiodun JO, Adesola AA.
The effect of different starter cultures on the protein content in fermented African locust bean (Parkia biglobosa) seeds.
International Journal of Engineering
Research & Technology. 2016;5(4):249-253. ISSN: 2278-0181.
36. Ng WK. Palm oil in salmoind feeds. Aqua
feeds: Fermentation and beyond.
2004;1:7-9.
37. David D, Kuhn MH, Schwarz SC.
Understanding fish nutrition, feeds and feeding; 2017.
38. Sotelo R, Sofowora A, Index EEIJ.
Medicinal plant and traditional medicine in African. Spectrum Book Ltd, Nigeria. 2007;25.
40. Jokotagba OA, Onasanya SS. The impact of fermentation on the proximate and mineral composition of Phoenix dactylifera
L flour. Covenant Journal of Physical and Life Sciences. 2015;3(1):9-14.
41. Nout MJR, Ngoddy PO. Technological
aspects of preparing affordable fermented
complementary foods. Food Control.
1997;8:279-287.
42. Niba AT, Beal JD, Kudi AC, Brooks PH. Potential of bacterial fermentation as a biosafe method of improving feeds for pigs and poultry. African Journal of Biotechnology. 2009;8(9):1758-1767. 43. Eka OU. Effect of fermentation on the
nutrient status locust beans. Food Chem. 1980;5:305-308.
44. Adams MR. Topical aspects of fermented foods. Trends in Food Science and Technology. 1990;8:140-144.
45. Jiang J, Lu S, Zhang H, Liu G, Lin K, Huang W, Luo R, et al. Dietary intake of human essential elements from a total diet study in Shenzhen, Guangdong Province, China. J Food Compos Anal. 2015;39:1-7.
46. Lara-Flores M, Olvera-Novoa MA,
Guzman-Mendez BE, Lopez-Madrid W. Use of the bacteria Streptococcus faecium
and Lactobacillus acidophilus, and the
yeast Saccharomyces cerevisiae as
growth promoters in Nile tilapia
(Oreochromis niloticus). Aquaculture.
2003;216:193-201.
47. De Schrijver R, Ollevier F. Protein diges on in juvenile turbot (Scophthalmus maximus) and effects of dietary administration of
Vibrio proteoly cus. Aquaculture. 2000;186: 107-116.
48. Li P, Galtin DM III. Evaluation of the prebiotic GroBiotic-A and brewers yeast as dietary supplements for sub-adult hybrid
striped bass (Morone chrysops x M.
saxatilis) to Streptococcus iniae infection. Aquaculture. 2005;231:445-456.
49. Adamu KM, Ikomi RB, Paiko-Aliyu M. The use of growth and biochemical indices to evaluate the interchangeability of some plant crude proteins in fishmeal in the diet of hybrid catfish. Development Journal of
Science and Technology Research.
2015;4(2):13-23.
50. Ai Q, Xu H, Mai K, Xu W, Wang J, Zhang
W. Effects of dietary supplementation of
Bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker,
Larimichthys crocea. Aquaculture.
2011;317:155-161.
51. Lamari F, Castex M, Larcher T, Ledevin M,
Mazurais D, Bakhrouf A, Gatesoupe FJ. Comparison of the effects of the dietary addition of two lactic acid bacteria on the development and conformation of sea bass larvae, Dicentrarchus labrax, and the
influence on associated microbiota.
Aquaculture. 2013;376–379.
52. Zlatica K, Jolana K. Fermentation of
cereals for specific purpose. Journal of Food and Nutrition Research. 2007;46(2): 51-57.
53. Hassaan MS, Wafa MA, Soltan MA, Goda
AS, Mogheth NMA. Effect of dietary
organic salts on growth, nutrient
digestibility, mineral absorption and some
biochemical indices of Nile Tilapia;
Oreochromis niloticus L. fingerlings. World Applied Sciences Journal. 2014;29:47- 55.
54. Ringø E, Gatesoupe FJ. Lactic acid
bacteria in fish: A review. Aquaculture. 198;160:177-203.
55. Zhang D, Pridgeon JW, Klesius PH.
Vaccination of channel catfish with
extracellular products of Aeromonas
hydrophila provides protection against
infection by the pathogen. Fish Shellfish Immunology. 2013;2:1-15.
56. Ighodaro OM, Akinloye OA. First line
defence antiodixants-superoxide
dismutase (SOD), catalase (CAT) and
glutathione perodixase (GPX): Their
fundamental role in the entire antioxidant defence grid. Alexandria Journal of Medicine.
Available:https://doi.org/10.1016/j.ajme.20 17.09.001
57. Timbrel J. Principles of biochemical
toxicology. 3rd Edition. Taylor and Francis. 2000;394.
58. Misra CK, Das BK, Mukherjee SC, Pattnaik
P. Effect of multiple injections of ß -glucan on non-specific immune response and
disease resistance in Labeo rohita
fingerlings. Fish Shellfish Immunol.
2006;20:305-319.
59. Salinas I, Diaz-Rosales P, Cuesta A. Effect of heat-inactivated fish and nonfish derived
probiotics on the innate immune
parameters of a teleost fish (Sparus
auratus L.). Veterinary Immunology and
Immunopathology. 2006;111:279–286.
60. Jandhyala SM, Talukdar R, Subramanyam
Role of the normal gut microbiota. World J.
Gastroenterol.2015;21(29):8836-8847.
61. Wu HJ, Wu E. The role of gut microbiota in
immune homeostasis and autoimmunity. Gut Microbes Journals. 2012;3(1):4-14.
62. Rahman MA. Biochemical composition of
some small indigenous fresh water fishes from the river Mouri, Khulna, Bangladesh. Pakistan Journal of Biological Sciences. 2007;10(9):1559-1561.
_________________________________________________________________________________
© 2019 Abafi et al.; This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Peer-review history: