Yeast RSC Function Is Required for Organization of the Cellular Cytoskeleton
via an Alternative
PKC1
Pathway
Bob Chai,
1Jing-mei Hsu,
1Jian Du
1,2and Brehon C. Laurent
3Department of Microbiology and Immunology and Morse Institute for Molecular Genetics, State University of New York, Brooklyn, New York 11203
Manuscript received January 2, 2002 Accepted for publication March 18, 2002
ABSTRACT
RSC is a 15-protein ATP-dependent chromatin-remodeling complex related to Snf-Swi, the prototypical ATP-dependent nucleosome remodeler in budding yeast. Despite insight into the mechanism by which purified RSC remodels nucleosomes, little is known about the chromosomal targets or cellular pathways in which RSC acts. To better understand the cellular function of RSC, a screen was undertaken for gene dosage suppressors of sth1-3ts, a temperature-sensitive mutation inSTH1, which encodes the essential ATPase subunit. Slg1p and Mid2p, two type I transmembrane stress sensors of cell wall integrity that function upstream of protein kinase C (Pkc1p), were identified as multicopy suppressors of sth1-3ts cells. Although the sth1-3ts mutant exhibits defects characteristic of PKC1 pathway mutants (caffeine and staurosporine sensitivities and an osmoremedial phenotype), only upstream components and not downstream effectors of thePKC1-MAP kinase pathway can suppress defects conferred bysth1-3ts, suggesting that RSC functions in an alternativePKC1-dependent pathway. Moreover,sth1-3tscells display defects in actin cytoskeletal rearrangements and are hypersensitive to the microtubule depolymerizing drug, TBZ; both of these defects can be corrected by the high-copy suppressors. Together, these data reveal an important functional connection between the RSC remodeler andPKC1-dependent signaling in regulating the cellular architecture.
C
HROMOSOMES are the substrates for many essen- dependent chromatin-remodeling complex, the SWI/ SNF subfamily also includes the yeast RSC (for remodels tial nuclear processes including DNA replication,repair, recombination, and transcription. These events the structure of chromatin), Drosophila brahma, and human BRG-1 and hbrm complexes (Coˆ te´et al.1994; must be coordinated with the dynamic changes in
chro-mosome structure and cell morphology that occur Kwonet al. 1994; Cairns et al. 1996; Papoulas et al. 1998). RSC is an abundant 1–2 MD complex composed throughout the cell cycle. Although chromatin
inter-feres with several of these DNA-dependent cellular func- of 15 proteins (Cairnset al.1996). Like all ATP-depen-dent remodelers, RSC contains a Snf2p/Swi2p-related tions, chromatin-remodeling complexes are able to alter
accessibility to DNA. Two general classes of remodeling ATPase subunit, Sth1p. However, despite its similarity to the yeast Snf-Swi complex, RSC is required for cell enzymes have been identified. The first includes
ATP-dependent chromatin remodelers, which use the energy viability. The G2/M arrest conferred by four tempera-ture-sensitive (ts)rscmutations suggests a requirement released from ATP hydrolysis to disrupt histone-DNA
interactions (for reviews seeKingstonandNarlikar for RSC function in cell cycle progression (Tsuchiya et al.1992;Caoet al.1997;Duet al.1998;Angus-Hill 1999;Vignaliet al.2000). The second includes enzymes
that covalently modify histones by adding ubiquitin, ace- et al. 2001). RSC function has also been implicated in transcription (Caoet al.1997;Cairnset al.1998, 1999; tyl, methyl, or phosphate groups (for reviews seeStrahl
andAllis2000;Berger2001;BergerandFelsenfeld Moreira and Holmberg 1999; Yukawa et al. 1999; Angus-Hillet al.2001). Interestingly, RSC shares func-2001; Jenuwein and Allis 2001; Roth et al. 2001).
These two types of enzyme function coordinately to tional and structural similarities with human SWI/ SNF-B (Nieet al.2000;Vignali et al.2000).
regulate transcription at some promoters (seeFryand
Peterson2001). Cellular signaling plays an important role in relaying extracellular information to intracellular pathways that In addition to yeast Snf-Swi, the prototypical
ATP-impact on nuclear function. Protein kinase C (PKC)/ ras and Ca2⫹-mediated membrane signaling has been
1These authors contributed equally to this work. shown to regulate hSWI/SNF chromatin remodeling
2Present address:Cold Spring Harbor Laboratory, Cold Spring
Har-during T-lymphocyte activation (Zhaoet al.1998).
Mam-bor, NY 11724-2203.
malian PKC pathways regulate diverse cellular processes 3Corresponding author:Department of Microbiology and
Immunol-including cell cycle progression, differentiation, growth
ogy, SUNY Downstate Medical Center, 450 Clarkson Ave., Box 44,
Brooklyn, NY 11203. E-mail: [email protected] control, and tumor promotion (Livneh andFishman
1997). Similarly, the single isozyme of protein kinase C TABLE 1 in Saccharomyces cerevisiaecontrols a variety of cellular
S. cerevisiaestrains processes such as cell cycle progression, mating,
nutri-ent sensing, and the structural organization of the cy- Strain Genotype toskeleton (Heinischet al.1999). Pkc1p is activated by
BLY1 MAT␣ura3-52 his3-⌬200 lys2-801 a small GTPase of the Rho family, Rho1p, which receives
BLY49 MATasth1-3ts ura3-52 his3-⌬200 ade2-101 upstream signals from the Slg1p and Mid2p
transmem-BLY76 MATaura3-52 his3-⌬200 ade2-101 lys2-801 brane sensors (Philip and Levin 2001). The down- BLY452 MAT␣slg1⌬ura3-52 his3-⌬200 lys2-801 streamPKC1-mitogen-activated protein kinase (MAPK) BLY453 MATasth1-3ts slg1⌬ura3-52 his3-⌬200 ade2-101 cascade, consisting of Bck1p-Mkk1p/Mkk2p-Mpk1p, BLY454 MAT␣mid2⌬ura3-52 his3-⌬200 lys2-801
BLY455 MATasth1-3ts mid2⌬ura3-52 his3-⌬200 ade2-101 phosphorylates transcription factors that regulate cell
wall and cytoskeleton integrity in polarized cell growth (Heinischet al.1999). Although less well defined,
sev-eral studies report evidence for pathways that branch library cloned into the YEp24 multicopy vector (Carlsonand
Botstein1982) and selected for uracil prototrophy on SC-Ura at Pkc1p (DelleyandHall 1999;Ketelaet al.1999;
medium at 30⬚for 2 days. Approximately 75,000 independent AndrewsandStark2000;Liet al.2000;Nanduriand
Ura⫹transformants were subsequently replica printed to SC-Tartakoff2001).
Ura and grown at 37⬚ for 5 days. Twenty-eight colonies that Despite progress in characterizing the remodeling conferred growth at 37⬚ were isolated. Library clones were mechanism of purified RSC, the cellular processes in recovered from 25 of the yeast strains and retested for the suppression phenotype. Restriction endonuclease digestion which RSC functions and the chromosomal targets of
of the plasmid DNAs recovered from the yeast strains indicated RSC remain largely unknown. In this study, we carried
that 13 plasmids containedSTH1sequences; the remaining out a genetic screen for multicopy suppressors of the 12 clones represented suppressors in eight different groups. sth1-3tscell cycle mutation to identify targets that would To identify the genes responsible for the suppression pheno-define the pathways in which Sth1p and RSC are in- type, candidate genes encoded by the suppressor plasmids were amplified by PCR, subcloned into YEp24, and retested volved. The identification of two related cell wall
integ-for suppression of thesth1-3tsts phenotype. The two strongest rity sensors that function upstream of protein kinase C
suppressors, pMID2 and pSLG1, were pursued in this study. (PKC1) as suppressors revealed a functional connection Flow cytometry:Cells were grown in SC selective medium to the PKC1-dependent signal transduction pathway. at 30⬚to midlog phase, split, and diluted into prewarmed 30⬚ or 37⬚SC-Ura media at concentrations ofⵑ4⫻106cells/ml. Together, analysis of the rsc mutant phenotypes and
Shifted cells were fixed in 70% ethanol, stained with propid-the identities of suppressors and profiles of suppression
ium iodide, and their fluorescence intensities were measured suggest that RSC function is required for regulating the as described (
Duet al.1998).
cellular cytoskeleton. Actin staining:Midlog phase cells carrying plasmids grown in SC selective medium at 25⬚were diluted and shifted into 37⬚ prewarmed SC media. Shifted cells were fixed in 3.7% formaldehyde in 100 mmpotassium phosphate buffer (pH 6.5) MATERIALS AND METHODS and stained with 1.2 units of rhodamine phalloidin (Molecular Probes, Eugene, OR) for 2 hr at room temperature (Adams
Yeast strains, media, and standard methods:AllS. cerevisiae and Pringle1984). Stained cells were washed three to five strains were isogenic to S288c and are listed in Table 1. Yeast times in PBS (137 mmNaCl, 2.7 mmKCl, 4.3 mmNa2HPO4, cultures were grown in rich media, consisting of yeast extract, 1.4 mmKH2PO4). Fluorescence images were acquired using peptone, and 2% dextrose (YPD), or selective synthetic com- a KAI-2092M cooled CCD camera (Diagnostic Instruments) plete (SC) media containing 2% dextrose (GuthrieandFink with Spot software on a Nikon Labophoto-II microscope.
1991). Yeast transformations were carried out by the lithium RNA analysis:Total RNA was prepared by glass bead disrup-acetate procedure (Itoet al.1983). The plasmids and oligonu- tion as described previously (Laurentet al.1990).32P-labeled cleotides used in this study are listed in Tables 2 and 3, respec- hybridization probes were generated by PCR amplification
tively. using primer pairs MID2 5⬘and MID2 3⬘, SLG1 5⬘and SLG1
A one-step PCR gene disruption method was used to gener- 3⬘, ACT1-up and ACT1-down, and 5upTUB1 and 3dwTUB1 ate mid2⌬, slg1⌬, mid2⌬ sth1-3ts, orslg1⌬ sth1-3tsstrains as as described (Duet al.1998). RNA levels were quantified with described (Baudinet al.1993). The coding sequences ofSLG1 ImageQuant software.
or MID2were replaced with the HIS3 gene amplified from pRS313 using oligos SLG1-up and SLG1-down or MID2-up
and MID2-down, respectively. The resulting PCR products, RESULTS containing theHIS3gene flanked by 40 nucleotides of the 5⬘
and 3⬘ noncoding sequences of SLG1 or MID2 were trans- Isolation ofSLG1/WSC1andMID2as multicopy
sup-formed into the wild-typeSTH1(BLY1) andsth1-3ts(BLY49) pressors of the sth1-3ts mutant: To better understand strains to generate BLY452 and BLY453 or BLY454 and the cellular function of the RSC complex, we sought to BLY455, respectively. Deletion ofSLG1orMID2was verified
identify gene dosage suppressors ofsth1-3ts, a tempera-by PCR analysis using primersⵑ100 bp upstream of theSLG1
ture-sensitive mutation inSTH1, which encodes the es-orMID2genes and within theHIS3gene.
sential ATPase subunit. sth1-3ts mutants arrest in the Identification of high-copy extragenic suppressors of
nonpermis-TABLE 2
Plasmids
Name Description/relevant markers Source/reference
p9-YEp24 MID2in YEp24 (2,URA3) This study
p15-YEp24 SLG1in YEp24 (2,URA3) This study
pMID2 BamHI-StuI fragment containing theMID2gene generated This study by PCR amplification of p9-YEp24 primed by oligos
MID2-5BamHI and MID2-3StuI replaced theBamHI-StuI fragment of YEp24
pSLG1 BamHI-StuI fragment containing theSLG1gene generated This study by PCR amplification of p15-YEp24 primed by oligos
SLG1-5BamHI and SLG1-3StuI replaced theBamHI-StuI fragment of YEp24
pPKC1 PKC1in pSEY18 (2,URA3) Helliwellet al.(1998)
pMKK1 MKK1in YEp352 (2,URA3) Helliwellet al.(1998)
pMPK1 MPK1in YEp352 (2,URA3) Helliwellet al.(1998)
pBCK1-20 BCK1-20in pRS316 (CEN6,URA3) Helliwellet al.(1998) pROM2a ROM2in YEplac195 (2,URA3) Helliwellet al.(1998) pRHO2b RHO2in YEp352A (2,URA3) Helliwellet al.(1998)
pBL50 STH1in YEp24 (2,URA3) Laurentet al.(1992)
pRS313 CEN6, HIS3, Ampr SikorskiandHieter(1989)
YEp24 2,URA3, Ampr HurleyandDonelson(1980)
aOriginal name, pAS30. bOriginal name, pC-186.
sive temperature (37⬚) and exhibit a variety of cellular lated several genes that when overexpressed rescued the ts lethal phenotype of the mutant at 37⬚. Here we defects (Duet al.1998). We reasoned that the isolation
of gene suppressors that could restore growth ofsth1-3ts report on the two strongest suppressors. DNA sequence analysis identified a single clone containing SLG1/ cells at 37⬚would identify factors that interact with RSC
and/or uncover the cellular processes that require RSC WSC1/HCS77(synthetic lethal with Gap 1/cell wall in-tegrity and stress response component 1) and three function.
To screen for multicopy suppressors, the sth1-3ts independent clones containing MID2 (mating phero-mone-induced death 2; Figure 1). The encoded pro-strain was transformed with a yeast genomic DNA library
cloned into YEp24 (CarlsonandBotstein 1982). In teins, Slg1p and Mid2p, are plasma membrane-associ-ated proteins with overlapping functions as upstream addition to the expected wild-typeSTH1gene, we
iso-TABLE 3
Oligonucleotides
Oligonucleotide Sequence (5⬘to 3⬘)
SLG1-5BamHI CGGGATCCGCTGCAACTTTGGCATTCTA
SLG1-3StuI AAGGCCTTGTTCCTCGTTACTTCCAGC
MID2-5BamHI CGGGATCCGGCCGTCGCGCTCTACG
MID2-3StuI AAGGCCTGCTTCCCTGAAATTCTCCTCC
MID2-up TTCGTTGAAGATTGGACATATAAAATACGCAAATCATAGTCGTTTTAAGAGCTTGGTGAG
MID2-down GAAAAGTAGCCATAAGCACTAAATGATATGAATGGATATGGTGTCACTACATAAGAACAC
SLG1-up GGCTGATTTAGTACTCAGGATAAAAAATTCTATTTAAATACGTTTTAAGAGCTTGGTGAG
SLG1-down GGCAATAGTTTAAGAATATAATAATTTTTTTTGGGTTTCTGTGTCACTACATAAGAACAC
MID2 5⬘ CAACCAAGAATAGTTTCCGC
MID2 3⬘ GGAGGTTGAAGGTGCAGAAG
SLG1 5⬘ CCATTTTATCGCAAGCGAAC
SLG1 3⬘ CCGCCATTTATGAATGTTCC
ACT1-up GGTATCATGGTCGGTATGGG
ACT1-down ATGGACCACTTTCGTCGTATTC
5upTUB1 TACAACTGCAAACAAACAATGAGA
3dwTUB1 CTAAATCCACGTAAATAGCCCTTG
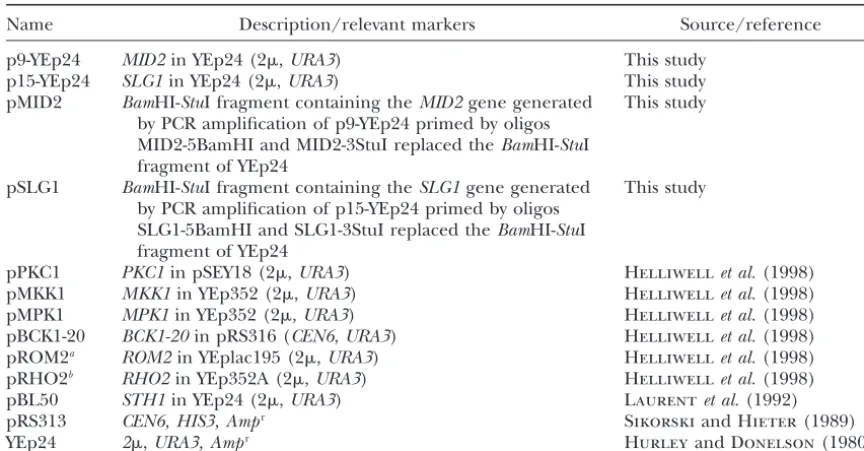
Figure1.—High-copyMID2orSLG1suppresses the temper-ature-sensitive growth ofsth1-3tsmutants. Tenfold serial dilu-tions of midlogarithmic phasesth1-3ts(BLY49) cells harboring pBL50 (STH1), YEp24 (vector), pMID2 (MID2), or pSLG1 (SLG1) were plated onto SC plates to determine growth at
permissive (25⬚) or nonpermissive (37⬚) temperatures. The Figure2.—sth1-3tsmutants exhibit phenotypes characteris-plates were photographed after 3–4 days of growth. tic ofPKC1pathway mutants. (A)sth1-3tsmutants show osmo-remedial phenotypes. Threefold serial dilutions of log phase sth1-3ts(BLY49) and isogenic wild-type STH1(BLY76) cells were spotted onto YPD plates or YPD plates containing 1 m
cell wall stress sensors that activate thePKC1-MAP kinase
sorbitol and growth was compared at 37⬚. (B)sth1-3tscells are pathway (Grayet al.1997;Vernaet al.1997;Jacobyet sensitive to caffeine and staurosporine and these sensitivities al.1998;Rajavelet al.1999;StirlingandStark2000). are suppressed byMID2orSLG1. Threefold serial dilutions of log phasesth1-3ts(BLY49) cells carrying the indicated plasmids Interestingly, overexpression of eitherSLG1orMID2
were plated onto YPD or YPD containing caffeine (6 mm) or can also suppress the temperature sensitivity conferred
staurosporine (5 g/ml) and growth was compared at 30⬚ by a different STH1 allele, sth1-1ts (data not shown). after 3 days (caffeine) or 5 days (staurosporine).
However, neither gene can suppresssfh1-1::HIS3, a mu-tant allele of a second well-conserved RSC component,
Sfh1p. High-copySLG1orMID2also failed to suppress denet al.1997;Heinischet al.1999). For example, the ssn20-1, a mutant allele of a chromatin-associated pro- caffeine sensitivity ofslg1⌬mutants can be suppressed tein, Spt6p (data not shown). These results indicate that by overexpression of PKC1 or RHO1 (Ketela et al. thesth1-3tssuppressors are gene specific. 1999). Thus, several key features of pkc1pathway
mu-sth1-3ts mutants display phenotypes characteristic of tants are shared bysth1-3tsmutants.
PKC1 pathway mutants: Pkc1p plays a key regulatory STH1interacts with upstream activators of thePKC1
(Mad-Figure4.—Thesth1-3tsmutation interacts synthetically with theslg1mutation. Wild-type (BLY1),sth1-3ts(BLY49), slg1⌬ (BLY452),sth1-3ts slg1⌬(BLY453),mid2⌬(BLY454), and sth1-3ts mid2⌬ (BLY455) cells were grown to midlog phase, 10-fold serially diluted, and spotted on YPD plates. Growth was compared at 30⬚and 36⬚after 2 days.
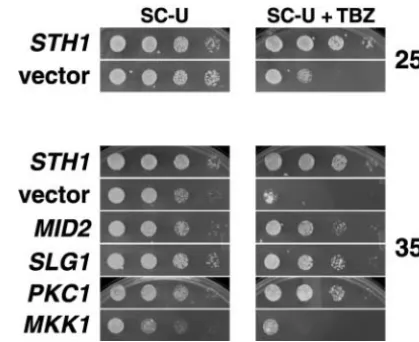
Figure3.—PKC1and its upstream activators,MID2,SLG1, ROM2, andRHO2, but not the downstream effector kinases MKK1orMPK1, suppress thesth1-3tstemperature sensitivity.
sth1-3ts cell cycle arrest (data not shown). This result Tenfold serial dilutions of log phase sth1-3ts (BLY49) cells
suggests that the alternative PKC1-mediated signaling carrying the indicated high-copy plasmids were plated onto
SC plates to determine growth at 25⬚or 37⬚. (RHO2was tested pathway participates in the cell cycle function of RSC. becauseRHO1in high copy can be toxic;AndrewsandStark Delocalization of the cortical actin cytoskeleton insth1
2000.) All plates were photographed after 2–3 days of growth.
mutants can be suppressed by high-copyMID2,SLG1, or
PKC1:The established role of Pkc1p in mediating the cell cycle-dependent organization of actin (Helliwell the interaction between RSC and the upstream activa- et al.1998) implicates RSC inPKC1-dependent signaling tors but not the downstream effectors of thePKC1-MAP pathways that control the cortical actin cytoskeleton. To kinase pathway suggests that RSC functions in alternate assess the integrity and distribution of the actin cytoskel-PKC1-mediated pathways. eton, we first visualized intracellular polymerized actin
Deletion ofSLG1but notMID2is synthetically lethal filaments by staining sth1-3ts cells carrying wild-type with thesth1-3ts mutation:To further characterize the STH1or vector alone grown at 25⬚ or 37⬚with rhoda-genetic connection between RSC and the upstream mine-coupled phalloidin. In mutants carrying vector PKC1 pathway, double mutants between sth1-3ts and grown at permissive temperature (25⬚), both cortical deletions of the cell wall integrity signaling genesSLG1 actin patches and cable arrays were polarized in cells or MID2 were constructed. The sth1-3ts mutation was throughout the cell cycle in a pattern that was indistin-found to be lethal at the semipermissive temperature guishable from that insth1-3tscells expressing wild-type (36⬚) in combination with theslg1⌬mutation (Figure STH1at 25⬚or 37⬚(Figure 6A). In contrast, upon shift-4). In contrast, the growth phenotypes of the sth1-3ts ing mutants harboring vector alone to 37⬚, the majority mid2⌬double mutants were no different from those of of cells arrested with large, misshapen buds and actin the single mutants. These data distinguish the genetic patches were dispersed randomly throughout both interactions betweenSTH1and the cell wall stress sen- mother and daughter cells. Most of these cells also con-sors SLG1or MID2. Furthermore, the ability of MID2 tained unusual large actin clumps. Actin cables were not to suppresssth1-3ts orslg1⌬ single mutants but not an visible in these mutants, and in cells that had completed sth1-3ts slg1⌬double mutant (data not shown) suggests nuclear division, actin rings were never seen at the bud that Sth1p and Slg1p function in genetically redundant necks.
or parallel pathways and is in agreement with the previ- We next investigated whether the high-copy suppres-ous study that SLG1 and MID2 have overlapping and sors could correct the delocalization of the cortical actin distinct functions (StirlingandStark2000). cytoskeleton in thesth1-3tscells at 37⬚. In mutants car-PKC1 and its upstream activators relieve the G2/M ryingMID2orSLG1, we detected polarized actin rings
arrest ofsth1-3ts cells:The sth1-3tsmutation confers a at the bud necks of large-budded cells and polarized G2/M cell cycle arrest (Duet al. 1998). The ability of actin patches and cable arrays in small- or medium-sized SLG1,MID2, orPKC1to suppress the ts growth defects buds (Figure 6A). In addition, the unusual actin clumps caused bysth1-3ts prompted us to determine whether characteristic of the mutants were seen only rarely. these genes improved growth by relieving these cells High-copyPKC1also partially corrected the depolarized from G2/M arrest. Indeed, high-copyMID2,SLG1, and actin patches in sth1-3ts cells, although less effectively PKC1could partially suppress the G2/M arrest pheno- than did MID2or SLG1(data not shown).
of ACT1, the sole actin gene in yeast, and TUB1, an ␣-tubulin gene, was largely unaffected (Figure 6B). Thus, RSC appears to only mildly affect transcription of the genes that act in the alternative PKC1 effector pathway or the downstream cytoskeletal target genes.
MID2 or PKC1 partially suppresses the sth1 defects in mating projection formation:Remodeling of the actin cytoskeleton is required for polarized cell growth in mitotically growing cells and projection formation dur-ing matdur-ing. The known involvement of Pkc1p signaldur-ing in actin cytoskeletal rearrangements prompted us to investigate pheromone-induced projection formation in the sth1-3ts cells (Errede et al.1995; Zarzov et al. 1996; Buehrer and Errede 1997). Following a 3-hr exposure to␣-factor, 87% ofsth1-3tscells carrying wild-type STH1 formed mating projections while only 5% were found in S or G2/M cell cycle stages, consistent with
Figure 5.—MID2 suppresses the sth1-3ts G2/M cell cycle
arrest.sth1-3ts(BLY49) cells carrying pBL50 (STH1), YEp24 results in wild-type cells. In contrast, 34% of ␣ -factor-(vector), or pMID2 (MID2) were analyzed by flow cytometry treatedsth1-3tsmutants carrying vector alone were still 12 hr after being shifted from 30⬚to 37⬚.
found in S or G2/M cell cycle stages and were therefore defective in responding to pheromone-induced late G1 arrest; untreated wild-type and mutant cells showed simi-requiresSTH1 function. However, we found that RNA lar cellular distributions (Table 4). Significantly, 92% levels ofMID2insth1-3tscells grown at 37⬚were compa- (87/87 ⫹ 8) of the unbudded sth1-3ts cells carrying rable to those in wild-type cells shifted to 37⬚, and levels STH1 formed mating projections or “shmoos,” com-of SLG1 were, on average, only twofold lower (Figure pared to only 29% (19/19 ⫹ 47) of the sth1-3ts cells 6B; data not shown). Alternatively, expression of genes carrying vector. (The percentage of shmoo formation that encode components of the cytoskeleton could be is expressed as the fraction of cells forming shmoos in the unbudded population.) Moreover, compared to affected by loss ofSTH1function. Again, transcription
TABLE 4
MID2orPKC1partially suppresses thesth1-3tsmating projection formation defect
% of cells in the phase shown
Plasmid G1 S→G2/M Shmoo
No␣-factor
STH1 52 48 0
Vector 49 51 0
␣-Factor
STH1 8 5 87
Figure7.—MID2,SLG1, orPKC1can suppress the
hypersen-Vector 47 34 19
sitivity of sth1-3ts to the microtubule-depolymerizing drug
MID2 34 21 45
TBZ. Midlog phase sth1-3ts (BLY49) cells, carrying pBL50
PKC1 41 20 39
(STH1), YEp24 (vector), pMID2 (MID2), pSLG1 (SLG1), pPKC1 (PKC1), or pMKK1 (MKK1) were 10-fold serially di-Aliquots of logarithmic phasesth1-3ts(BLY49) cells
harbor-luted, spotted on SC-Ura plates or SC-Ura plates containing ing pBL50 (STH1), YEp24 (vector), pMID2 (MID2), or pPKC1
60g/ml TBZ, and incubated at 25⬚or 35⬚for 3–4 days. (PKC1) were diluted to OD600⫽0.15 prior to (no␣-factor)
or 3 hr following treatment with␣-factor (7m) and analyzed microscopically to determine the cell cycle stage (n⬎200). Percentages of cells in G1, S or G2/M (combined), or those
Thesth1mutants shifted to the semipermissive tempera-forming mating projections (shmoos) are indicated.
ture (35⬚) were also sensitive to TBZ (Figure 7). Interest-ingly, like suppression of the ts and cell cycle pheno-types,PKC1and its upstream activators,MID2andSLG1, mutants expressingSTH1,sth1-3ts cells carrying vector
also suppressed the TBZ growth inhibition of sth1-3ts alone have a higher proportion of cells in G1 (47vs.
mutants, but only at 35⬚. DownstreamPKC1-MAP kinase 8%) and S→ G2/M (34 vs. 5%), suggesting that the
module effectors failed to suppress the TBZ sensitivity, mutants are defective either in the remodeling of the
suggesting that the link to microtubule function is also actin cytoskeleton required for shmoo formation or in
via the alternativePKC1pathway. Suppression at 35⬚but the cortical patch-dependent endocytosis critical for
not 25⬚ supports the notion that this alternativePKC1 mating signal transduction.
pathway is more active at higher temperature. The suppression of the actin cytoskeletal defects in
budding sth1-3ts cells by Mid2p, Slg1p, or Pkc1p and the known involvement of these proteins in polarized
DISCUSSION growth for mating prompted us to test whether any of
these proteins could rescue the mutant insensitivities Here, we have uncovered a link between Sth1p, the to pheromone. We testedPKC1 and MID2and found essential Snf2p-related ATPase component of RSC, and that either partially restored the response to mating the Pkc1p-dependent signaling pathway. Our genetic pheromone insth1-3tsmutants: 49% of mutants carrying analysis indicates thatsth1 interacts specifically with an PKC1 and 57% of mutants carrying MID2 formed alternative PKC1 pathway that shares upstream activa-shmoos. Additionally, fewer of these cells accumulated tors with thePKC1-MAP kinase pathway. Moreover, we in either G1 or S→G2/M. propose a model that this alternative pathway regulates
Thesth1 mutant sensitivity to the microtubule depo- the organization of the cellular cytoskeleton since this
lymerizing agent TBZ is corrected by high-copy suppres- pathway rescues thesth1-3tsmutant defects in cortical
sors:Although a functional connection betweenPKC1 actin polarization and its sensitivity to TBZ. Thus, our and the actin cytoskeleton is more firmly established, data provide important new insight into the cellular several reports indicate thatPKC1signaling is also func- pathways in which RSC function is required.
tionally linked to the microtubule cytoskeleton (Man- RSC functions through an alternative PKC1
death (Mid) phenotype ofmid2⌬and the ts phenotype tion, the ability of the majority ofsth1-3tscells to arrest in G1 in the presence of mating pheromone suggests ofglc7-10, an allele of a type 1 protein phosphatase, can
be suppressed only by overexpression of PKC1 or its that Far1p is still capable of inhibiting Cdc28p activity, which allows cells to enter S phase. Thus, we infer that upstream activators (Ketelaet al.1999;Andrewsand
Stark 2000). Likewise, the transcriptional repression downstream events required for actin cytoskeletal re-arrangements are defective insth1-3tsmutants.
of ribosome synthesis and a novel arrest of secretion
response in secretion-defective cells require only up- Several lines of evidence also link RSC to microtubule-dependent processes viaPKC1-dependent pathways. In stream elements of the PKC1 pathway (Li et al.2000;
NanduriandTartakoff2001). Interestingly, suppres- this study, we showed thatsth1-3tscells exhibited hyper-sensitivity to TBZ, a microtubule depolymerizing agent, sion of the sth1-3ts G2/M arrest, actin delocalization,
and TBZ sensitivity also distinguished upstream from and that this sensitivity was suppressed by upstream PKC1activators, includingMID2. Previously,MID2was downstream PKC1-MAPK components. Furthermore,
high-copyMID2 can no longer suppress thesth1-3tsts shown to suppress mutations in Kar3p and Cik1p, two interacting proteins that regulate spindle pole body phenotype when thePKC1pathway function is
compro-mised by deletion of SLG1, indicating thatPKC1func- (SPB) and microtubule function (Manninget al.1997; Barrett et al. 2000). A PKC1-mediated pathway has tion is required for sth1-3tssuppression. Thus, several
of thesth1-3tsdefects are suppressed only byPKC1and recently been shown to interact with components of the SPB as well (Khalfanet al.2000). In addition, an its upstream activators, strongly suggesting that RSC
functions through an alternativePKC1-mediated signal- activator of the Rom2p exchange factor, Tor2p, trans-mits signals to the actin cytoskeleton and has been impli-ing pathway, although we cannot rule out the possibility
that RSC partly signals through the downstream MAP cated in regulating microtubule structure and function (Choi et al. 2000). Furthermore, the G2/M arrest of kinase cascade.
RSC function is necessary for cytoskeletal organiza- tworscmutants has been shown to engage theMAD1 -dependent spindle-assembly checkpoint (Tsuchiya et
tion:Our analysis ofsth1-3tscells has revealed a
require-ment for RSC function in actin cytoskeletal rearrange- al. 1998;Angus-Hill et al.2001), a pathway that can be activated by defects in microtubule structure and ments necessary for polarized growth. First, the actin
cytoskeleton is delocalized in mitotically arrestingsth1- function. More importantly, both the actin delocaliza-tion and TBZ sensitivity of sth1-3ts can be rescued by 3ts cells at 37⬚. Second, sth1-3ts cells are defective in
forming mating projections in the presence of mating PKC1and its upstream activators. Taken together, these data strongly suggest a requirement for RSC in actin pheromone, even at 30⬚.
The delocalized cortical actin patches observed in cytoskeleton organization and microtubule function via aPKC1-dependent signaling pathway, thus linking nu-mitotically growing sth1-3tscells at 37⬚ have also been
seen inpfy1⌬andrho1-2mutants (Helliwellet al.1998; clear chromatin remodeling activities with intracellular signaling.
Marcoux et al. 2000). PFY1 encodes profilin, a small
actin-binding protein proposed to function downstream Similar to the functional connection between hSWI/ SNF and PKC/ras signaling, here we have demonstrated of Rho1p. In addition, the unusual large actin clumps
characteristic of sth1-3ts cells grown at the restrictive a link between RSC and a PKC1-dependent pathway. Although Pkc1p regulates diverse cellular processes, we temperature have also been observed in sla1 and in
nhp6A⌬nhp6B⌬double mutants (Holtzmanet al.1993; show that RSC participates specifically in an alternative PKC1pathway. Recent studies indicate that this alterna-Costiganet al.1994). Sla1p participates in cortical actin
cytoskeleton assembly, and Nhp6A and Nhp6B are re- tive pathway may regulate interorganellar signaling from the secretory path to the nucleus (Nanduriand dundant HMG1-like DNA-binding proteins that
func-tion downstream of thePKC1-MAP kinase pathway. The Tartakoff2001). Interestingly, in an independent ge-netic screen, we recovered mutations in a secretory fac-inappropriate cortical patch distribution insth1-3tscells
resembles that in mutants that affect aspects of actin tor as extragenic suppressors ofsth1-3tsmultiple times (J. Duand B. C. Laurent, unpublished results). Fur-cytoskeletal dynamics, implicating RSC in related
actin-dependent functions. thermore, defects in the secretory pathway can result
in transcriptional repression of the genes encoding ribo-A key event in polarizing the actin cytoskeleton in
both mitotic growth and shmoo formation is the local- somal proteins (RPs;Warner1999), and recently, the expression profiles of RP genes in tworscmutants were ization of the Rho-GTPase Cdc42p to selected growth
sites.sth1-3tsmutants grown at the permissive tempera- shown to be altered (Angus-Hillet al.2001). Our data connect RSC function to the actin cytoskeleton via the ture are capable of normal polarized mitotic cell growth
but limited pheromone-induced polarized growth, sug- alternativePKC1pathway. It will be important to exam-ine expression levels of candidate target genes,e.g., RP gesting that only the cell polarity signaling pathway
unique to pheromone induction is defective at permis- genes, cell wall integrity genes, and secretory pathway genes. The connection between the alternativePKC1and sive temperature. Defects in the recruitment of Cdc42p
tions with histones and chromatin-associated proteins. Genetics
model and supports the idea that RSC function is required
150:987–1005.
for regulating actin-dependent secretion through this al- Errede, B., R. M. Cade, B. M. Yashar, Y. Kamada, D. E. Levinet al.,
1995 Dynamics and organization of MAP kinase signal pathways.
ternativePKC1 pathway.
Mol. Reprod. Dev.42:477–485.
We thank Pierre-Alain Delley for providing plasmids, Mara Amoros Fry, C. J., andC. L. Peterson, 2001 Chromatin remodeling en-for assistance with RNA analysis, and members of the Laurent labora- zymes: who’s on first? Curr. Biol.11:R185–197.
tory for helpful comments on the manuscript. This work was supported Gray, J. V., J. P. Ogas, Y. Kamada, M. Stone, D. E. Levinet al., 1997 A role for the Pkc1 MAP kinase pathway ofSaccharomyces cerevisiae
by Public Health Service grant GM56700 from the National Institutes
in bud emergence and identification of a putative upstream regu-of Health to B.C.L.
lator. EMBO J.16:4924–4937.
Guthrie, S., and G. R. Fink, 1991 Guide to yeast genetics and molecular biology. Methods Enzymol.194:12–17.
Heinisch, J. J., A. Lorberg, H.-P. SchmitzandJ. J. Jacoby, 1999
LITERATURE CITED The protein kinase C-mediated MAP kinase pathway involved in
the maintenance of cellular integrity inSaccharomyces cerevisiae.
Adams, A. E., and J. R. Pringle, 1984 Relationship of actin and
Mol. Microb.32:671–680. tubulin distribution to bud growth in wild-type and
morphoge-Helliwell, S. B., A. Schmidt, Y. OhyaandM. N. Hall, 1998 The netic-mutant Saccharomyces cerevisiae. J. Cell Biol.98:934–945.
Rho1 effector Pkc1, but not Bni1, mediates signalling from Tor2
Andrews, P. D., andM. J. R. Stark, 2000 Type 1 protein
phospha-to the actin cyphospha-toskelephospha-ton. Curr. Biol.8:1211–1214. tase is required for maintenance of cell wall integrity,
morphogen-Holtzman, D. A., S. YangandD. G. Drubin, 1993 Synthetic-lethal esis and cell cycle progression in Saccharomyces cerevisiae. J.
interactions identify novel genes, SLA1 and SLA2, that control Cell Sci.113:507–520.
membrane cytoskeleton assembly in Saccharomyces cerevisiae. J.
Angus-Hill, M. L., A. Schlichter, D. Roberts, H. Erdjument
-Cell Biol.122:635–644.
Bromage, P. Tempst et al., 2001 A Rsc3/Rsc30 zinc cluster
Hurley, J. L., andJ. E. Donelson, 1980 Nucleotide sequence of dimer reveals novel roles for the chromatin remodeler RSC in
the yeast plasmid. Nature286:860–865. gene expression and cell cycle control. Mol. Cell7:741–751.
Ito, H., Y. Fukuda, K. MurataandA. Kimura, 1983 Transformation
Banuett, F., 1998 Signalling in the yeasts: an informational cascade
of intact yeast cells treated with alkali cations. J. Bacteriol.153: with links to the filamentous fungi. Microb. Mol. Biol. Rev.62:
163–168. 249–274.
Jacoby, J. J., S. M. NiliusandJ. J. Heinisch, 1998 A screen for
Barrett, J. G., B. D. ManningandM. Snyder, 2000 The Kar3p upstream components of the yeast protein kinase C signal trans-kinesin-related protein forms a novel heterodimeric structure
duction pathway identifies the product of theSLG1gene. Mol. with its associated protein Cik1p. Mol. Biol. Cell11:2373–2385. Gen. Genet.258:148–155.
Baudin, A., O. Ozier-Kalogeropoulos, A. Denouel, F. Lacroute Jenuwein, T., andC. D. Allis, 2001 Translating the histone code.
andC. Cullin, 1993 A simple and efficient method for direct Science293:1074–1080.
gene deletion inSaccharomyces cerevisiae.Nucleic Acids Res.21: Ketela, T., R. GreenandH. Bussey, 1999 Saccharomyces cerevisiae
3329–3330. Mid2p is a potential cell wall stress sensor and upstream activator
Berger, S. L., 2001 Molecular biology. The histone modification of thePKC1–MPK1cell integrity pathway. J. Bacteriol.181:3330–
circus. Science292:64–65. 3340.
Berger, S. L., and G. Felsenfeld, 2001 Chromatin goes global. Khalfan, W., I. IvanovskaandM. D. Rose, 2000 Functional
interac-Mol. Cell8:263–268. tion between the PKC1 pathway and CDC31 network of SPB
Buehrer, B. M., andB. Errede, 1997 Coordination of the mating duplication genes. Genetics155:1543–1559.
and cell integrity mitogen-activated protein kinase pathways in Kingston, R. E., andG. J. Narlikar, 1999 ATP-dependent
remodel-Saccharomyces cerevisiae.Mol. Cell. Biol.17:6517–6525. ing and acetylation as regulators of chromatin fluidity. Genes
Cairns, B. R., Y. Lorch, Y. Li, M. Zhang, L. Lacomiset al., 1996 Dev.13:2339–2352.
RSC, an essential, abundant chromatin-remodeling complex. Cell Kwon, H., A. N. Imbalzano, P. A. Khavari, R. E. Kingstonand
87:1249–1260. M. R. Green, 1994 Nucleosome disruption and enhancement
Cairns, B. R., H. Erdjument-Bromage, P. Tempst, F. Winstonand of activator binding by a human SWI/SNF complex. Nature370:
R. D. Kornberg, 1998 Two actin-related proteins are shared 477–481.
functional components of the chromatin-remodeling complexes Laurent, B. C., M. A. TreitelandM. Carlson, 1990 The SNF5 RSC and SWI/SNF. Mol. Cell2:639–651. protein of Saccharomyces cerevisiae is a glutamine- and
proline-Cairns, B. R., A. Schlichter, H. Erdjument-Bromage, P. Tempst, rich transcriptional activator that affects expression of a broad R. D. Kornberget al., 1999 Two functionally distinct forms of spectrum of genes. Mol. Cell. Biol.10:5616–5625.
the RSC nucleosome-remodeling complex, containing essential Laurent, B. C., X. YangandM. Carlson, 1992 An essential Saccha-AT hook, BAH, and bromodomains. Mol. Cell4:715–723. romyces cerevisiaegene homologous toSNF2encodes a
helicase-Cao, Y., B. R. Cairns, R. D. KornbergandB. C. Laurent, 1997 related protein in a new family. Mol. Cell. Biol.12:1893–1902. Sfh1p, a component of a novel chromatin-remodeling complex, is Lee, K. S., andD. E. Levin, 1992 Dominant mutations in a gene required for cell cycle progression. Mol. Cell. Biol.17:3323–3334. encoding a putative protein kinase (BCK1) bypass the
require-Carlson, M., andD. Botstein, 1982 Two differentially regulated ment for aSaccharomyces cerevisiaeprotein kinase C homolog. Mol. mRNAs with different 5⬘ends encode secreted and intracellular Cell. Biol.12:172–182.
forms of yeast invertase. Cell28:145–154. Levin, D. E., andE. Bartlett-Heubusch, 1992 Mutants in theS.
Choi, J. H., N. R. Adames, T.-F. Chan, C. Zeng, J. A. Cooperet cerevisiae PKC1gene display a cell cycle-specific osmotic stability
al., 2000 TOR signaling regulates microtubule structure and defect. J. Cell Biol.116:1221–1229.
function. Curr. Biol.10:861–864. Levin, D. E., F. O. Fields, R. Kunisawa, J. M. BishopandJ. Thorner,
Costigan, C., D. KolodrubetzandM. Snyder, 1994 NHP6A and 1990 A candidate protein kinase C gene,PKC1, is required for NHP6B, which encode HMG1-like proteins, are candidates for theS. cerevisiaecell cycle. Cell62:213–224.
downstream components of the yeast SLT2 mitogen-activated Li, Y., R. D. Moir, I. K. Sethy-Coraci, J. R. WarnerandI. M. Willis, protein kinase pathway. Mol. Cell. Biol.14:2391–2403. 2000 Repression of ribosome and tRNA synthesis in
secretion-Coˆ te´, J., J. Quinn, J. L. WorkmanandC. L. Peterson, 1994 Stimu- defective cells is signaled by a novel branch of the cell integrity lation of GAL4 derivative binding to nucleosomal DNA by the pathway. Mol. Cell. Biol.20:3843–3851.
yeast SWI/SNF complex. Science265:53–60. Livneh, E., andD. D. Fishman, 1997 Linking protein kinase C to
Delley, P. A., andM. N. Hall, 1999 Cell wall stress depolarizes cell cell-cycle control. Eur. J. Biochem.248:1–9.
growth via hyperactivation of RHO1. J. Cell Biol.147:163–174. Madden, K., Y. J. Sheu, K. Baetz, B. AndrewsandM. Snyder, 1997
Du, J., I. Nasir, B. K. Benton, M. P. KladdeandB. C. Laurent, SBF cell cycle regulator as a target of the yeast PKC-MAP kinase 1998 Sth1p, aSaccharomyces cerevisiaeSnf2p/Swi2p homolog, is pathway. Science275:1781–1784.
interac-GEF Rom2p localizes to sites of polarized growth and participates yeast host strains designed for efficient manipulation of DNA in
Saccharomyces cerevisiae.Genetics122:19–27. in cytoskeletal functions in Saccharomyces cerevisiae. Mol. Biol.
Cell8:1829–1844. Stirling, D. A., and M. J. R. Stark, 2000 Mutations in SPC110,
encoding the yeast spindle pole body calmodulin-binding
pro-Marcoux, N., S. Cloutier, E. Zakrzewska, P.-M. Charest, Y.
Bour-tein, cause defects in cell integrity as well as spindle formation.
bonnaiset al., 2000 Suppression of the profilin-deficient
pheno-Biochim. Biophys. Acta1499:85–100. type by the RHO2 signaling pathway inSaccharomyces cerevisiae.
Strahl, B. D., andC. D. Allis, 2000 The language of covalent Genetics156:579–592.
histone modifications. Nature403:41–45.
Moreira, J. M., andS. Holmberg, 1999 Transcriptional repression
Tsuchiya, E., T. HosotaniandT. Miyakawa, 1998 A mutation in of the yeast CHA1 gene requires the chromatin-remodeling
com-NPS1/STH1, an essential gene encoding a component of a novel plex RSC. EMBO J.18:2836–2844.
chromatin-remodeling complex RSC, alters the chromatin
struc-Nanduri, J., andA. M. Tartakoff, 2001 The arrest of secretion
ture ofSaccharomyces cerevisiaecentromeres. Nucleic Acids Res. response in yeast: signaling from the secretory path to the nucleus
26:3286–3292. via Wsc proteins and Pkc1p. Mol. Cell8:281–289.
Tsuchiya, E., M. Uno, A. Kiguchi, K. Masuoka, Y. Kanemoriet al.,
Nie, Z., Y. Xue, D. Yang, S. Zhou, B. J. Derooet al., 2000 A specificity
1992 TheSaccharomyces cerevisiae NPS1gene, a novelCDCgene and targeting subunit of a human SWI/SNF family-related
chro-which encodes a 160 kDa nuclear protein involved in G2phase
matin-remodeling complex. Mol. Cell. Biol.20:8879–8888.
control. EMBO J.11:4017–4026.
Papoulas, O., S. J. Beek, S. L. Moseley, C. M. Mccallum, M. Sarte
Verna, J., A. Lodder, K. Lee, A. VagtsandR. Ballester, 1997 A
et al., 1998 The Drosophila trithorax group proteins BRM, ASH1
family of genes required for maintenance of cell wall integrity and ASH2 are subunits of distinct protein complexes.
Develop-and for the stress response inSaccharomyces cerevisiae.Proc. Natl. ment125:3955–3966.
Acad. Sci. USA94:13804–13809.
Paravicini, G., M. Cooper, L. Friedli, D. J. Smith, J.-L. Carpentier
Vignali, M., A. H. Hassan, K. E. NeelyandJ. L. Workman, 2000
et al., 1992 The osmotic integrity of the yeast cell requires a
ATP-dependent chromatin-remodeling complexes. Mol. Cell. functionalPKC1gene product. Mol. Cell. Biol.12:4896–4905.
Biol.20:1899–1910.
Philip, B., andD. E. Levin, 2001 Wsc1 and Mid2 are cell surface
Warner, J. R., 1999 The economics of ribosome biosynthesis. sensors for cell wall integrity signaling that act through Rom2,
Trends Biochem. Sci.24:437–440. a guanine nucleotide exchange factor for Rho1. Mol. Cell. Biol.
Yukawa, M., S. Katoh, T. MiyakawaandE. Tsuchiya, 1999 Nps1/
21:271–280. Sth1p, a component of an essential chromatin-remodeling
com-Rajavel, M., B. Philip, B. M. Buehrer, B. ErredeandD. E. Levin, plex ofSaccharomyces cerevisiae, is required for the maximal expres-1999 Mid2 is a putative sensor for cell integrity signaling in sion of early meiotic genes. Genes Cells4:99–110.
Saccharomyces cerevisiae.Mol. Cell. Biol.19:3969–3976. Zarzov, P., C. MazzoniandC. Mann, 1996 TheSLT2(MPK1) MAP
Roemer, T., G. Paravicini, M. A. Payton andH. Bussey, 1994 kinase is activated during periods of polarized cell growth in Characterization of the yeast (1-⬎6)-b-glucan biosynthetic com- yeast. EMBO J.15:83–91.
ponents, Kre6p and Skn1p, and genetic interactions between the Zhao, K., W. Wang, O. J. Rando, Y. Xue, K. Swidereket al., 1998
PKC1pathway and extracellular matrix pathway. J. Cell Biol.127: Rapid and phosphoinositol-dependent binding of the
SWI/SNF-567–579. like BAF complex to chromatin after T lymphocyte receptor
Roth, S. Y., J. M. DenuandC. D. Allis, 2001 Histone acetyltransfer- signaling. Cell95:625–636. ases. Annu. Rev. Biochem.70:81–120.