IN DROSOPHILA PAVANP
DANK0 BRNCIC, SUS1 KOREF-SANTIBAREZ, MYRIAM BUDNIK
AND MADELEINE LAMBOROT
Uniuersity of Chile2
Received June 17, 1968
DROSOPHZLA PAVANI BRNCIC 1957 is a neotropical species belonging to the mesophragmatica group which is very abundant in central Chile and on the eastern slope 01 the Andes in Argentine. The natural populations of this species are polymorphic for the gene arrangements in two of its chromosomes. Chromosome 2 has two inversions, one included within the other, which have up to now never been found separately, that is, no individuals have been found in nature or in the laboratory which differ from the arbitrarily designed “Standard” by only one of these arrangements. In the right arm of the fourth chromosome, besides “Standard” there is an arrangement constituted by three overlapping inversions, which are always found together, and in the left arm there is another complex rearrangement which also differs from “Standard” by three overlapping inversions. Just as occurs with the gene orders in the right arm, in the left one no intermediate steps have been found between the actual complex arrangements and the “Standard” sequence (BRNCIC 1957). There is a tendency for non- random association between the arrangements in both anns of this chromosome
(BRNCIC 1961).
After 15 years of cytogenetic studies, both in natural populations as well as in laboratory stocks of D. pauani, it has been possible to establish that there are no appreciable geographic, seasonal, altitudinal o r other environmental fluctuations of the different chromosomal variants. In all the samples analyzed, the frequency of the structural heterozygotes for the genetic arrangements in the fourth chromo- some is always around 50%. In the stocks maintained in the laboratory, the different chromosomal arrangements conserve in general the same frequencies found in nature. Summarizing,
D.
paumi is a good example of a species bearing a “rigid” o r “stable” polymorphism, according to the definition given by DOBZ- HANSKY (1962), in contrast to other species in which polymorphism is of the “flexible” type, where the frequencies of the different arrangements are modified by environmental changes both in nature and under laboratory conditions (ref. in DOBZHANSKY 1961 ; CARSON 1965; SPERLICH 1967). These facts have led to the conclusion that the chromosomal polymorphism actually present inD.
pauani represents the final product of a long and continuous selective process, which has1 The work reported in this a r t d e has been partially supported by grants from the School of Medicine, Uryverslty of Departmento de Genetlca, Faculted de Medicina y Facultad de Ciencias, Unwersidad de Chile, Zaiiartu 1042, Chile, and the Comiu6n Nauonal de Investigacibn Cientifica y Technoloeca
Santiago, Chile
472 DANKO BRNCIC et al.
mainly favored the heterokaryotypes in most of the environments exploited by the species (“hetero-selection” according to CARSON 1959). As a result, the hetero- karyotypes may be expected to be heterotic, that is, superior in Darwinian Fitness. I n searching for the heterotic properties of the heterozygotes €or inversions in D. pauani it was found that they show a greater longevity (BRNCIC and DEL SOLAR
1961 )
,
aiid exhibit a greater mating activity (BRNCIC and KOREF-SANTIBA~EZ1964; KOREF-SANTIBA~~EZ and BRNCIC 1965). In the present paper the authors wish to discuss another physiological property of the heterokaryotypes in D. pawni, which may also be a good indicator of heterosis. This refers to their faster rate of development.
EXPERIMENTS A N D RESULTS
The experiments to be described were performed with a stock of D. pauani originated from flies collected in the central part of Chile (Bellavista) i n 1961.
The stock was polymorphic for the gene arrangements in its second and fourth chromosomes ( BRNCIC 195 7; BRNCIC and KOREF-SANTIBA~EZ 1964) and at the time of the present work the frequency of heterokaryotypes for the left arm of the fourth chromosome was of the order of 55
%,
and that of the right arm of the same chromosome was around 48%. The second chromosome arrangements were not considered, due tc a greater variability in the frequencies of the different gene orders.The heterogeneity of the strain was assured by the method of maintenance used. The stock was started from at least 100 females inseminated in nature, and every generation, the progeny of at least ten culture bottles was mixed, and re- distributed in another ten containers. Further details may be found in a previous paper ( BRNCIC and KOREF-SANTIBA~EZ 1964).
For the present experiments the following procedure was employed. 250 ten- day-old virgin females were allowed to mate with an equal number of males for
48 hrs in a plastic population box ( 1 0 x 2 5 ~ 1 8 cm) containing several food vials. These were renewed, and the eggs laid during 24 h r periods were collected for three consecutive days. Batches of 50 eggs were placed in vials containing equal amounts of food medium enriched with dried yeast. The same process was re- peated 3 times with new series of parent flies. The whole experiment was repli- cated 3 months later (Experiment 2). A total of 29,700 eggs was collected. The vials containing the eggs were placed in an incubator at 16°C. The first flies eclosed twenty-nine days later and all individuals emerging thereafter were sexed and recorded; the last ones hatched 65 days after the eggs had been collected. A
total of 13,936 imagoes developed.
Figure 1 shows die distribution of the flies which hatched over a period of 36
days. The mean developmental time was 41.62 days with a Standard deviation of
6.14 days. The period in which a maximum of flies emerged was at 38 days, after which the rate of emergence decreased gradually, up to the 65th day. The flies were classified into three groups according to their developmental rate: those of
18
16
-
-
Z
1 4 -
t
Q,
y
12-
E
Q,10:
-
-E
8 -
L
6 -
4 -
2 -
Q,
0
Q,
n
-
0
parrani
FAST
days
FIGURE 1.-Developmental time (from egg to adult) of 13,936 indivi,duals of rhe species
Drosophila pavani, maintained at 16°C.
From each group, adults of both sexes were mated individually to males or females of a stock homozygous for the Standard gene arrangements, the sibling species
D.
gaucha. Previous studies ( KOREF-SANTIBA~EZ 1963; BRNCIC andKOREF-SANTIBA~EZ 1964) had shown that there is no significant sexual isolation between
D.
pauani andD.
gaucha, and that they produce hybrids easily. The pairs were maintained in an incubator at 25°C until larvae appeared. The food was then enriched with a live yeast suspension and the vials were transferred to an incubator at 16°C until the larvae were ready to pupate.The salivary gland chromosomes from eight larvae of each vial were prepared by means of the aceto-orcein rapid squash method, and examined under the microscope in order to determine whether the progenitor was a structural homo-
474 DANKO BRNCIC et al.
TABLE 1
Observed and expected number of individuals heterozygous for the gene mrangements in the fourth chromosome, among the flies emerging between 29-35 days after egg laying
(fast), between 37-40 days (medium) and after 44 days (slow)
Experiment 1
Fast Medium
Slow
X 2
P (df 2)
Experiment 2 Fast Medium Slow
X 2
P (df 2)
Chromosome 4R heterokaryotypes Chromosome 4L heterokaryotypes Number Observed Expected Observed Expected
163 110 89.7
79 38 43.5
125 54 68.8
18.80 <0.0001
232 167 131.2
250 131 141.4
250 116 141.4
34.7 <0.0001
110 89.3
38 43.3
53 68.4
19.78
<
0.000 1156 125.6
131 135.2
109 135.2
27.5 <0.0001
The x L homogeneity test summarized in Table 1 shows that there were signifi- cant differences between the chromosomal constitutions of the flies that emerged first and those that hatched later. The number of heterokaryotypes for the gene arrangements in the fourth chromosome was much higher in the former than in those which took longer to develop. As the frequencies of homo- and hetero- karyotypes were similar in both sexes, the Table includes the data of both males and females jointly. It is of interest to note that the results of the two experiments performed with an interval of three months were strikingly similar.
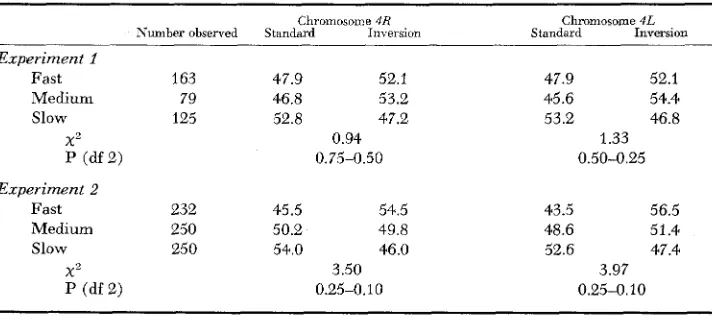
TABLE 2
Frequencies ( i n percent) of Standard and Inversion gene arrangements in the fourth chromosome among flies which emerge first (fast), during the period of maximum
emergence (medium) and which emerge last (slow)
Chromosome 4R Chmmosome 4L
Number observed Standard Inversion Standard Inversion
Experiment I
Fast 163 47.9 52.1 47.9 52.1
Medium 79 46.8 53.2 45.6 54.4
Slow 125 52.8 47.2 53.2 46.8
X 2 0.94 1.33
P (df 2) 0.75-0.50 0.50-0.25
Experiment 2
Fast 232 45.5 54.5
Medium 250 50.2 49.8
Slow 250 54.0 46.0
X 2 3.50
P (df 2) 0.25-0.10
43.5 56.5
48.6 51.4
52.6 47.4
TABLE 3
Observed and expected number of indiuiduals that are heterozygous for the gene arrangements in the fourth chromosome among larvae which are about to pupate first, and those
which are the last to do so ( 5 to 7 days later)
Chromosome 4R heterokaryotypes Chromosome 4L heterokaryotypes Number Observed Expected Observed Expected
First 271 152 145.5 119 125.5
Last 21 7 110 116.5 107 100.5
X* 1.41 1.32
P (df 1) 0.50-0.25 0.75-0.50
Table 2 shows that the frequencies of the “Standard” and “Inversion” gene arrangements in the three groups of adults, were not significantly different. Nevertheless there existed a tendency for the “Standard” arrangement to increase in the slowly developing individuals.
An analysis of the Hardy-Weinberg equilibrium, not reported in any Table, indicated that the whole sample was in agreement with the expectations ( x 2 homo- geneity tests gave values of 1.03 for the gene orders in the right arm, and of 0.92 for those in the left one), although the fast group showed an increase of the hetero- karyotypes
( x 2
value of 27.54 for arrangements in the left and right arm).A group of vials containing eggs, collected as mentioned above, was set apart in order to compare the chromosomal constitution of larvae which reach the pupal stage with di€ferent rates. It is interesting to note that in contrast to the total developmental time taken by the pre-adults (Figure I ) , larval develop- mental time showed a dispersion of only 5 to 7 days. The chromosomes of 271 larvae which were the first ones ready to pupate were examined and the frequen- cies of the different gene arrangements obtained were compared with those of the last 21 7 larvae to pupate within the same vials.
In contrast to the results reported in Table 1 which show highly significant differences in the number of heterokaryotypes in fast and slowly developing flies, it is noteworthy to point to the findings in larvae of different rates of growth, summarized in Table 3. These show that the number of heterokaryotypes was not significantly different in the larvae which pupated first and those which did so 5 to 7 days later.
DISCUSSION
Since the pioneer work of WRIGHT and DOBZHANSKY (1946), it is known that chromosomal polymorphism in Drosophila can be maintained by the superior adaptive fitness of the structural heterozygotes. Following the classic experiments in D. pseudoobscura, heterotic properties of the inversion heterozygotes have been reported in many species of the genus, such as D. melanogaster,
D.
persinilis,D. robusta, D. willistoni, D. subobscurfl,
D.
ananassue, D. pauani,D.
flawpilosa476 DANKO BRNCIC et aZ.
work done on this subject may be found in the reviews of DOBZHANSKY (1961), SPIESS (1962) and SPERLICH (1967). Most of the demonstrations of the superi- ority of the structural heterozygotes in Drosophila come from experiments per- formed in population cages, or comparing the F, hybrids between homozygous stocks with their parents. Although most of the latter type of experiments are designed to avoid confusion between inversion heterokaryotype effects and the effect of heterozygosity for other genes not related to the inversions ( SPIESS 1962), this is not always easy to accomplish. I n the preient study, the method used tends to overcome this difficulty. It allows the comparison of the rate of development of homozygotes and heterozygotes for the inverted zones of the chromosomes within the same population, cultured in the same bottles, therefore reducing to a mini- mum differences in environment or genetic background.
In D. melunogaster, BONNIER (1961) demonstrated that the rate of develop- ment results from the compound effect of several causes, both genetic and environ- mental. Among the latter, crowding, food, and other conditions that affect com- peting ability of the larvae may increase or decrease their developmental rate. The circumstance that in the experiments in
D.
puuani here analyzed, the number of eggs per vial, the amount of food, and temperature were not variables, sug- gests that the differences in the rate of development may be determined by the homozygous or heterozygous condition of the genetic contents of the inverted zones of the chromosomes, and not by other factors.The fact that there are no great variations in the chromosomal structure of larvae which pupate first compared to those which pupate last (Table 3), seems to indicate that the differences in developmental speed between homo- and hetero- karyotypes express themselves mainly during the pupal stage. This result seems of interest, as the environmental agents related to the amount of food, to crowding or other factors affecting competition, which may affect the rate oE development, act almost exclusively during the larval stage. On the other hand, it can be imagined that differences in the rate of development found during the pupal period, may depend more on the individual genotypes or on experimentally con- trollable environmental factors such as temperature. To test the validity of this hypothesis, further experiments will have to be designed in which different developniental stages of the polymorphic strain of D. pavani shall be subject to several competitive and environmental conditions.
I n the introduction it was suggested that the "rigid" character of the chromo- somal polymorphism found in
D.
pauani may be the expression of the strong heterotic properties oE the heterokaryotypes in most of the environmental con- ditions in which the species lives. Utilizing the same method here employed,BRNCIC and DEL SOLAR (1 961 )
,
showed that heterokaryotypes survive longer; and BRNCIC and KOREF-SANTIBA~~EZ ( 1964) and KOREF-SANTIBAGEZ and BRNCICpauani
It is difficult to evaluate the extent to which the faster rate of development of the he terokaryotypes will contribute to maintain their high frequency in natural populations or laboratory stocks, as other components of fitness, such as longevity and mating activity, are also enhanced in the heterozygotes. Although the experi- ments here reported were not designed to compare the relative importance of
these factors in the reproductive efficiency of the heterolraryotypes, some con- siderations may be advanced. The individuals that develop earlier probably mate first and thus contribute more offspring to the population. Moreover, the hetero- karyotypes will occupy the available ecological niches before the corresponding homokaryotypes. I n this way the advantage in rate of development may represent one of the critical factors in maintaining the chromosomal polymorphism ex-
hibited by D. pauani.
The authors wish to acknowledge the valuable technical aid of MRS. HERTHA FENNER DE GPEZ,
MISS PATRICIA ITURRA and MR. ENRIQUE Z ~ A T E . They also wish to thank PROFESSOR GEORGE HODGSON for reading and criticizing the manuscript.
SUYMARY
The present work is part of a series dedicated to study the physiological proper- ties of heterokaryotypes in the neotropical species,
D.
pauani, that exhibits “sta- ble” polymorphism for gene arrangements due to the presence of inversion. Ithad previously been found that the heterozygotes show greater longevity and have a superior mating activity. In the experiments here described, rate of development was analyzed. Eggs collected from a heterogenous stock of the species were placed in equal numbers in vials at 16”C, and all the imagoes which emerged were recorded, and divided into three groups: those of fast development which took 29 to 35 days to hatch, those which emerged between 37 to 40 days, and the slow ones, between 4 4 and 65 days. There was a significantly higher number of hetero- karyotypes for the gene orders in the fourth chromosome among the flies that developed rapidly compared to those which took longer. On the other hand, no significant differences were found between the frequency of heterozygotes for inversions in larvae which reached the pupal stage first and those which did so last,
5
to 7 days later.L I T E R A T U R E CITED
BONNIER, G., 1960
BRNCIC, D., 1957
Experiments on hybrid superiority in Drosophila melanogaster. 11. Rate of development from egg hatching to eclosion. Genetics 46: 86-91.
Chromosomal polymorphism i n natural populations of Drosophila pavani. Chromosoma %: 699-708.
-
1961 Nonrandom association of inversions i n Drosophila pauani. Genetics 46: 401406.Life cycle and the expression of heterosis in inversion heterozygotes in Drosophila funebris and Drosophila pauani. Am. Naturalist 95 : 21 1-216.
Mating activity of homo- and heterokaryotypes in Drosophila pauani. Genetics 49 : 585-591.
The genetic characteristics of marginal populations of Drosophila. Cold Spring Harbor Symp. Quant. Biol. 2 0 : 276-287. __ 1965 Chromosomal morphisms in geo- BRNCIC, D., and E. DEL SOLAR, 1961
BRNCIC, D., and S. KOREF-SANTIBAAEZ, 1964
478 DANKO BRNCIC et al.
graphically widespread species of Drosophila. In: T h e Genetics of Colonizing Species. pp. 503- 531. Academic Press, New York.
On the dynamics of chromosomal polymorphism in Drosophila. Symp. Rev. Entomol. Soc. London 1: 3042. __ 1962 Rigid us. flexible chromomnal poly- morphism in Drosophila. Am. Natwralist 96: 321-328.
KOREF-SANTIBAWEZ, S., 1963 Courtship and sexual isolation i n five species of the mesophrag- matica group of the genus Drosophila. Evolution 17: 99-106.
KOREF-SANTIBAWEZ, S., and D. BRNCIC, 1965 Mating activity and chromosomal polymorphism i n Drosophila pauani females. Genetics 52 : 453.
SPERLICH, D., 1967 Populationsgenetik (Teil. I. Drosophila). Fortschritte der Zoologie 18: 223-278.
SPIESS, E. B. 1962 Papers on Animal Population Genetics. (Introduction: xi-xii). Little, Brown, Boston, Mass.
WRIGHT, S., and TH. DOBZHANSKY, 1946 Genetics of natural populations. XII. Experimental reproduction of some of the changes caused by natural selection in certain populations of