Template RNA
Ralph T. Mosley,aThomas E. Edwards,bEisuke Murakami,aAngela M. Lam,aRena L. Grice,bJinfa Du,aMichael J. Sofia,a Philip A. Furman,aand Michael J. Ottoa
Pharmasset Inc., Princeton, New Jersey, USA,a
and Emerald BioStructures, Bainbridge Island, Washington, USAb
The replication of the hepatitis C viral (HCV) genome is accomplished by the NS5B RNA-dependent RNA polymerase (RdRp),
for which mechanistic understanding and structure-guided drug design efforts have been hampered by its propensity to
crystal-lize in a closed, polymerization-incompetent state. The removal of an autoinhibitory

-hairpin loop from genotype 2a HCV
NS5B increases
de novo
RNA synthesis by
>
100-fold, promotes RNA binding, and facilitated the determination of the first
crys-tallographic structures of HCV polymerase in complex with RNA primer-template pairs. These crystal structures demonstrate
the structural realignment required for primer-template recognition and elongation, provide new insights into HCV RNA
syn-thesis at the molecular level, and may prove useful in the structure-based design of novel antiviral compounds. Additionally, our
approach for obtaining the RNA primer-template-bound structure of HCV polymerase may be generally applicable to solving
RNA-bound complexes for other viral RdRps that contain similar regulatory

-hairpin loops, including bovine viral diarrhea
virus, dengue virus, and West Nile virus.
A
n estimated 180 million persons worldwide are infected with
hepatitis C virus (HCV). Approximately 80% of these will
develop chronic liver disease, and a significant subset will progress
to cirrhosis of the liver and eventually death (20). HCV is a small,
single-stranded, positive-sense RNA virus and, like dengue virus,
bovine viral diarrhea virus, and West Nile virus, is a member of the
Flaviviridae
family of viruses. The nonstructural 5B (NS5B)
pro-tein, a 66-kDa protein of 591 amino acids found at the C terminus
of the virally encoded HCV polyprotein, provides the requisite
RNA-dependent RNA polymerase (RdRp) functionality (32). The
polymerase produces positive RNA strands for encapsidation into
viral particles by using an intermediate negative RNA strand,
which it synthesizes from the initial positive-strand RNA template
provided by the virus. GTP-dependent
de novo
initiation is likely
the preferred mode of nucleotide polymerization
in vivo
(24).The
essential nature of this enzyme to HCV replication makes it an
important target for direct-acting antivirals (DAAs), including
nucleoside-based (NI) and nonnucleoside allosteric (NNI)
inhib-itors (34,
39).
Nearly 100 crystal structures of HCV NS5B have been
re-ported, covering genotypes 1a, 1b, 2a, and 2b, although all
struc-tures lack the C-terminal membrane-anchoring tail (5). HCV
NS5B exhibits the so-called right-hand shape common to many
polymerases, along with readily recognized finger, palm, and
thumb domains (1,
3,
21), that appears in a closed-fist
conforma-tion without sufficient space for the RNA primer-template (Fig.
1A). However, it has been suggested that the closed conformation
is almost suitable for
de novo
initiation, with enough room to bind
a single strand of template RNA and priming nucleotides (37). By
analogy to a bacteriophage
-6 polymerase initiation complex
with GTP and template (4), Tyr448 of a

-hairpin loop spanning
residues 442 to 454 of the thumb domain may stack against the
initiating GTP during
de novo
initiation. Intriguingly, other
RdRps of the
Flaviviridae
family, such as bovine viral diarrhea
virus (7), dengue virus (43), and West Nile virus (26), contain a
similar thumb domain

-hairpin loop descending into the palm
domain with an appropriately situated aromatic residue
analo-gous to Tyr448 in HCV NS5B.
Extensive efforts to obtain a high-resolution crystal
struc-ture of wild-type HCV polymerase in complex with growing
RNA primer-template pairs have proven unsuccessful,
al-though a structure has been reported with a polyuridine
tem-plate in an unproductive conformation (31). The
superposi-tion of NS5B and HIV-1 reverse transcriptase (RT) crystal
structures (17) provided the earliest models for HCV
elonga-tion (1,
3,
21). However, the autoinhibitory

-hairpin loop and
a C-terminal linker blocks the egress necessary for elongation,
and as was observed with HIV-1 RT, the thumb domain has
been predicted to move in the presence of RNA (1,
3). Insights
from more recent RNA-bound complexes of RdRps from
Nor-walk virus (44), poliovirus (15), and foot-and-mouth disease
virus (FMDV) (12), which lack an equivalent

-hairpin loop,
prompted us to evaluate different HCV NS5B constructs in
which this loop had been modified with the goal of obtaining a
high-resolution crystal structure of the mutant HCV
polymer-ase complexed with an RNA primer-template pair.
In the work presented here, we show that the modified HCV
NS5B polymerase has a significant fold increase in
de novo
RNA
synthesis, remains susceptible to chain termination by known
nucleotide inhibitors, and exhibits inhibitory patterns similar
to those of the wild-type enzyme upon the introduction of the
resistance-derived mutation, S282T. Additionally, we describe
three crystal structures, including two different RNA primer
and template strands complexed with the modified HCV
poly-Received14 February 2012 Accepted2 April 2012
Published ahead of print11 April 2012
Address correspondence to Ralph T. Mosley, [email protected], or Thomas E. Edwards, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00386-12
on November 7, 2019 by guest
http://jvi.asm.org/
merase. These provide the first structure-based insights into
how the enzyme interacts directly with RNA and, by extension,
the nucleos(t)ide-derived medicines which terminate the
growing RNA strand. This new information may make it
pos-sible to design even better nucleos(t)ide and nonnucleoside
medicines than those being developed currently. Our approach
to solving these complexes may extend to other structurally
similar viral polymerases.
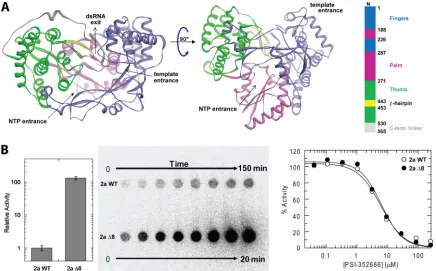
FIG 1Structure of HCV NS5B polymerase and activity of an internal deletion mutant. (A) Crystal structure of genotype 2a HCV NS5B RdRp (37) with the finger, palm, thumb, and C-terminal linker domains numbered and colored according to convention (21). The palm domain is the most well conserved domain across all of the known polymerases and contains the catalytic residues. The thumb domain has the most variability among polymerases and is significantly larger in HCV and otherFlaviviridaeRdRps. This region contains a relatively unique-hairpin loop which descends toward the palm domain, partially blocking what is undoubtedly the exit path for the RNA product strand. This-hairpin loop, colored in yellow, was deleted in the current study. dsRNA, double-stranded RNA. (B)De novoRNA synthesis activity of a genotype 2a JFH1 isolate, wild-type HCV NS5B (2a WT), and a construct in which the-hairpin loop has been deleted and replaced with a Gly-Gly linker (2a⌬8), demonstrating⬎100-fold higher total activity for 2a⌬8 than for the 2a WT. The time-dependent formation of the radiolabeled products is shown in the blot. At the right, the activity for both 2a WT and 2a⌬8 were measured in the presence of the nucleotide triphosphate analog inhibitor PSI-352666, which resulted in an IC50of 6.05⫾0.82M for 2a WT and 6.41⫾0.75M for 2a⌬8.
FIG 2Chain termination of HCV NS5B 1b⌬8. (A) RNA synthesis and chain termination by PSI-352666 for the HCV NS5B polymerase 1b⌬8 construct. GG primer (0.5M) was preincubated with 5M RNA template (3=-CCGGCUGUUAUUAAUAUGUA-5=), 5M 1b⌬8, and 50M CTP for 15 min to form *GGC. One hundredM GTP or PSI-352666 alone or in combination with 100M ATP then was added. The PSI-352666-terminated product (*GGCX) was not further elongated in the presence of the next correct incoming nucleotide (ATP). Lanes indicate 0-, 2-, 5-, 10-, 20-, 40-, and 60-min time courses after preincubation. (B) Chain termination of HCV polymerase 1b WT, 1b S282T, 1b⌬8, and 1b⌬8 S282T with PSI-352666 or 2=-C-MeGTP. Assays were performed as described in Materials and Methods. The calculated IC50s show that the 1b⌬8 construct replicates the previously observed resistance at position S282T relative to the wild-type 1b polymerase.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.75.511.66.337.2]MATERIALS AND METHODS
Protein expression and purification.A construct of the wild-type HCV polymerase genotype 2a JFH-1 isolate was designed with the C-terminal 21 amino acids removed and replaced with a noncleavable hexahistidine tag, and it was cloned into a pET-28a-derived vector. Two surface solubi-lization mutations, E86Q and E87Q, were introduced via site-directed mutagenesis (designated 2a WT). The construct (designated 2a⌬8) in which the-hairpin residues 444 to 453 were removed and replaced with a Gly-Gly linker (seeFig. 3A) was designed using GeneComposer (22,35) and engineered using site-directed mutagenesis. Both mutated constructs were transformed into Rosetta (DE3)Escherichia colicells (Novagen). Recombinant protein was expressed in the Overnight Expression Autoin-duction system (Novagen) at 22°C overnight. Cells were harvested by centrifugation at 5,000⫻gfor 20 min, and the cell pellet was resuspended in 20 mM Tris, pH 8.0, 500 mM NaCl, 20% glycerol, 2 mM Tris(2-car-boxyethyl)phosphine (TCEP), 10 mM imidazole. The lysate was stirred in the presence of benzonase and egg white lysozyme for 1 h at 4°C, followed by clarification by centrifugation at 18,000 rpm for 40 min. The protein was purified by nickel immobilized affinity chromatography, resulting in
⬃90% pure protein as determined by Bio-Rad Experion capillary gel electro-phoresis and SDS-PAGE analysis. The protein was concentrated to 3.9 to 4.4 mg/ml (⬃60M) in 20 mM Tris, pH 8.0, 500 mM NaCl, 10% glycerol, 2 mM TCEP, and⬃200 mM imidazole.
HCV activity assay.Activity assays were performed in a 200-l mix-ture containing 1M the four natural ribonucleotides, [␣-32P]UTP, 20 ng/l of genotype 1b (⫺) 341-nucleotide internal ribosomal entry site (IRES) RNA template (36), 1 U/l of Superase In (Ambion, Austin, TX),
40 ng/l of NS5B, 5 mM MgCl2, and 2 mM dithiothreitol (DTT) in 50 mM HEPES buffer (pH 7.5). Activity assays with genotype 1b NS5b-de-rived proteins were performed in a 120-l mixture containing 5M the four natural ribonucleotides. The reaction mixture was incubated at 27°C, and 20-l aliquots were taken at the desired time points and quenched by mixing in 80l of stop solution (12.5 mM EDTA, 2.25 M NaCl, and 225 mM sodium citrate). Inhibition assays were performed in a 20-l mixture containing various concentrations of PSI-352666 or 2=-C-methyl GTP (MeGTP), 5M the four natural ribonucleotides, [␣-32P]UTP, 20 ng/l of genotype 1b (⫺) 341-nucleotide IRES RNA template, 1 U/l of Su-perase In (Ambion, Austin, TX), 40 ng/l of NS5B, 5 mM MgCl2, and 2 mM DTT in 50 mM HEPES buffer (pH 7.5). The reaction mixture was incubated at 27°C and quenched by adding 80l of stop solution after 60 min of incubation with 2a WT or 15 min of incubation with 2a⌬8. The radioactive RNA products were separated from unreacted substrates us-ing a Hybond N⫹membrane (GE Healthcare) as described previously (29). The products were visualized and quantified using a phosphor-imager. The reaction rates and the 50% inhibitory concentrations (IC50s) were calculated using GraphFit (Erithacus Software, Horley, Surrey, United Kingdom).
Crystallization and structure determination.Crystals were grown using the sitting-drop vapor diffusion method in 96-well-format Com-pact Junior crystallization plates (from Emerald BioSystems) using 0.4l of protein solution and an equal volume of precipitant equilibrated against 80l of precipitant at 16°C. Rod-shaped crystals (20 by 20 by 120
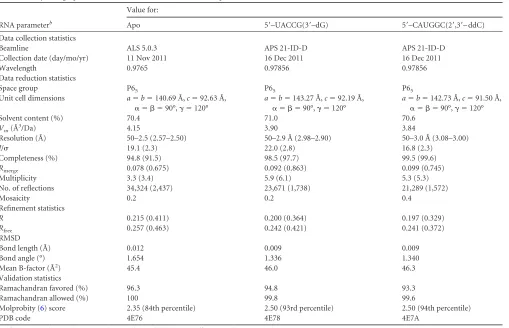
[image:3.585.42.551.75.403.2]m3) appeared within 3 to 5 days in several conditions from the JCSG⫹ (Emerald BioSystems) and the Index (Hampton Research) sparse matrix TABLE 1Crystallographic statistics for HCV NS5B 2a JFH1⌬8 apo and RNA-bound structuresa
RNA parameterb
Value for:
Apo 5=–UACCG(3=–dG) 5=–CAUGGC(2=,3=– ddC)
Data collection statistics
Beamline ALS 5.0.3 APS 21-ID-D APS 21-ID-D
Collection date (day/mo/yr) 11 Nov 2011 16 Dec 2011 16 Dec 2011
Wavelength 0.9765 0.97856 0.97856
Data reduction statistics
Space group P65 P65 P65
Unit cell dimensions a⫽b⫽140.69 Å,c⫽92.63 Å,
␣ ⫽  ⫽90°,␥ ⫽120°
a⫽b⫽143.27 Å,c⫽92.19 Å,
␣ ⫽  ⫽90°,␥ ⫽120°
a⫽b⫽142.73 Å,c⫽91.50 Å,
␣ ⫽  ⫽90°,␥ ⫽120°
Solvent content (%) 70.4 71.0 70.6
Vm(Å
3/Da) 4.15 3.90 3.84
Resolution (Å) 50–2.5 (2.57–2.50) 50–2.9 Å (2.98–2.90) 50–3.0 Å (3.08–3.00)
I/ 19.1 (2.3) 22.0 (2.8) 16.8 (2.3)
Completeness (%) 94.8 (91.5) 98.5 (97.7) 99.5 (99.6)
Rmerge 0.078 (0.675) 0.092 (0.863) 0.099 (0.745)
Multiplicity 3.3 (3.4) 5.9 (6.1) 5.3 (5.3)
No. of reflections 34,324 (2,437) 23,671 (1,738) 21,289 (1,572)
Mosaicity 0.2 0.2 0.4
Refinement statistics
R 0.215 (0.411) 0.200 (0.364) 0.197 (0.329)
Rfree 0.257 (0.463) 0.242 (0.421) 0.241 (0.372)
RMSD
Bond length (Å) 0.012 0.009 0.009
Bond angle (°) 1.654 1.336 1.340
Mean B-factor (Å2) 45.4 46.0 46.3
Validation statistics
Ramachandran favored (%) 96.3 94.8 93.3
Ramachandran allowed (%) 100 99.8 99.6
Molprobity (6) score 2.35 (84th percentile) 2.50 (93rd percentile) 2.50 (94th percentile)
PDB code 4E76 4E78 4E7A
aValues in parentheses indicate the highest-resolution shell. Twenty shells were used in XSCALE (18). b
Rmerge⫽ ⌺|I⫺ ⬍I⬎|/⌺I⫻100, whereIis the intensity of a reflection and⬍I⬎is the average intensity.R⫽ ⌺|Fo⫺Fc|/⌺|Fo|⫻100. A unique 5% of the reflections were used to
calculate theRfreefor each structure. VM is the Matthews coefficient, the crystal volume per unit of protein molecular weight (http://www.ruppweb.org/Mattprob/).
on November 7, 2019 by guest
http://jvi.asm.org/
screens. The apo NS5B 2a⌬8 structure was obtained from a crystal grown in the presence of 30% pentaerithritol ethoxylate, 0.1 M bis-Tris, pH 6.5, and 50 mM ammonium sulfate (Index E9). Apo NS5B 2a⌬8 crystals grown in the presence of 25% polyethylene glycol 3350, 0.1 M bis-Tris, pH 5.5 to 6.5, and 0.2 M ammonium acetate (Index G6-G7) were soaked overnight at 16°C with 0.2 mM 5=UACCG 3=-deoxyguanosine (3=dG) or 5=-CAUGGCdideoxycytosine (ddC) (Dharmacon), precipitant, and 15% ethylene glycol as a cryoprotectant. NS5B 2a⌬8 crystals grown or soaked in the presence of morpholineethanesulfonic acid (MES) buffer were in-compatible with RNA binding. Crystals were harvested and flash-frozen in liquid nitrogen for cryocrystallography. The apo data set was collected at the Advanced Light Source (ALS 5.0.3), and the RNA-bound data sets were collected at the Advanced Photon Source (APS LS-CAT 21-ID-G). The data were reduced in XDS/XSCALE (18). The structures were solved by molecular replacement in PHASER (28) using a previously deter-mined apo structure of a HCV NS5B 2a triple mutant (unpublished), which in turn was solved by molecular replacement using the wild-type HCV NS5B 2a structure (Protein Data Bank [PDB] code 2XXD). The final models were produced after numerous reiterative rounds of refinement in REFMAC5 (30) and manual model building in COOT (11). Structures were assessed for correctness and validated using Mol-probity (6). Figures were generated with PyMol (9).
Protein structure accession numbers.Atomic coordinates and struc-ture factors have been deposited in the Protein Data Bank under accession codes 4E76 (2a⌬8 apo), 4E78 [2a⌬8 with RNA1, 5=-UACCG(3=-dG)], and 4E7A [2a⌬8 with RNA2, 5=-CAUGGC(ddC)].
RESULTS AND DISCUSSION
Based on the
in silico
superposition of the more recent
RNA-bound complexes of RdRps from Norwalk virus (44), poliovirus
(15), and foot-and-mouth disease virus (FMDV) (12), which lack
an equivalent

-hairpin loop, we synthesized an initial HCV NS5B
1b BK construct in which residues 442 to 456 had been excised.
While this 1b
⌬
14 polymerase had reduced activity (data not
shown), it provided the foundation for our interest in a report that
the replacement of residues 444 to 453 (
⌬
8) in this

-hairpin loop
with a Tyr-Gly linker in HCV NS5B genotype 1b (designated 1b
⌬
8) resulted in a 17-fold increase in primer extension activity
above that of the wild type, and the ability to bind
primer-tem-plate RNA (16,
25). We observed a
⬎
10-fold increase in
de novo
RNA synthesis for 1b
⌬
8 with a Gly-Gly linker (data not shown),
as well as evidence for RNA binding via thermofluor analysis (8,
27, and data not shown). We found that this construct was
sus-ceptible to chain termination (Fig. 2), and similar IC
50s were
ob-tained for the GTP analogues PSI-352666 (4
M) and 2
=
-C-MeGTP (7 to 13
M) for both wild-type 1b (1b WT) and 1b
⌬
8
(Fig. 2). Furthermore, upon the introduction of the most
com-monly identified resistance mutation, S282T, similar inhibitory
patterns for either the 1b WT or the 1b
⌬
8 construct were noted for
these same compounds (IC
50for PSI-352666, 9 to 14
M, an
⬃
2.5-fold increase; IC
50for 2
=
-C-MeGTP,
⬎
100
M, a
⬎
10-fold increase)
(Fig. 2) (14). Although the biological characterization demonstrated
that 1b
⌬
8 was functionally similar to 1b WT, we were unable to
obtain a crystal structure of apo 1b
⌬
8 or of it complexed with RNA.
The JFH1 isolate of genotype 2a is the only HCV strain capable
of efficient replication in cell culture as well as
in vivo
(37). A
construct of 2a JFH1 NS5B with the

-hairpin loop (residues
N444 to S453) replaced with a Gly-Gly linker (designated 2a
⌬
8)
was observed to be
⬎
100-fold more active than wild-type 2a in
de
novo
RNA synthesis assays (Fig. 1B), capable of binding RNA in
thermofluor analysis (data not shown), and resulted in a similar
IC
50for chain termination with PSI-352666 (6
M) relative to
wild-type 2a (Fig. 1B). We obtained a 2.5-Å resolution apo crystal
structure of 2a
⌬
8 (Table 1) which revealed substantial structural
changes relative to previously determined 2a NS5B structures (2,
37,
38), with an overall root mean square deviation (RMSD) value
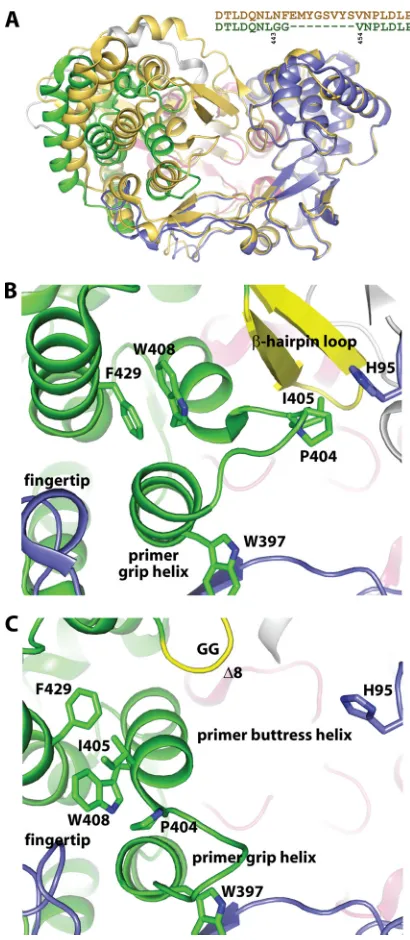
FIG 3Comparison of crystal structures depicting structural changes upon dele-tion of-hairpin loop. (A) Overlay of a previously determined closed crystal struc-ture of genotype 2a HCV NS5B, PDB code 2XXD (37), shown in gold ribbons with the 2.5-Å-resolution crystal structure of genotype 2a HCV NS5B⌬8 polymerase determined here and colored as described forFig. 1. The sequence alignment excerpt explicitly depicts residues excised from the-hairpin loop and replaced with a GG linker. Residues 62 to 350 of the finger and palm domains were aligned to demonstrate the large movement in the thumb domain. (B) Interactions of the thumb domain in the closed 2a WT structure (37). (C) Interactions of the thumb domain in the open 2a⌬8 structure showing the dramatic rearrangement of loop residues 397 to 405 that connect the primer buttress helix with the primer grip helix as well as movement of several helixes in the thumb domain. The structures in panels B and C were overlaid as described for panel A and are shown from the same orientation.on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.317.522.63.533.2]of 1.8 Å (Fig. 3A). The 2a
⌬
8 Gly444-Gly445 linker was ordered,
and Phe551 was the last ordered residue, indicating that an
addi-tional 27 amino acids of the C terminus, including the affinity tag,
were disordered. The alignment of the palm and finger domains of
a closed apo 2a structure (37) with the apo 2a
⌬
8 structure shows
an overall
⬃
20° movement of the thumb domain (Fig. 3A). The
lack of the

-hairpin loop, the disorder of the C-terminal linker
region, and the movement of the thumb domain combine to
gen-erate a large cavity in the center of the polymerase. The thumb
domain movement is accompanied by the significant reordering
of residues 397 to 412, which connect the primer grip helix with
the primer buttress helix (Fig. 3). In particular, Ile405, which is a
valine in other HCV strains and was previously detailed to be
important for
de novo
initiation across all genotypes (37), moved
more than 12 Å away from the

-hairpin loop in the closed
wild-type structure to extend the primer buttress helix and pack on top
FIG 4Crystal structures of 2a⌬8 HCV NS5B with primer-template RNA. (A) Cartoon representation of the structure viewed from the template RNA entrance tunnel, colored as described forFig. 1. The template strand is in salmon, while the primer strand is colored in cyan. (B) View from the RNA exit tunnel of the polymerase. (C) Overlay of the crystal structures of 2a⌬8 with the symmetrical primer-template RNA1 and RNA2. Despite different sequences and chain terminators, both primer-template pairs reside within the central cavity in the same general conformation. (D) Based on a previous report (16,25), we designed a series of symmetrical primer-template RNA constructs. Bases in the template strand are salmon and in the primer are cyan; bases in gray were not observed in the electron density analysis. Base-pairing regions are highlighted with a green background and are numbered by polymerase convention. (E) Overlay of the apo 2a⌬8 HCV NS5B, shown in gold ribbons, with the 2a⌬8 HCV NS5B complexed with RNA1, colored as described forFig. 1, with the template strand in salmon and primer strand in cyan. The alignment of residues 62 to 350 shows modest movement in the thumb domain to accommodate the RNA dimer with the concomitant additional disorder of the C terminus (L545 versus F551 of apo 2a⌬8 versus S563 of apo 2a WT 2XXD).on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.75.511.63.517.2]of the highly conserved Trp408 (Fig. 3). Trp408 stacked on top of
the nearly invariant Phe429 in the closed wild-type structure, and
both residues adopt different rotamer conformations in the 2a
⌬
8
structure. In addition, the highly conserved Pro404, which
con-tacts His95 of the finger domain in the closed apo structure (Fig.
3B), forms a key turn in the loop while packing on top of the main
chain of Trp397 in the apo 2a
⌬
8 structure (Fig. 3C). This loop
reordering may be critical to the transition from
de novo
initiation
with GTP to the elongation of the growing primer-template RNA.
Intriguingly, this same loop reordering was observed in the 2.5-Å
structure of an apo 2a JFH1 isolate HCV NS5B resistance-derived
triple mutant (S15G/C223H/V321I) (19) in which the

-hairpin
loop is extant (unpublished data). The comparison of the apo 2a
⌬
8 structure to other RdRp ternary complexes (12,
15,
44)
sug-gested that in this open conformation, HCV NS5B is able to bind
primer-template RNA.
Crystals of apo 2a
⌬
8 were soaked with self-annealing RNA,
yielding symmetrical primer-template RNA with 5
=
overhangs,
and structures were determined at 2.9- and 3.0-Å resolution (Fig.
4
and
Table 1). A-form RNA was readily apparent in the resulting
electron density maps (Fig. 5), clearly showing the differences in
the purine/pyrimidine pairings of the two RNA sequences. Both
symmetrical primer-template RNA pairs were designed as
obli-gate chain terminators with either 3
=
-dG (RNA1) or 2
=
,3
=
-ddC
(RNA2), and thus, unsurprisingly, a product state,
pretransloca-tion registry, was observed in both complexes (Fig. 4). In both
structures, four bases are paired in the primer-template RNA
dimer with a 5
=
1-base overhang observed within the electron
density map. None of the nucleobase hydrogen bond acceptors or
donors is recognized by the polymerase, indicating
sequence-in-dependent recognition by the polymerase. The interactions
be-tween the polymerase and the nucleic acids include packing with
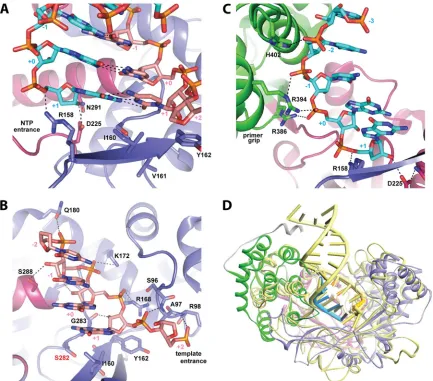
the pairing nucleotide of the template strand (residue
⫹
1 by
con-vention) and residue
⫹
1 of the primer strand in the current,
product-state assembly (equivalent to the incoming nucleoside
triphosphate [NTP]), as well as numerous interactions with the
phosphodiester backbone. The sugar and phosphate of the
⫹
2
residue of the template strand head down the template entrance
tunnel. Template-strand phosphates are recognized by the
back-bone amide nitrogen atoms of Arg98 (
⫹
2) and Ala97 (
⫹
1) and
the side chains of Arg168 (
⫹
0), Lys172 (
⫺
1), and Gln180 (
⫺
2)
(Fig. 6). The 2
=
-hydroxyl of the pairing nucleotide (
⫹
1) is
recog-nized by the backbone oxygen of strictly conserved Gly283, while
the other 2
=
-hydroxyls of the template strand are recognized by
the backbone oxygen of Val284 (
⫹
0), the side chain of Ser288
(
⫺
1), and possibly the backbone oxygen of Phe193 (
⫺
2) (Fig. 6B),
demonstrating the importance of an RNA template for HCV. The
phosphates of the primer strand are recognized by the side chains
of Arg158 (
⫹
1) of the finger domain, Arg386 and Arg394 (
⫹
0),
Arg394 (
⫺
1), and His402 (
⫺
2) from the primer grip helix in the
thumb domain (Fig. 6C). The primer buttress helix, spanning
residues 405 to 414, has moved away from the central cavity
rela-tive to the closed apo wild-type structure, allowing for strictly
conserved residue Gly410 to come within 3.3 Å of the 2
=
-hydroxyl
of primer-strand residue
⫺
2, demonstrating that any other amino
acid at this position would likely clash with the primer. The 2
=
-hydroxyl of primer residue
⫹
1 of the product, pretranslocation
state, which resides at the same position as the incoming NTP in
the substrate registry, is recognized by the side chain of Asp225
(Fig. 6A). Since both structures contain a 3
=
-deoxy terminal
resi-due, the other carboxylate oxygen of Asp225 is free to hydrogen
bond with Asn291. The equivalent residue (Asp238) of the
polio-virus RdRp was shown to adopt different conformations
depend-ing on the incomdepend-ing NTP, translocation state, and presence of
divalent metal ions (15). None of the other 2
=
-hydroxyls of the
primer strand are recognized by NS5B, which is consistent with
reduced activity with DNA primers (42). Despite low sequence
identity outside the catalytic residues, the overall primer-template
RNA recognition strategy of HCV is essentially identical to that
observed for Norwalk virus (44), poliovirus (15), and FMDV (12)
(Fig. 6D).
Although the nucleobase hydrogen bond acceptors or donors
are not recognized by the polymerase, it does appear that the
poly-merase is able to differentiate between a purine and pyrimidine
base at the
⫹
1 position in the RNA template strand, as seen in
these two complexes of 2a HCV NS5b
⌬
8 with RNA1 and RNA2.
The nucleobase of the pairing nucleotide of the template strand
(residue
⫹
1) stacks on top of the strictly conserved Ile160, as
predicted (3), while the sugar stacks on top of Tyr162, which is
conserved as Tyr or Phe (Fig. 7). In the RNA1 complex, the C
␥
2 of
the Ile160 side chain appears to interact with the face of the C3
base, the
⫹
1 position of the template RNA, while the C
␦
1 of the
side chain extends beyond the C3 base to interact with the
nucleo-base of dG6, the
⫹
1 position of the primer RNA (Fig. 7A).
Con-versely, in the RNA2 complex, both C
␥
2 and C
␦
1 of Ile160 appear
to interact only with the bulkier nucleobase of G4, the
⫹
1 position
of the template strand, and do not seem capable of interacting
directly with the necessarily more distant ddC7, the
⫹
1 position in
the RNA2 primer strand (Fig. 7B). To our knowledge, this is the
first crystallographic evidence for such differentiation in an
RdRp-RNA complex: in the poliovirus RdRp-RdRp-RNA complexes (15),
which have the analogous Ile176, the
⫹
1 RNA template base is
always a purine; in the norovirus RdRp-RNA complexes (44),
with the analogous Leu184, the
⫹
1 RNA template base is also
always a purine. Only in the FMDV-RNA complexes (13), with the
analogous Val181, which is too small to extend beyond the pairing
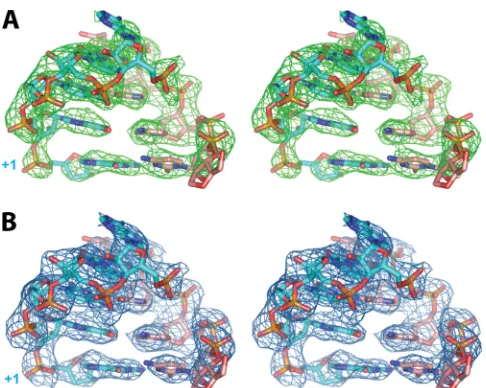
FIG 5Relaxed-eye stereo view of omit electron density map |Fo|-|Fc|, intowhich the symmetrical primer-template RNA1 5=-UACCG(3=-dG) model was built (shown in green mesh) contoured at 3.0(A), and the final 2|Fo|-|Fc| electron density map is shown in blue mesh contoured at 1.0(B).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.42.285.63.257.2]nucleobase, does the
⫹
1 RNA template base vary between purine
and pyrimidine. If these same observations are made in additional
HCV RdRp-RNA complexes, it will be intriguing to speculate how
such a differentiation is evidenced in the efficiency of the enzyme
as it relates to the incorporation of natural nucleotides or various
NIs during initiation or elongation or when perturbed by nearby
mutations, such as S282T, as has been previously documented
(10).
In addition to demonstrating the molecular basis for
primer-template recognition and elongation by HCV polymerase, these
structures may provide insight into the structural basis by which
resistance-derived mutations permit the polymerase to continue
to function while diminishing the impact of the inhibitor, be it an
NI or NNI. For example, two of the most frequently arising
resis-tance mutations for NIs in HCV are S96T and S282T. In our
struc-tures, Ser96 hydrogen bonds with Arg168, which directly
posi-tions the
⫹
0 phosphate of the template strand and indirectly
positions the pairing nucleotide phosphate (Fig. 6B). Mutation to
threonine likely repositions the template strand toward the thumb
domain, decreasing the space available and thereby increasing the
preference for the incoming NTP over the typically bulkier NI.
With regard to the S282T mutant, the threonine has been
specu-lated to directly impede the incoming 2
=
-C-MeNTP as its mode of
action (10). However, proximity to two nearby residues, the
afore-mentioned Ile160 and Gly283, which interacts with the 2
=
-hy-droxyl of template RNA
⫹
1 ribose, suggests a more complicated
picture for S282T-derived resistance. One can also imagine a
sce-nario in which resistance-derived mutations are modeled into
these new structures, potentially providing insights into which
mutated residues block access to an inhibitory binding cavity and
which allow the protein to compensate for concomitantly reduced
efficacy.
The more open nature of the primer-template-bound
com-plexes described here also offers a more complete glimpse into the
FIG 6Primer-template recognition by HCV polymerase. (A) The nucleobase of the pairing nucleotide (⫹1) stacks on top of the highly conserved Ile160 while base pairing with the primer strand in the product, pretranslocation state. The 3=residue of the primer strand contains the obligate chain terminator 3=-dG and resides in the pretranslocation state, where one would expect the incoming NTP to bind after translocation. (B) All of the phosphates and 2=-hydroxyls of the template strand are recognized by HCV polymerase. The main source of resistance to certain nucleotide analog inhibitors occurs at Ser282, directly below the terminal nucleotide of the primer strand. (C) Interactions of the polymerase with the primer strand. (D) Overlay of a primer-template RNA-bound crystal structure of HCV polymerase, colored as described forFig. 4, with a primer-template RNA-bound crystal structure of poliovirus polymerase, PDB code 3OL6 (15), colored yellow, showing similar overall primer-template-polymerase recognition despite substantially different thumb domains.on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.76.509.65.446.2]mode of action for some allosteric inhibitor classes. Most of the
palm domain binding allosteric NNIs stabilize the

-hairpin loop
via critical interactions with Tyr448, thereby inhibiting the
poly-merase in both the initiation and elongation stages (33). Our
structures underscore the necessity of retracting the

-hairpin
loop for elongation to proceed. Allosteric inhibitors which target
thumb site I, the region into which Leu30 and Leu31 of the finger
loop bind, have been shown to affect the initiation process but are
ineffectual once the RdRp-RNA complex has formed (40).
There-fore, these structures are unlikely to provide additional insights
into this class of NNIs. The second allosteric inhibitor site found
in the thumb domain is
⬃
15 Å removed from thumb site I. The
mode of action for thumb site II NNIs is not known but has been
suggested to be due to the limiting of dynamism (2,
41) and/or
blocking association with other proteins/RNA important for
rep-lication (23). With the large movement of the thumb domain
from the closed form typically observed for HCV NS5B, one can
anticipate that these structures would provide clarity. However,
this region of the thumb domain is nearly identical to that seen for
an inhibited complex, e.g., 1YVZ (2), perhaps lending more
cre-dence to blocking association with other proteins/RNA as the
mode of action for thumb site II NNIs. Clearly, these structures
provide a valuable crystallization platform for structure-guided
drug design, in particular for nucleotide analog inhibitors or NNIs
that target the ternary complex. Finally, the methodology of
de-leting elements of the

-hairpin loop to afford the
primer-tem-plate-bound complex may prove similarly useful through iterative
application to other viral RdRps with this structural feature.
ACKNOWLEDGMENTS
We thank K. Potts for assistance with molecular biology, S. Moen for fermentation, J. Bullen and M. Namekata for protein purification, R. Baydo for thermofluor experiments, M. Clifton for structure peer review, C. Lugo and H. Bao for assisting in biochemical characterization, and N. Dave at ALS and J. Brunzelle at APS LS-CAT for assistance during data collection.
R.T.M., E.M., A.M.L., J.D., M.J.S., P.A.F., and M.J.O. are employees of Pharmasset, Inc., and T.E.E. and R.L.G. are employees of Emerald Bio-Structures, a structural biology CRO contracted by Pharmasset, Inc.
REFERENCES
1.Ago H, et al.1999. Crystal structure of the RNA-dependent RNA poly-merase of hepatitis C virus. Structure7:1417–1426.
2.Biswal BK, et al.2005. Crystal structures of the RNA-dependent RNA polymerase genotype 2a of hepatitis C virus reveal two conformations and suggest mechanisms of inhibition by nonnucleoside inhibitors. J. Biol. Chem.280:18202–18210.
3.Bressanelli S, et al.1999. Crystal structure of the RNA-dependent RNA polymerase of hepatitis C virus. Proc. Natl. Acad. Sci. U. S. A.96:13034 – 13039.
4.Butcher SJ, Grimes JM, Makeyev EV, Bamford DH, Stuart DI.2001. A mechanism for initiating RNA-dependent RNA polymerization. Nature 410:235–240.
5.Caillet-Saguy C, Simister PC, Bressanelli S.2011. An objective assess-ment of conformational variability in complexes of hepatitis C virus poly-merase with non-nucleoside inhibitors. J. Mol. Biol.414:370 –384. 6.Chen VB, et al.2010. MolProbity: all-atom structure validation for
macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66:12–21.
7.Choi KH, et al.2004. The structure of the RNA-dependent RNA poly-merase from bovine viral diarrhea virus establishes the role of GTP in de novo initiation. Proc. Natl. Acad. Sci. U. S. A.101:4425– 4430.
8.Crowther GJ, et al.2010. Use of thermal melt curves to assess the quality of enzyme preparations. Anal. Biochem.399:268 –275.
9.DeLano WL.2002. The PyMOL molecular graphics system. DeLano Sci-entific, San Carlos, CA.
10. Dutartre H, Bussetta C, Boretto J, Canard B.2006. General catalytic deficiency of hepatitis C virus RNA polymerase with an S282T mutation and mutually exclusive resistance towards 2=-modified nucleotide ana-logues. Antimicrob. Agents Chemother.50:4161– 4169.
11. Emsley P, Cowtan K.2004. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr.60:2126 –2132.
12. Ferrer-Orta C, et al.2004. Structure of foot-and-mouth disease virus RNA-dependent RNA polymerase and its complex with a template-primer RNA. J. Biol. Chem.279:47212– 47221.
13. Ferrer-Orta C, et al.2010. Structure of foot-and-mouth disease virus mutant polymerases with reduced sensitivity to ribavirin. J. Virol.84: 6188 – 6199.
14. Furman PA, et al.2011. Activity and the metabolic activation pathway of the potent and selective hepatitis C virus pronucleotide inhibitor PSI-353661. Antiviral Res.91:120 –132.
15. Gong P, Peersen OB.2010. Structural basis for active site closure by the poliovirus RNA-dependent RNA polymerase. Proc. Natl. Acad. Sci. U. S. A.107:22505–22510.
16. Hong Z, et al.2001. A novel mechanism to ensure terminal initiation by hepatitis C virus NS5B polymerase. Virology285:6 –11.
17. Huang H, Chopra R, Verdine GL, Harrison SC.1998. Structure of a covalently trapped catalytic complex of HIV-1 reverse transcriptase: im-plications for drug resistance. Science282:1669 –1675.
18. Kabsch W.2010. Xds. Acta Crystallogr. D Biol. Crystallogr.66:125–132. FIG 7Interactions of Ile160 with the pairing nucleotide of the terminal base
pair during elongation. (A) In the RNA1 crystal structure, where the pairing nucleotide of the template strand is a pyrimidine, Ile160 also interacts with the
⫹1 residue of the primer strand, which is equivalent to the incoming NTP. (B) In the RNA2 crystal structure, the pairing nucleotide of the template strand (⫹1) is a purine, thus Ile160 does not interact with the⫹1 residue of the primer strand.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.61.263.65.394.2]19. Lam AM, et al.2011. Hepatitis C virus nucleotide inhibitors PSI-352938 and PSI-353661 exhibit a novel mechanism of resistance requiring multi-ple mutations within replicon RNA. J. Virol.85:12334 –12342.
20. Lavanchy D.2009. The global burden of hepatitis C. Liver Int.29(Suppl. 1):74 – 81.
21. Lesburg CA, et al.1999. Crystal structure of the RNA-dependent RNA polymerase from hepatitis C virus reveals a fully encircled active site. Nat. Struct. Biol.6:937–943.
22. Lorimer D, et al.2009. Gene composer: database software for protein construct design, codon engineering, and gene synthesis. BMC Biotech-nol.9:36.
23. Love RA, et al.2003. Crystallographic identification of a noncompetitive inhibitor binding site on the hepatitis C virus NS5B RNA polymerase enzyme. J. Virol.77:7575–7581.
24. Luo G, et al.2000. De novo initiation of RNA synthesis by the RNA-dependent RNA polymerase (NS5B) of hepatitis C virus. J. Virol.74:851– 863.
25. Maag D, Castro C, Hong Z, Cameron CE. 2001. Hepatitis C virus RNA-dependent RNA polymerase (NS5B) as a mediator of the antiviral activity of ribavirin. J. Biol. Chem.276:46094 – 46098.
26. Malet H, et al.2007. Crystal structure of the RNA polymerase domain of the West Nile virus non-structural protein 5. J. Biol. Chem.282:10678 – 10689.
27. Matulis D, Kranz JK, Salemme FR, Todd MJ.2005. Thermodynamic stability of carbonic anhydrase: measurements of binding affinity and stoi-chiometry using ThermoFluor. Biochemistry44:5258 –5266.
28. McCoy AJ, et al.2007. Phaser crystallographic software. J. Appl. Crystal-logr.40:658 – 674.
29. Murakami E, et al.2007. Mechanism of activation of beta-D-2= -deoxy-2=-fluoro-2=-c-methylcytidine and inhibition of hepatitis C virus NS5B RNA polymerase. Antimicrob. Agents Chemother.51:503–509. 30. Murshudov GN, Vagin AA, Dodson EJ.1997. Refinement of
macromo-lecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr.53:240 –255.
31. O’Farrell D, Trowbridge R, Rowlands D, Jager J.2003. Substrate com-plexes of hepatitis C virus RNA polymerase (HC-J4): structural evidence for nucleotide import and de-novo initiation. J. Mol. Biol.326:1025–1035.
32. Penin F, Dubuisson J, Rey FA, Moradpour D, Pawlotsky JM. 2004. Structural biology of hepatitis C virus. Hepatology39:5–19.
33. Pfefferkorn JA, et al.2005. Inhibitors of HCV NS5B polymerase. Part 1: evaluation of the southern region of (2Z)-2-(benzoylamino)-3-(5-phenyl-2-furyl)acrylic acid. Bioorg. Med. Chem. Lett.15:2481–2486. 34. Powdrill MH, Bernatchez JA, Gotte M.2010. Inhibitors of the hepatitis
C virus RNA-dependent RNA polymerase NS5B. Viruses2:2169 –2195. 35. Raymond A, et al.2009. Combined protein construct and synthetic gene
engineering for heterologous protein expression and crystallization using Gene Composer. BMC Biotechnol.9:37.
36. Reigadas S, et al.2001. HCV RNA-dependent RNA polymerase replicates in vitro the 3=terminal region of the minus-strand viral RNA more effi-ciently than the 3=terminal region of the plus RNA. Eur. J. Biochem. 268:5857–5867.
37. Schmitt M, et al.2011. A comprehensive structure-function comparison of hepatitis C virus strain JFH1 and J6 polymerases reveals a key residue stimu-lating replication in cell culture across genotypes. J. Virol.85:2565–2581. 38. Simister P, et al.2009. Structural and functional analysis of hepatitis C
virus strain JFH1 polymerase. J. Virol.83:11926 –11939.
39. Sofia MJ, Chang W, Furman PA, Mosley RT, Ross BS.2012. Nucleoside, nucleotide, and non-nucleoside inhibitors of hepatitis C virus NS5B RNA-dependent RNA-polymerase. J. Med. Chem.55:2481–2531.
40. Tomei L, et al.2003. Mechanism of action and antiviral activity of ben-zimidazole-based allosteric inhibitors of the hepatitis C virus RNA-dependent RNA polymerase. J. Virol.77:13225–13231.
41. Wang M, et al.2003. Non-nucleoside analogue inhibitors bind to an allosteric site on HCV NS5B polymerase. Crystal structures and mecha-nism of inhibition. J. Biol. Chem.278:9489 –9495.
42. Yamashita T, et al.1998. RNA-dependent RNA polymerase activity of the soluble recombinant hepatitis C virus NS5B protein truncated at the C-terminal region. J. Biol. Chem.273:15479 –15486.
43. Yap TL, et al.2007. Crystal structure of the dengue virus RNA-dependent RNA polymerase catalytic domain at 1.85-angstrom resolution. J. Virol. 81:4753– 4765.
44. Zamyatkin DF, et al.2008. Structural insights into mechanisms of catalysis and inhibition in Norwalk virus polymerase. J. Biol. Chem. 283:7705–7712.
on November 7, 2019 by guest
http://jvi.asm.org/