Bertina Rudman, Louise B. Preer, Barry Polisky and John R. Preer, Jr.
Program in Molecular, Cellular and Developmental Biology, Department of Biology, Indiana University, Bloomington, Indiana
47405
Manuscript received March 18, 199 1 Accepted for publication May 17, 1991
ABSTRACT
In Paramecium tetraurelia, stock 51, the A surface protein is coded by the wild type AI’ gene, present in micronuclei in two copies and in macronuclei in about 1500 copies. DNA processing, comprised of DNA cleavage, copy number amplification and telomere addition occurs at autogamy and conjugation when old macronuclei degrade and new macronuclei are formed from micronuclei. In this paper we characterize mutants with macronuclear A gene deletions. These mutants are notable in three respects. First, the mutants do not appear to be simple micronuclear deletions. Although genetic analysis shows that the d l 2 mutant d l 2(-1300) is homozygous for the allele A-”” and the mutant d 12(+ 1) for A+’, analysis by the polymerase chain reaction indicates that the micronucIei in these two mutants contain intact, but presumably altered, micronuclear A genes. They undergo deletion during DNA processing when new macronuclei are formed. Second, the position of the deletions in these alleles has been shown to change. T h e deficiency present in the d l 2 allele A”3w was originally determined to extend from position
-
1300 (relative to the start of translation of the A gene) to the end of the chromosome. Later, a derivative of this strain, homozygous for the d l 2 allele A+’ was isolated in which the start site of the deletion was found to have moved from-
1300 to+
1. Third, a surprising interaction occurs in crosses between a line homozygous for the d l 2 allele and one homozygous for the wild-type A’’ allele. Previous work on the non-Mendelian d48 mutant (which has intact A’’ genes in its micronucleus, but has truncated A’’ genes in its macronucleus) has shown that intact A” alleles must be present in the old macronucleus in order for As’ alleles to undergo proper processing. We find that d l 2 alleles act on A5’ alleles in heterozygotes such that intact macronuclear A genes are no longer required for proper processing of A”. Thus, in crosses of 51 X d l 2 (either +1 or -1300) d l 2 exconjugants, as well as 51 exconjugants, give rise to clones carrying both intact A’’ and truncated d l 2 alleles. Remarkably the d l 2 alleles, which are themselves deleted during processing, are capable in the heterozygote of fostering normal processing of the As’ allele.T
HE A gene that determines surface protein A in wild-type strain 51 of Paramecium tetraurelia is found near the end of a macronuclear chromosome oriented with the 3‘ end of the gene nearest a telom- ere (FORNEY et al. 1983). Two copies of the gene are present in the diploid micronucleus and about 1500 copies in the macronucleus. In the non-Mendelian mutant, d48, the intact A gene is present in the micronucleus, but was reported to be absent from the macronucleus (EPSTEIN and FORNEY 1984). EPSTEIN and FORNEY (1984) showed that the macronuclear deletion in d48 starts in the vicinity of+
1, measured from the start of translation. Subsequently, in four separate cloned genes, DNA sequencing revealed that the deletions started at -79, +33, +121 and +149 (FORNEY and BLACKBURN 1988) with a telomere at- tached at the end of each cloned gene. Since the A gene is missing, serotype A cannot be expressed under conditions known to produce A and the strain is designated A-. EPSTEIN and FORNEY (1984) reported that when d48 (A-) is crossed to 51 wild type (A+) the d48 exconjugant usually produces an A- F1 clone andGenetics 1 4 9 47-56 (September. 1991)
the 5 1 wild-type exconjugant produces an A+ F1 clone. No further change in type occurs after autogamy in the F2 generation. Although half of the F2 clones derived from the 5 1 wild-type exconjugant are homo- zygous for the A allele that came from the d48 parent, all of the F2 clones of the 51 exconjugant are wild- type A+. These results demonstrate that the micro- nuclei of d48 must contain the complete wild-type A gene, A5’.
However, the non-Mendelian pattern of inheritance shown by d48 does not indicate true cytoplasmic in- heritance but appears instead to be due to the influ- ence of the old macronucleus on the newly forming macronucleus during conjugation and autogamy. This conclusion is reached on the basis of a series of trans- fers of macronucleoplasm and cytoplasm by HARU-
MOTO (1 986) and KOIZUMI and KOBAYASHI (1 989).
gene into the macronucleus of d48 would cause per- manent reversion of d48 to wild type. KOIZUMI and KOBAYASHI (1 989) showed that injection of the com- plete A gene into the macronucleus of d48 results at the next autogamy in the permanent restoration of the ability of the cells to produce A. More recently, YOU et al. (1991) and H. JESSOP-MURRAY and L.
MARTIN (personal communication) have shown that injection of specific subfragments of the A gene into d48 results in permanent rescue. Thus, old macro- nuclei bearing at least a portion of the A gene must be present for proper DNA processing of new A genes during development of new macronuclei. Since d48 is deficient in complete macronuclear A genes, proper processing of new A genes at conjugation and auto- gamy does not occur. Possibly, d48 arose as a conse- quence of a developmental abnormality in DNA proc- essing. Once lost from the macronucleus, A genes could not be restored at subsequent autogamies be- cause the A gene, necessary for correct processing, would be missing. This dependence of new macro- nuclear A genes on the presence of old macronuclear A genes constitutes an anomalous genetic mechanism which appears to account for the facts acquired thus far. Its molecular mechanism is completely unknown. In addition to the non-Mendelian mutant, d48, which requires the presence of A genes in the old macronucleus for proper A gene processing, mutants
d 1 and d 12 were isolated by L. EPSTEIN and J. FORNEY (unpublished results). These mutants also lack com- plete A genes in their macronuclei, and preliminary results suggested that these mutants were inherited in a Mendelian fashion.
In this paper we have characterized the d l and d l 2 mutants, and show that there are actually two d l 2 strains that have mutant alleles at the A locus. T h e d l 2 strains exhibit both Mendelian and non-Mende- lian inheritance. We show that in heterozygotes car- rying either of the d l 2 alleles and the wild-type allele, processing of the wild type allele is usually normal. Normal processing of the wild-type allele occurs not only in cells whose macronuclei have normal A genes, but also in cells whose macronuclei have defective A genes. Finally, we present evidence that the d l 2 mu- tant micronuclear A gene copies are not truncated, but that they affect DNA processing by causing dele- tions during the formation of new macronuclei during autogamy and conjugation.
MATERIALS AND METHODS
Strains: Stock 51 (ATCC #30303) is homozygous for the wild-type A'' gene. Strain d48 was derived from 51 as described earlier by EPSTEIN and FORNEY (1 984) using X- ray mutagenesis and antiserum selection. Strains d12, d l and d l 6 were obtained in a similar fashion. Several ami- cronucleate lines, and also chimeric strains produced by transplanting wild type micronuclei from stock 51 into ami- cronucleate d12, were provided by S. KOIZUMI and S. KO-
BAYASHI. They removed micronuclei from a number of d l 2 cells, designating them K1, K2, etc. They then transplanted wild-type micronuclei into these cells, and used each to start new lines, Kl-I, K1-2, K2-1, etc. d l , d l 2 or d48 cannot produce serotype A surface antigen when cultured at 34" for several fissions. By contrast, wild-type stock 5 1, cultured at 34", generally switches to serotype A within 3 or 4
fissions. The original d l 2 was later redesignated d 12(- 1 300) to denote its deletion start relative to the begin- ning of translation of the A gene. A variant of d l 2(-1300) with an altered deletion appeared spontaneously in our culture collection and was designated d l 2(+ 1). These strains are characterized in more detail in RESULTS. d48 has been known to revert to wild type sporadically with low fre- quency, and d l 2 has been suspected of reverting two or three times over the course of several years. The strain d4- 6 (ATCC #30987) is isogenic with stock 5 1 but homozygous for several marker genes including the A'' allele (SONNE- BORN 1975).
Culture of paramecia: Paramecia were cultured in an infusion of 1.5 g/liter of wheat Cerophyl (Pine Brothers, Kansas City, Missouri) supplemented with 0.1 g of Bacto- Yeast Extract, 1 mg of stigmasterol, and 0.4 g Na2HPO4. It was inoculated with Klebsiella pneumoniae a day or two before use.
Crosses: Matings, induction of autogamy and serotype testing were carried out as described by SONNEBORN (1975). A cross of two lines in Paramecium yields F1 exconjugant clones with identical genotypes. An FP obtained by inducing autogamy in such FI clones yields a series of homozygous F2 clones. For each genetic locus, 1/2 the F2 clones are homo- zygous for the allele found in one parent and 1/2 are homozygous for the allele found in the other parent. In the crosses described here at least one Mendelian marker was present, and lines were eliminated from further considera- tion if these markers did not indicate proper exchange of nuclei and normal genetic behavior. The Mendelian mark- ers used were twisty, paranoic A, and a trichocyst nondis- charge mutant (SONNEBORN 1975). Mating type was used as a cytoplasmic marker. Moreover, the paranoic marker also usually made it possible to distinguish the two parents, for the original phenotypes were almost always still present at the time that exconjugants were separated. Tests for the ability to produce serotype A were done by allowing cells to undergo several fissions at 34 *, then using antiserum to test for serotype A. The most reliable method was that employed by EPSTEIN and FORNEY (1984), adding fresh medium to double the volume of cultures in test tubes daily for four days at 34" In most cases wild-type cultures were found to be more than 50% A, usually nearly 100% A and were designated A+. Those producing no A were designated A-. Only in very rare instances was a culture that failed to show serotype A by this test found to have the A gene by molecular tests. In such cases a second growth period of four days at 34" always revealed serotype A antigen expression. EPSTEIN and FORNEY (1984) reported that in the progeny of the cross d48 X 51, occasional exconjugant clones were pro- duced which yielded both A+ and A- subclones. We have confirmed their results and refer to such clones as "mixed" clones. They also occur in progeny of the cross d48 X d12.
Preparation of DNA from Paramecium: Up to 500,000
Affecting DNA Processing 49
once with 75% ethanol and desiccated. Occasionally a sec-
ond phenol treatment was necessary in order to obtain
efficient cutting with restriction enzymes.
Hybridization: Southern blots and hybridizations were
carried out as previously described (GODISKA et al. 1987). Plasmids: The plasmid pSAl4SB contains the region -1590 to +8229 of the A gene in a pT7/T3-18 vector (GODISKA et al. 1987). The plasmid pSA3.75Hd spans the region -3300 to +456 of the A gene and is in a pUC8 vector. It was prepared by J. D. FORNEY.
Polymerase chain reaction (PCR): The reaction mixes
were prepared by adding: 3 PI of purified DNA solution of
the desired concentration, 5 pl of 1% NP-40, 1 pl Tetrahy-
mena DNA (0.1 pg/pl) as carrier, 10 pl of 10 X buffer (500 mM KCI, 100 mM Tris-CI, pH 8.3, 15 mM MgCln, 0.1% gelatin (wt/vol)), 16 pl of NTPs (12.5 mM each of dCTP, dGTP, dATP and dTTP), 5 pl of each primer (OD260 =
5.0), 3 pl of Taq polymerase (total of 2 units) and Hz0 to
100 pl. When using whole cells rather than isolated DNA,
the following procedure was used. A population count on
the culture to be used was made by spreading 20-60 pl of
the culture in a long narrow streak onto a glass microscope slide and counting the number of cells using low magnifi- cation. The volume of the culture needed to give the re-
quired number of cells was computed, and this volume was
added to the 0.5-ml reaction tube. Then 1.7 times the
volume of 1% NP-40 was added and the mixture quickly
spun for 10 sec at top speed in a microfuge. T h e supernatant was removed with suction to leave a volume of 8 pl. Rapid processing is necessary to avoid premature lysis of the cells.
After 10 min at 65" the 8-p1 aliquot was used instead of 3
pl of DNA and 5 pl of 1% NP-40 in the reaction mix
described above. The amplifications were made in a Perkin
Elmer PCR cycler as follows. A preliminary incubation at 94" for 15 sec then 60 cycles of 92" for 1 min, 50" for 2 min, 72" for 2 min with a 3-sec extension on the 72" segment, followed by a final 7 min at 72".
RESULTS
A gene deletions begin at different sites in differ- ent mutants: Before investigating the genetics of the mutants other than d48 we needed to characterize the position of their deletions. L. N. EPSTEIN and J. D.
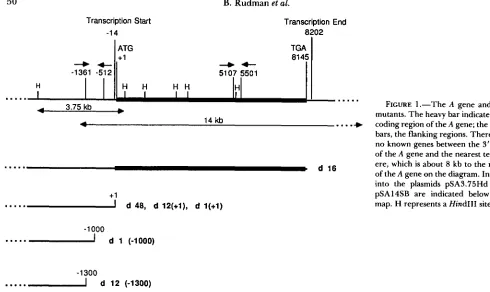
FORNEY (unpublished results) isolated the A- Mende- lian mutant, d12, which they found, using hybridiza- tion to Southern blots, was deleted starting at approx- imately position -1300. In our laboratory a variant of this strain arose which proved to be inherited in the same way as the original d l
2,
but which we found to be deleted starting farther downstream at approx- imately position+
1. T h e point of deletion appears to be very close or identical to that in d48. These two d l 2 strains are designated d12(-1300) and d12(+1). See Figure 1 . T h e two strains can be distinguished from each other and from wild-type strain 51 byhybridization to Southern blots of restriction enzyme- digested genomic DNA. See Figure 2. It is also possi- ble to distinguish them by PCR using primers span- ning the region from
-
136 1 to -5 12 (Figure 1) which give an 850-bp product with 100 cells of d12(+1) and no product with d l 2(-1300). Experiments utilizing this technique will be described later and are shown in Figure 4.In addition, a Mendelian A- mutant, d l , was iso- lated by L. N. EPSTEIN and
J.
D. FORNEY (personal communication), which they found was deleted start- ing at approximately-
1000. We find, using Southern blots, that the deletion point in all the current lines of d l has shifted and is now found to start at approxi- mately +1 (data not shown).Mendelian inheritance in the crosses of d l to 51 wild type: Crosses of d l 2 to wild type yield the results expected if d l 2 differed from wild type by a single recessive gene mutation. We find essentially the same results for both d12(-1300) and d12(+1). Most of the F1 exconjugant clones were A+, i.e., capable of ex- pressing A, before they went into autogamy, even those derived from the d l 2 exconjugant. Although a few of the F1 lines did not express A, it is thought that the number of fissions before autogamy is insufficient to allow adequate testing of the ability of all cultures to go to A at high temperature. In a cross of d12(-1300) to 51 wild type the genotype of the F1 was examined by hybridization to blots. T h e DNA from four of these FI lines (the exconjugants from two pairs, both of which were A+) was isolated and cut with HindIII, electrophoresed, blotted and probed with labeled pSA3.75Hd DNA. This plasmid contains the sequences upstream of the A gene from -3300 to +456. four showed two bands, the larger one indicat- ing the presence of the 51 wild-type allele, and the smaller one indicating the presence of the -1 300 deletion. Three of the F1 lines are shown in Figure 3, lanes 1-3.
An F2 was obtained by establishing a series of lines from autogamous F1 isolations. T h e resulting F2 lines were then tested for their ability to produce A after four or more fissions at 34". The ratio of A+ to A- lines was determined for each exconjugant clone that went through autogamy. T h e results are shown in Table
Z
and summarized in Figure7.
T h e FP pattern is clearly Mendelian, very close to the expected single factor ratio of 1 A+: 1 A- both for the individual ratios and for the overall ratio of 149: 154 summed for the crosses d12(+1) X 51 and d12(-1300) X 51. PCR using primers spanning the region from -1 361 to -512 (Figure 1) was used to determine the deletion points in the FZ lines. In the cross of d12(+1) X 51, 20 of the A- FP clones all showed the +1 deletion as expected. In the cross of d12(-1300) x 51, 12 ran- domly selected A- F2 clones all showed the -1 300 deletion. See lanes 1-6, Figure 4, which show some of the lines with the -1 300 deletion.Transcription Start
-1 4
B. Rudman et al.
Transcription End 8202
+4-
-1361 -512 5107 5501
...
4 3.75 kb
...
b
4 14 kb
.
.
. ..)
...
d 16.d 48, d 12(+1), d 1(+1) .-.*I
+1
-1 000
-.-..I
d 1 (-1000)-1300
...
d 12 (-1300)macronuclei of A+ cells. On the basis of the latter hypothesis one would predict that the lack of the normal number of A genes in the macronucleus of d l 2 would have resulted in the production of all A- progeny from the d l 2 exconjugant clones. However, in the heterozygous genotype, consisting of one d l 2 allele and one 51 wild-type A allele, the wild type A gene is processed normally into macronuclear A genes at conjugation, even in the absence of complete A genes in the old macronucleus. We will return to a consideration of this effect later.
Deletions in Mendelian mutants are determined by alleles at the A locus: A cross of d12(-1300) A- X d l
2(+
1) A- gave all A- F1 exconjugant clones. TheDNA from six of these F1 lines was isolated and the DNA was cut with HindIII, electrophoresed, blotted and probed with pSA3.75Hd. All six preparations showed two bands, indicating approximately equal amounts of the two parental macronuclear chromo- somes. Three of the F1 lines are shown in lanes 4-6 of Figure 3.
Fz lines were obtained by inducing autogamy in the F, lines. Again all the lines were A-. Eighteen of the
Fz lines chosen at random were screened as described above with blots and hybridization, in order to see how much of the A chromosome each contained (data not shown). It was found that nine of these lines were deleted at -1 300 and nine were deleted at +1, pre- cisely the 1:l ratio expected on the basis of a single Mendelian factor difference. PCR analysis, using the primers spanning the region
-
136 1 to -5 12 as de-scribed above, confirmed these results. See Figure 4, lanes 7-10. Thus, the position of the deletion in the
FIGURE 1.-The A gene and the mutants. The heavy bar indicates the coding region of the A gene; the light bars, the flanking regions. There are no known genes between the 3' end
of the A gene and the nearest telom- ere, which is about 8 kb to the right
of the A gene on the diagram. Inserts into the plasmids pSA3.75Hd and pSA14SB are indicated below the map. H represents a Hind111 site.
two strains is determined by a pair of simple Mende- lian alleles.
The question of whether the d l 2 mutations are allelic with the A locus that determines the amino acid sequence of the A protein was addressed by further crosses. d l 2 was derived from stock 5 1 which is homo- zygous for the allele A5'. The strain d4-6 is isogenic with stock 51 but homozygous for several marker genes including the AZ9 allele
Fc
'NEBORN 1975).Serologically distinguishable iml:oi.ilization proteins determined by different alleles have been shown to differ in their primary amino acid sequence (reviewed in SONNEBORN 1975). We have utilized appropriate antisera that can distinguish the two antigenic types. If the two d l 2 strains are mutant at the A locus, a cross of d l 2 to a strain bearing AZ9 should give in the Fz A+ and A- clones in a 1: 1 ratio and all the A+ would be serotype 29A. If the two are not allelic, then the gene for serotype 5 1 A should be present in d 12, since d 12 was derived from stock 5 1. Therefore, 5 1 A recombinants, as well as 29A should appear in the
Fz.
Mutants Affecting DNA Processing 5 1
FIGURE 2.-DNA blot analysis of the d l 2 deletions. DNAs ( 5
pg/lane) were cut with Hind111 and electrophoresed on agarose. Lanes I and 4. d 12(- 1300). Lanes 2 and 5 , d 12(+ 1). Lanes 3 and
6. stock 51. The gel was blotted and probed with pSA3.75Hd (spanning the region -3300 to +456) and shown in lanes 1, 2 and
3 . The blot was then washed and reprobed with pSA 14SB (spanning the region -1590 to +8229) and shown in lanes 4, 5 and 6. The 3.75Hd probe reveals the short remaining portion of d12(-1300) in lane 1 and the longer piece of d l 2(+1) in lane 2. The latter terminates near a Hind111 site and hence appears fuzzy because of variation in the precise cut site and variation in the length of telomeres. The pSA14SB probe reveals all the internal fragments of the A gene in stock 5 1 which are missing in the two mutant deletion strains as well as a few weaker cross-reacting bands derived from regions of the genome other than the A gene. The differences i n the cross-hybridizing bands (approximately 8 kb) in the two d l 2 mutants are not specific for the mutants.
one of these lines judged to be A- on a routine test was examined by PCR, the results indicated that the complete A gene was present. Subsequent serotype testing revealed that it was indeed able to produce 29A.
Thus, there is no evidence for the presence of additional independent loci that affect the deletions in any way. T h e only interpretation that we find compatible with these results is that d12(-1300) is homozygous for the allele A-””” and that d 12(+ 1) is homozygous for the allele A+’.
Mendelian non-Mendelian inheritance in crosses
of d l 2 and d48: We find (see Table 1 and summary in Figure
7)
that the crosses of d48 X d 12 (both-
1 300 and +1) often show a simple Mendelian pattern of inheritance in the F2 (1 : 1 ratio of A+:A-). However, we also see that in many cases the ratios are strongly biased in favor of the A- class. These biased ratios are derived not only from the d48 exconjugant but also from the d l 2 exconjugant. Thus, the non-Mendelian effect arising from a deficiency of intact macronuclearA genes is characteristic not only of d48, but also of
FIGURE 3.-DNA blot analysis of heterozygotes. DNA (2 pg/ lane) was extracted from F, cultures, cut with Hindlll, and electro- phoresed on agarose. The gel was blotted and probed with pSA3.75Hd. The DNA in lanes 1-3 was taken from different exconjugant clones of the cross d l 2(-1300) X 5 1 . The DNA in lanes 4-6 was taken from the cross d12(-1300) X d12(+1). The band of about 2.0 kb represents the sequences present in the d l 2(-1300) chromosome. The band at approximately 3.8 is derived from wild-type chromosomes in lanes 1-3. and from the d12(+1) chromosomes in lanes 4-6.
d 12. KOBAYASHI and KOIZUMI (1 990) reported results similar to these, except that they occasionally found FI exconjugants that failed to segregate in the
FP,
all progeny remaining A+. They also reported such lines from the cross d l 2x
51. We have never observed such clones. Perhaps such lines represent reversion of d l 2 to wild type. I t is not known in their crosses whether they were working with d12(+1) or d 12(- 1300), since the crosses predated the discoveryof the two types of d 12.
Although in the data reported in Table 1, deviations from a 1 : 1 segregation appeared more frequently in the cross d48 X d 12(- 1300) than in the cross d48 X d12(+1) we cannot be sure that d12(+1) and
d 12(- 1300) really differ in this respect, for consider- able variation was found in the results from crosses made at different times. In addition, it appears that the d48 exconjugant is more likely to yield aberrant results than the d l 2 exconjugant, although this con- clusion is also not certain.
TABLE 1
F n ratios from crosses involving 51, d48 and d l 2
Progeny of 5 I Progeny of d 12
A':A- /\+:A-
exconjugant exconjugant Cross
5 1 A * X d12(+1)A- 21:16 24:24
28:20 18:18
Sun1 = 9 I :78
51A' X d12(-1300)A- 8:4 6:6
5:7 6:6
5:7 4:8
2:10 7:5
6% 7:13
Sum = 58.76
Sum for both crosses = 149: 154
Progeny of d4A Progeny of d I2 exconjugmt
A+:A-
exconjupnt
A':A-
d48A- X d 12(+ 1)A- 11:11 7:7
11:13 10:14
19:13 12:13
13:l 1 2 5 2 2
15:16 21:17
19:22 18:17
0:48 0:15
21:15
d48A- X d12(-1300) 5:19 14:10
2:20 13:10
0:23 10:14
6:17 3:2 1
6:18 0:23
Autogamous cells from the indicated crosses were isolated and later tested for their ability to produce serotype A. The ratios of A
producers (A') to nonproducers (A-) are given in the table. The cytoplasmic parent of each group of F2 progeny is indicated for
each exconjugant.
run on these A- F2 cultures one would expect that the half of these clones of genotype A-'300/A"300 would have the
-
1300 deletion. The other half of the clones of genotype A5'/A5' that are derived from the d48 exconjugant in which the old macronucleus contains a +1 deficiency, would be expected to have the +1 deficiency. These predictions were confirmed by PCR; four F2 clones derived from a d48 exconjugant were found to be +1 and four were -1 300. It is more difficult to predict what might be the nature of the deletion in the A- A5'/As' segregants derived from the d12(-1300) exconjugant. In this case the oldmacronucleus that influences the nature of the newly forming macronuclei has a -1300 deletion and not a +1 deletion. Similar PCR tests were run on F2 segre- gants derived from a single d12(-1300) exconjugant, all of which were A-. Of 23
F2
clones 7 were +1 and 16 were -1 300 deletions. Although the fit to a 1 : 1 ratio is far from perfect in this case, a chi square test shows that the fit is not significantly different from1:l (chi square with Yates correction = 2.78, proba- bility 0.1). Thus, the A- A5'/A5' segregants from the d 12 exconjugant had the
+
1 deletion and the position of the deletion was not affected by the -1 300 alleleA- F2 F2
-;;io
::n
r-~
d12(-13M))x51 d12(-1300)xd12(+1)
1 2 3 4 5 6 7 8 9 1 0 M
1 .o
0.5
FIGURE 4.-I'CR analysis of I;? segregation. A pair of primers spanning the region from
-
136 1 to -5 12 was used with 100 cells in PCR in order to distinguish d12(+1) molecules which give an 850-bp PCR product from d l 2(-1300) molecules which give no product. A portion of each reaction product was run on an agarose gel and stained with ethidium bromide. Lane 1, d l 2(-1300). Lane2, d l 2(+1). Lanes 3-6, four of the A- FY segregant clones obtained by autogamy from a cross of d 12(- 1300) X 5 1 show no band and hence are d 12(- 1300). Lanes 7-1 0 , four randomly chosen F2 clones from the cross d 12(- 1300) X d 12(+ 1) (all clones from the cross were A-.) Lanes 8 and 10 show no band and hence are d 12(- 1300):
lanes 7 and 9 show the product characteristic of d12(+1): M. markers. A PCR product of the predicted size, 850 bp. is found only in d 12(+1) and not in d 12(- 1 300).
TABLE 2
Tests for allelism: Fn ratios from exconjugants of d4-6 X d l 2 crosses
Progeny of d4-6 Progeny of d l 2 exconjugant exconjugant
Cross A':A- A+:A-
d4-6A' X d I2(+ 1)A- 11:16 15:21
14:10 10:13
9:11
Sum = 5 9 7 1 d4-6A+ X d 12(-1300 1)A- 14:27 7:17
8:12 14:U
10:12 1 4 9
9:15
Sum = 76: 100
Sum for both crosses = 135: 17 1
Autogamous cells from the indicated crosses were isolated and later tested for their ability to produce serotype A. d4-6 homozy- gous for the allele A'". The ratios of A producers (A') to non- producers (A-) are given in the table. The cytoplasmic parent of each group of F2 progeny is indicated for each exconjugant. All
lines were 29A.
Mutants Affecting DNA Processing
TABLE 3
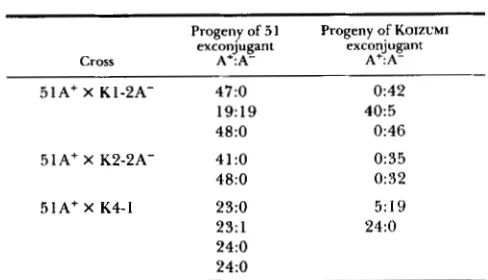
Fn ratios from crosses of KOIZUMI’S micronuclear transplant strains
Progeny of 5 1 Progeny of KOIZUMI Cross
exconjugant
A+:A-
exconjugant
A+:A-
51A+ X K1-2A- 47:O 0:42
19:19 40:5
48:O 0:46
5 1A+ X K2-2A- 41:O 0:35
48:O 0:32
51Af X K4-1 23:O 5:19
23: 1 24:O
24:O 24:O
Autogamous cells from the indicated crosses were isolated and later tested for their ability to produce serotype A. The ratios of A
producers (A+) to nonproducers (A-) are given in the table. The cytoplasmic parent of each group of F2 progeny is indicated for each exconjugant.
even when the old macronucleus is derived from d l 2 with a deficiency starting at - 1300.
Finally, an apparent effect of micronucleoplasm on the non-Mendelian determination is seen in the d l 2 X d48 crosses. In these crosses the d l 2 exconjugants often produce A- FI clones and give an excess of A- at the next autogamy. However in the cross d l 2 x 5 1 the heterozygous d 12 exconjugant always changes to
A+ in the F1 and gives a 1 : 1 ratio of A+:A- in the F2
by autogamy. Thus a wild-type A” allele received from a 5 1 wild-type cell is less likely to suffer deletions than a wild-type A5’ allele received from a d48 cell.
Creation of pseudod48 lines: If our idea about the nature of d48 is correct, it should be possible to recreate a d48 strain artificially by micronuclear trans- plantation. KOBAYASHI and KOIZUMI (1 990) removed the micronuclei from several individuals of d 12 (start- ing point of the dl2 deletion not known) and replaced them with micronuclei from 51 wild type. Since such cells have macronuclei with few copies of the A gene, and micronuclei with normal A genes, they should be incapable of producing normal macronuclei with in- tact A genes at autogamy. In fact, they should be exactly like d48, even though derived from wild type and d12. They should breed true as A- strains through vegetative divisions and also through auto- gamy. Furthermore, they should behave like d48 in crosses. T h e data given in Table 3 represent the results of crosses between wild-type and the Koizumi strains. They are typical of the results obtained when wild type is crossed to d48 (EPSTEIN and FORNEY 1984), most pairs yielding A+ clones from the wild- type exconjugant and A- from the transplant excon- jugant, both in F1 and F P . T h e occasional A+ clones
arising from the A- transplant exconjugant are also typical of d48. In two cases A- lines were derived from a wild type exconjugant (1 9 A+: 19 A- and 23 A+:l A-. See Table 3). This result was unexpected,
and no follow-up was made in these cases to investigate the nature of the A- lines. Tests using PCR showed that all the chimeric lines contained +1 deletions.
A cross of one of the Koizumi “pseudo d48” strains to d l 2(+1) (data not shown) is consistent with the results obtained by KOBAYASHI and KOIZUMI (1 990) in a similar cross. T h e phenotypes of the progeny were similar to those obtained in the cross of d48 x d12(+1). See Figure
7.
Downstream A gene sequences are present in d l 2
micronuclei and in low copy number in micronuclei: Perhaps the simplest conclusion concerning the two d l 2 deletions is that the micronuclei have sustained deletions which are passed on to the macronuclei. On the other hand, data shown below suggest that the micronuclei of the d 12 strains contain possibly com- plete, but mutant A genes, and that the deletions occur during the formation of new macronuclei at conju- gation and autogamy.
T o probe directly for the presence of the down- stream A gene sequences in d12, PCR reactions were carried out. T w o 20-bp primers were constructed for a region that is about 5 kb downstream from the apparent break points for macronuclear DNA in d48 and d12. T h e primers direct amplification of a 395- bp region starting at position 5107 and ending at 5501. See Figure 1. T h e region also contains an asymmetrically placed Hind111 site at 5277, which is useful in identifying the amplified product. Figure 5 shows the results of PCR on whole cell DNA from 5 1, d48 and d12. All three DNA preparations contain the downstream region. In each case, Hind111 digestion yields two fragments of the expected sizes (222 bp and 173 bp), confirming the identification of the product.
T o obtain a rough estimate of the relative number of the putative full length A genes in d l 2, d48 and 5 1, we carried out PCR reactions with fixed numbers of whole cells as starting material. T h e results are shown in Figure 6. Positions are displaced in some cases because of overloading. T h e results suggest that the number of copies of the downstream region in d48 is considerably less than the number in wild-type strain 51, yet more than the number in d12. If this were the case, it would indicate that d48 contains a small number of A genes in its macronucleus. T o investigate this possibility we obtained clones of ami- cronucleate d48 and d12. PCR carried o u t on the amicronucleate d48 and d l 2 both showed that the macronuclei do indeed contain the sequences, but in low copy number (data not shown). Thus, the defects in DNA processing are not absolute, and a few copies of possibly intact A genes are present in macronuclei of all the mutants.
FIGURE 5.-PCR detection of d~wnstream A gene sequences in the mutants. A pair of primers spanning the region from 5107 to 550 I was used with 1 pg of purified D N A in PCR in order to detect the presence of downstream A gene sequences. PCR products were purified and portions cut with HindIII. The HindIII site in the 395- bp region of the A gene spanned by the primers starts 222 bp from one end of the region and 173 from the other end. Appropriate quantities of the uncleaved and cleaved DNAs (lanes designated U and C, respectively) were run on a 5% acrylamide gel. M, markers. Lanes 1 and 2, d48. Lanes 3 and 4, d12(-1300). Lanes 5 and 6,
51. Lanes 7 and 8, pSA14SB. All the mutants appear to have the downstream sequences. Control uncut DNAs in lane 9 (from the unrelated ciliate, Colpidium) and lane 10 (from Escherichia coli) fail to show the 395-bp PCR product.
in the macronucleus. Truncated A genes make
up
thebulk of the macronuclear copies. DISCUSSION
Nature of the Mendelian mutations: T h e d 12 mi- cronuclear alleles A-""" and A+' always give rise to macronuclear deletions beginning at -1 300 and +1 respectively, even when they are present in heterozy- gous combinations with each other or with wild-type alleles. Thus, in a cross of d l 2(-1300) to wild type, both F1 exconjugants, even though they are A+, each contains -1300 as well as wild-type DNA molecules in the macronucleus. Moreover, deletions associated with the A"300 and A+' alleles are not influenced by the composition of the old macronucleus during for- mation of new macronuclei; they always produce dele- tions. On the basis of the genetic behavior alone, one might conclude that these two alleles represent simple micronuclear deletions.
However, certain observations suggest that the d 12 phenotype is not due to micronuclear deletion. First, in two instances we have found A+ cells in stock cultures of d 12. Barring the unlikely chance of con- tamination in two separate cases, these represent re- versions of d l 2 to wild type. Second, strains with extensive deletions such as d l 2(-1300) and d 1(- 1000) have given rise to strains with less extensive deletions, e.g., d 12(+ 1) and dl(+ 1). Third, PCR analy-
FIGURE 6.-Effectiveness of different numbers of cells as tem- plates for PCR. Cells of d48, 5 I and d 12 were used as templates for primers spanning the region 5 10 1 to 5495. Lanes 1, 2, 3, d48:
10, 60, 600 cells, respectively. Lanes 4, 5, 6, 5 1: 1, 10, 60 cells, respectively. Lanes 7, 8, 9, d12: 10, 60,600 cells, respectively. The results suggest that d48 has fewer copies of A gene than wild type, and that the amount in d 12 is even less.
sis showed that the micronuclei in the d l 2 lines con- tain downstream regions of the A genes as well as the upstream flanking portion. PCR analysis on amicron- ucleate d l 2 lines indicated that a small number of possibly intact A genes are present in the macronuclei as well. These facts argue against the view that the mutants represent simple micronuclear deletions. We conclude that A"300 and A+' are processing mutants, i e . , modified micronuclear A genes that yield dele- tions at specified regions when the micronuclei pro- duce macronuclei at autogamy and conjugation. How- ever, final proof requires isolation and characteriza- tion of the micronuclear A gene from d12.
Alleles that affect serotype specificity, as well as alleles affecting other traits, are normally extremely stable in Paramecium. In spite of intensive investiga- tions there are no reports, to our knowledge, of changes in specificity alleles. However, there are nu- merous reports of changes in the alleles that affect the position of the macronuclear A gene deletions, such as the d l 2 alleles. This lability evidently also applies to d l and to the d l 6 mutant described by EPSTEIN and FORNEY (1 984). d 16 was isolated in a screen for A- mutants, hence presumably was defective at the A
locus when first isolated. Later, however, the deletion point was found to be just downstream of the 3' end of the A gene, and the capacity to produce serotype A was then identical to that of wild type. Whether these frequent changes can be accounted for by mo- bile elements or by some other means will have to await molecular characterization of the micronuclear copies of the various alleles of the A gene.
Mutants Affecting DNA Processing 55
micronuclear alleles to undergo proper processing because of macronuclear deficiencies in d48 or d12. Direct evidence for the sensitivity of the wild-type A
gene processing to the macronuclear environment is provided by the observation that new lines with all the properties of d48 may be artificially constructed from d l 2 by simply replacing its micronuclei with micronuclei from 51 wild-type cells (KOIZUMI and
KOBAYASHI 1990).
This improper processing yields deletions in the A'' gene sometimes, but not always. A number of factors can be identified which influence the probability of occurrence of deletions.
First, The major factor is whether the old macron- ucleus in the cell in which autogamy or conjugation is taking place contains a downstream region of the A gene ( Y o u et al. 1991; unpublished work in our labo- ratory). If this region is present in sufficiently high numbers, as it is in wild type, then the deletions never occur. If the region is not present in high numbers, as in the case in d48, the deletions usually occur.
No satisfactory hypothesis has been suggested to account for this effect. KOIZUMI and KOBAYASHI ( 1989) provided evidence that 5 1 wild-type cytoplasm could rescue d48, provided it is taken from autoga- mous cells and injected into autogamous cells. How- ever, no smaller transcripts from the A gene that might encode a cytoplasmic factor have been detected (H. JESSOP-MURRAY, unpublished results). Perhaps portions of the A genes themselves may be liberated from the old degenerating macronucleus and play a direct role in DNA processing. Alternatively, they may act to bind a DNA processing factor to which the newly forming wild-type A genes are sensitive, thereby sequestering the processing factor and preventing it from making aberrant cleavages. However, the as- sumption that cytoplasmic DNA molecules bring about rescue does not fit well with the finding of
KOIZUMI and KOBAYASHI (1989) that injection of the cloned A gene into autogamous d48 cells gives only low level rescue.
Whatever the mechanism of protection provided by the A genes in the old macronucleus, it is clear that when it fails because of deletions starting at either
+
1or -1300 in the old macronucleus, the new macron- ucleus shows only the deletions starting at + l . This indicates that the section of DNA extending from
-
1300 to +1, which is present in+
1 macronuclei and missing in -1300 macronuclei, has no role in the determination of the deletion site. This result agrees with the finding in our laboratory (H. JESSOP-MUR- RAY, unpublished results) that this region is not able to prevent breaks and effect rescue of d48. Only a downstream portion of the A gene is effective in this way.Second, a major exception to the dependence of proper processing on the presence of downstream A
W
x
d4a W X d 1 2 d40 X d l 2P
FER
MAC DEV
I I
FIGURE 7.-A model to explain and summarize the results of crosses. The two kinds of d l 2 give similar results in crosses. It is assumed that a downstream portion of the wild-type A allele and the mutant alleles, A+' and A"3"" (designated in the figure as a )
produce a factor necessary for proper processing. Production of the factor occurs only during conjugation and autogamy. The A
genes do not produce the factor in the micronuclei and only rarely produce it in macronuclear anlagen (explaining the rare production of A+ clones from both exconjugants in the cross of d48 X 51). Production of the factor regularly takes place in macronuclear fragments. In wild type the factor passes through the cytoplasm and into the macronuclear anlagen. By contrast, in d48 the macronu- clear fragments lack the A gene and produce no factor. Therefore
A genes are usually not included in the new macronuclei forming in d48 cells when d48 conjugates with wild type. On the other hand, the mutant alleles in d l 2 usually produce the factor in early anlagen of heterozygotes and normal processing of A genes occurs. Since the mutant d l 2 alleles cannot respond to the factor, most are deleted from macronuclei before macronuclear formation is com- pleted. Nothing is known about the nature of the factor. For
example, it is possible that the alleles themselves may act as the factor.
gene sequences in the old macronucleus is provided by the behavior of the wild-type A'' allele when it is in heterozygous combination with the A+' or A"300 d l 2 alleles. Wild-type micronuclear alleles in such heterozygotes are often properly processed in spite of a reduced number of A genes in the old macronucleus. It is surprising that a mutation that causes defective processing has a "protective" effect on wild-type genes.
the d48 exconjugant changes quite frequently to A+. Thus, a migratory gametic nucleus from 5 1 wild type yields more intact chromosomes in d48 than a migra- tory gametic nucleus from d48, even though both nuclei contain the wild type A’’ allele. Furthermore, in the cross d l 2 X 5 1, the heterozygous d l
2
excon- jugant always changes to A+ in the F1 and gives a 1:l ratio of A+:A- in the F2 by autogamy. In the d l 2 Xd48 cross, however, the d l 2 exconjugants, which are supposedly identical with the d l
2
exconjugants of the d l2
X 5 1 cross, often produce A- F1 clones and give an excess of A- progeny at the next autogamy. In both these examples, the A5‘ wild-type allele received from a 51 wild-type cell is less likely to undergo deletions than a A” wild-type allele received from a d48 cell. These differences do not appear to be due to the transfer of cytoplasm between conjugants that occurs when cytoplasmic bridges are produced by “delayed separation” (SONNEBORN 1975), although the possibility that minute amounts of cytoplasm are ex- changed cannot be eliminated. Another possibility is that the micronucleoplasm derived from 5 1 wild-type cells acts much like the macronucleoplasm from 51wild-type cells to prevent deletions in developing ma- cronuclei. A similar effect was demonstrated in the case of mating type inheritance in P. tetraurelia (BRY- GOO et al. 1980). They used crosses of different mating types to amicronucleate lines to demonstrate the ef- fect. In view of these considerations, it is probably more accurate to conclude that the non-Mendelian phenomenon is due to effects of both micronucleo- plasm and macronucleoplasm, rather than simply ma- cronucleoplasm.
Figure ’7 represents an attempt to provide a sum- mary of the results of the crosses and a possible explanation as detailed in the figure legend. It should be emphasized that this working model is only one of several possible explanations.
Related phenomena: T h e involvement of the old macronucleus in the formation of the new macronu- cleus at autogamy and conjugation is likely to be of general occurrence and not simply restricted to a few specialized cases like the A gene. T h e inheritance of mating type in P. tetraurelia has been extensively studied (reviewed in SONNEBORN 1975) and the effect of the old macronucleus on the new macronucleus in mating type determination is well established. Even the mating type “selfers” have their parallel in the “mixed” clones (see MATERIALS AND METHODS) which
contain both wild type and d48 cells. Similar systems
of inheritance appear to exist in the case of the tricho- cyst mutants in P . tetraurelia (SONNEBORN and SCHNELLER 1979) and in a case involving surface proteins in Tetrahymena thermophila (DOERDER and BERKOWITZ 1987). Moreover, preliminary experi-
ments (our unpublished results) on new mutants una- ble to express surface proteins B and D also seem to show non-Mendelian as well as Mendelian inheritance. Since all the deletions of the A gene remove the entire end of the macronuclear chromosome bearing the A gene, prerequisites for the phenomenon may be a location adjacent to nonessential downstream se- quences and proximity to the telomere. Deletions might otherwise prove to be lethal.
We thank GUY HAMILTON for technical assistance and JAMFS D. FORNEY for the original cultures of mutants d l and d l 2. This work was supported by U.S. Public Health Service grant GM 31745 from the National Institutes of Health.
LITERATURE CITED
BRYGOO, V., T . M. SONNEBORN, A. M. KELLER, R. V. DIPPELL and M. V. SCHNELLER, 1980 Genetic analysis of mating type dif- ferentiation in Paramecium tetraurelia. 11. Role of the micron- ucleus in mating type determination. Genetics 94: :951 -959.
DOERDER, F. P., and M. S. BERKOWITZ, 1987 Nucleo-cytoplasmic interaction during macronuclear differentiation in ciliate pro- tists: genetic basis for cytoplasmic control of SerH expression during macronuclear development in Tetrahymena thermophila.
Genetics 117: 13-23.
EPSTEIN, L. N., and J. D. FORNEY, 1984 Mendelian and non- Mendelian mutations affecting surface antigen expression in
Paramecium tetraurelia. Mol. Cell. Biol. 4: 1583-1 590.
FORNEY, J. D., and E. H. BLACKBURN, 1988 Developmentally controlled telomere addition in wild-type and mutant parame- cia. Mol. Cell. Biol. 8: 251-258.
FORNEY, J. D., L. M. EPSTEIN, L. B. PREER, B. M. RUDMAN, D. J.
WIDMAYER, W. H. KLEINand J. R. PREER, JR., 1983 Structure and expression of genes for surface proteins in Paramecium.
Mol. Cell. Biol. 3: 466-474.
GODISKA, R., K. J. AUFDERHEIDE, D. GILLEY, P. HENDRIE, T . FITZWATER, L. PREER, B. POLLSKY and J. R. PREER, JR.,
1987 Transformation of Paramecium by microinjection of a cloned serotype gene. Proc. Natl. Acad. Sci. USA 8 4 7590- 7594.
HARUMOTO, T., 1986 Induced change in a non-Mendelian deter- minant by transplantation of macronucleoplasm in Paramecium tetraurelia. Mol. Cell. Biol. 6: 3498-3501.
KOBAYASHI, S., and K. KOIZUMI, 1990 Characterization of non-
Mendelian and Mendelian mutant strains by micronuclear transplantation in Paramecium tetraurelia. J. Protozool. 37:
KOIZUMI, S., and S. KOBAYASHI, 1989 Microinjection of plasmid
DNA encoding the A surface antigen of Paramecium tetraurelia
restores the ability to regenerate a wild-type macronucleus. Mol. Cell. Biol. 9 4398-4401.
SONNEBORN, T. M., 1950 Methods in the general biology and genetics of P. aurelia. J. Exp. Zool. 113: 87-143.
SONNEBORN, T. M., 1975 Paramecium aurelia, pp. 469-594 in
Handbook of Genetics, Vol. 2, edited by R. KING. Plenum Press, New York.
SONNEBORN, T. M., and M. V. SCHNELLER, 1979 A genetic system for alternative stable characteristics in genomically identical homozygous clones. Dev. Genet. 1: 21-46.
You, Y . , K. AUFDERHEIDE, J. MORAND, K. RODKEY and J. FORNEY,
1991 Macronuclear transformation with specific DNA frag- ments controls the content of the new macronuclear genome in Paramecium tetraurelia. Mol. Cell. Biol. 11: 1133-1 137.
Communicating editor: S . L. ALLEN