Cns1 Cochaperone Is a Novel Restriction Factor of Tombusvirus
Replication in Yeast
Jing-Yi Lin,a,bPeter D. Nagya
Department of Plant Pathology, University of Kentucky, Lexington, Kentucky, USAa
; School of Medical Laboratory Science and Biotechnology, College of Medical Science and Technology, Taipei Medical University, Taiwanb
A large number of host-encoded proteins affect the replication of plus-stranded RNA viruses by acting as susceptibility factors.
Many other cellular proteins are known to function as restriction factors of viral infections. Previous studies with tomato bushy
stunt tombusvirus (TBSV) in a yeast model host have revealed the inhibitory function of TPR (tetratricopeptide repeat)
domain-containing cyclophilins, which are members of the large family of host prolyl isomerases, in TBSV replication. In this paper, we
tested additional TPR-containing yeast proteins in a cell-free TBSV replication assay and identified the Cns1p cochaperone for
heat shock protein 70 (Hsp70) and Hsp90 chaperones as a strong inhibitor of TBSV replication. Cns1p interacted with the viral
replication proteins and inhibited the assembly of the viral replicase complex and viral RNA synthesis
in vitro
. Overexpression
of Cns1p inhibited TBSV replication in yeast. The use of a temperature-sensitive (TS) mutant of Cns1p in yeast revealed that at a
semipermissive temperature, TS Cns1p could not inhibit TBSV replication. Interestingly, Cns1p and the TPR-containing Cpr7p
cyclophilin have similar inhibitory functions during TBSV replication, although some of the details of their viral restriction
mechanisms are different. Our observations indicate that TPR-containing cellular proteins could act as virus restriction factors.
T
he host cell plays a major role in the replication of
plus-stranded RNA [(
⫹
)RNA] viruses, which takes place in
mem-brane-bound viral replicase complexes (VRCs) in the cytoplasm
of infected cells. To aid in viral replication, (⫹)RNA viruses coopt
a number of host-encoded proteins (
1–10
). However, the host
also restricts viral infections based on the expression of various
restriction factors and several groups of cellular proteins involved
in innate immune responses of the host (
11–14
). Accordingly,
genome-wide screens with various hosts and viruses, such as
to-mato bushy stunt virus (TBSV), West Nile virus, brome mosaic
virus (BMV), hepatitis C virus (HCV), dengue virus, and
Drosoph-ila
virus C, led to the identification of stimulatory as well as
inhib-itory host proteins (
15–23
). The functions of the majority of the
identified host proteins in (⫹)RNA virus replication have not
been fully revealed.
TBSV is a small-model (
⫹
)RNA virus used to study virus
rep-lication, recombination, and virus-host interactions based on a
yeast (
Saccharomyces cerevisiae
) model host (
24–27
). Several
ge-nome-wide screens of yeast genes and different global proteomics
approaches have led to the identification of over 500 host genes/
proteins that might be involved in TBSV replication or
recombi-nation (
15
,
17
,
28–37
). TBSV uses the peroxisomal membrane to
assemble the tombusvirus VRC, consisting of 2 viral replication
proteins (i.e., p33 and p92
pol) and 8 to 10 host proteins (
35–38
).
These host proteins have been shown to bind to the viral RNA and
the viral replication proteins (
1
,
37
,
39
). For example, heat shock
protein 70 (Hsp70), eukaryotic elongation factor 1A (eEF1A), and
the ESCRT (endosomal sorting complexes required for transport)
family of host proteins are required for the assembly of the VRC
(
40–43
). Additional host proteins, such as the Ded1 DEAD box
helicase, glyceraldehyde-3-phosphate dehydrogenase (GAPDH),
eEF1B
␥
, and eEF1A, have been shown to affect viral RNA
synthe-sis (
39
,
40
,
44–46
). The multifunctional p33 replication protein
plays a role in recruitment of the TBSV (⫹)RNA to the site of
replication, the assembly of the VRC, and viral RNA synthesis by
acting as an RNA chaperone (
38
,
47–50
). The RNA-dependent
RNA polymerase (RdRp) protein p92
polbinds to the essential p33
replication protein within the functional VRC (
26
,
49
,
51
,
52
).
Our knowledge on host regulatory or intrinsic restriction
fac-tors of TBSV replication is also growing rapidly. For example,
nucleolin, an RNA binding protein, interferes with the
recruit-ment of the viral RNA to the replication site (
53
). Rsp5p, an E3
ubiquitin ligase in the Nedd4 family, and other WW
motif-con-taining host proteins regulate the degradation of p92
polin yeast
cells and the activity of the VRC
in vitro
(
54
,
55
). Recent works also
revealed novel restriction functions for selected cyclophilins and
Ess1p parvulin, which decrease TBSV RNA accumulation in yeast
(
34
,
56
).
An especially remarkable restriction factor is the Cyp40-like
Cpr7p cyclophilin, which is a strong inhibitor of TBSV replication
in yeast and
in vitro
(
56
). The inhibitory effect of Cpr7p is due to
its ability to bind to the RNA binding domain of p33 via its TPR
(tetratricopeptide repeat) domain consisting of three TPR repeats
in Cpr7p that leads to inhibition of p33/p92-based recruitment of
the TBSV RNA for replication and a decrease of the efficiency of
VRC assembly. Surprisingly, the second Cyp40-like cyclophilin,
named Cpr6p in yeast, did not inhibit TBSV replication in spite of
the presence of the TPR domain, which bound to the p33 and p92
replication proteins (
56
). These works invited our attention to
TPR-like sequences, which are present in several host proteins.
Received21 January 2013 Accepted8 September 2013
Published ahead of print11 September 2013
Address correspondence to Peter D. Nagy, [email protected]. Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00196-13
on November 7, 2019 by guest
http://jvi.asm.org/
To gain further knowledge on the roles of TPR-containing host
proteins in TBSV replication, in this work, we tested four host
proteins in a cell-free TBSV replication assay. We have identified
the Cns1p cochaperone, which showed strong inhibitory function
in TBSV replication. Cns1p (cyclophilin seven suppressor)
con-tains a TPR domain, which is involved in binding to Hsp90s and
Hsp70s (
57
). Cns1p is known to suppress the growth defect caused
by deletion of Cpr7p (
58
,
59
). Cns1p is essential for yeast growth,
is not induced by heat stress, and cannot be complemented by
overexpression of other yeast cochaperones in
cns1
⌬
yeast strains
(
58
,
59
).
Detailed analysis of Cns1p revealed that it interacted with the
p33 and p92 replication proteins and ultimately restricted VRC
assembly
in vitro
and viral RNA accumulation in yeast. Altogether,
the functional role of Cns1p is reminiscent of the previously
char-acterized role for Cpr7p cyclophilin (
56
). Thus, TPR-containing
host proteins emerge as new host restriction factors of an (
⫹
)RNA
virus.
MATERIALS AND METHODS
Expression plasmids.For TBSV expression, plasmids pHis-GBK-CUP-His33/GAL-DI72 and pGAD-CUP-Hisp92 were used for thein vivoviral replication assay (36,60). For Nodamura virus (NoV) expression, plas-mid pESC-His/Cupm/NoV/RNA1/TRSVrz was used for thein vivoviral replication assay (61).
To generate theEscherichia coliexpression plasmids for glutathione
S-transferase (GST)-Sec17, GST-Pex5, GST-Sti1, and GST-Cns1, the re-striction sites used for ligation into the pGEX-2T vector (New England BioLabs) were BamHI and EcoRI. PCR amplification ofSEC17,PEX5,
STI1, andCNS1open reading frames (ORFs) was performed with primer sets 4849 (GGCGGGATCCATGTCAGACCCTGTAGACTTA)/4850 (CG CCGAATTCTCATAACAAATCATCTTCTTG), 4855 (GGCGGGATCC ATGGACGTAGGAAGTTGCTCA)/4856 (CGCCGAATTCTCAAAACG AAAATTCTCCTTT), 4859 (GGCGGGATCCATGTCATTGACAGCCGA TGAA)/4860 (CGCCGAATTCTTAGCGGCCAGTCCGGATGAT), and 4851 (GGCGGGATCCATGAGCTCCGTTAACGCAAAT)/4852 (CGCCG AATTCTCACACAGATCTTCTTTCTAA), respectively. PCR products were digested with BamHI and EcoRI and ligated into pGEX-2T by using the same enzymes.
To generate yeast plasmids for the split-ubiquitin assay and theCNS1
ORF and its truncated derivatives, the restriction sites used for ligation into the pPRN-N-RE vector (Dualsystems) were BamHI and EcoRI (54). PCR amplification ofCNS1ORF residues 1 to 188, 84 to 188, 84 to 385, and 189 to 385 were performed with primer sets 4851/4852, 4851/5022 (CGCCGAATTCCTCTGGGTCAATCCTTTGATTGGC), 5021 (GGCG GGATCCGAAAATTTCAAGAAGCAAGGTAAC)/5022, 5021/4852, and 5029 (GGCGGGATCCAACAAATCAATTTTGAATATGTTA)/4852, re-spectively. PCR products were digested with BamHI and EcoRI and li-gated into pPRN-N-RE by using the same enzymes.
Analysis of protein-protein interactions using the split-ubiquitin assay.The split-ubiquitin assay is based on the Dualmembrane kit3 (Dualsystems) and was performed as previously described (34,62). The bait constructs pGAD-BT2-N-His33 and pGAD-BT2-N-His92, express-ing tombusvirus replication proteins p33 and p92; pGAD-BT2-N-NoV protA; pGAD-BT2-N-TCV p28; pGAD-BT2-N-RCNMV p27; and pGAD-BT2-N-TMV 130K were described previously (34,36,56). Yeast strain NMY51 was cotransformed with pGAD-BT2-N-His33 and pPR-N-RE (NubG) or one of the prey constructs carryingCNS1and plated onto plates of synthetic minimal medium lacking Trp and Leu (Trp⫺/ Leu⫺) (TL⫺plates) for plasmid selection. Yeast colonies were then resus-pended in 50 l water and spotted onto Trp⫺/Leu⫺/His⫺/Ade⫺ (TLHA⫺) plates to detect bait-prey interactions based on yeast growth at 29°C for 3 to 5 days (34,62). A plasmid carrying theSSA1ORF served as the positive control in this assay (34,62).
Analysis of protein-protein interactionsin vitro.Maltose binding protein (MBP)-tagged p33C was purified fromE. colias described previ-ously (63). Cells were resuspended in cold column buffer (10 mM Tris-HCl [pH 7.4], 1 mM EDTA, 25 mM NaCl, 10 mM-mercaptoethanol) and broken by sonication. The cleared lysate was passed through an am-ylose column to bind the MBP-tagged viral proteins or MBP (negative control). The columns were washed three times with cold column buffer prior to the addition of the yeast lysate (63). For the pulldown assay, 100 mg of yeast pellets containing Flag-tagged Cns1p was resuspended in 150
l chilled buffer I (20 mM Tris-HCl [pH 7.5], 1 mM EDTA, 200 mM NaCl, 10% [vol/vol] glycerol, 0.1% [vol/vol] NP-40, 10 mM -mercapto-ethanol, 1% [vol/vol] yeast protease inhibitor cocktail [Ypic]) and 1l of RNase A (1 mg/ml) (56). Cells were broken in a Genogrinder by using a 250-l volume of acid-washed glass beads for 2 min at 1,500 rpm, fol-lowed by the addition of 600l of buffer I. The detailed procedure was described previously (56). The presence of Flag-Cns1p in the eluate was analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed by Western blotting using an anti-Flag antibody. The amount of MBP-tagged viral proteins in the eluent was visualized by Coomassie blue staining of the SDS-PAGE gels (56).
Purification of GST and GST-tagged Cns1p, Sti1p, Sec17p, and Pex5p proteins was carried out by using glutathione resin, and proteins were eluted with 10 mM glutathione–10 mM-mercaptoethanol in the col-umn buffer according to the protocol used for MBP-tagged proteins (64). Analysis of TBSV and NoV repRNA replication in yeast.For measur-ing TBSV replicon RNA (repRNA) accumulation, yeast strain BY4741 was transformed with plasmids pGAD-CUP-Hisp92, pHis-GBK-CUP-Hisp33/GAL-DI72, and Cns1-zz (yeast ORF collection; Open Biosys-tems). For measuring TBSV repRNA accumulation in yeast strain BY4741 and in its derivative yeast strain carrying temperature-sensitive (TS)CNS1
as the sole copy (65), these yeast strains were transformed with plasmids pGAD-CUP-Hisp92 and pHis-GBK-CUP-Hisp33/GAL-DI72. For mea-suring NoV repRNA accumulation, yeast strain BY4741 was transformed with plasmids pESC-His/Cupm/NoV/RNA1/TRSVrz and Cns1-zz. Rep-lication assays were performed by measuring the accumulation of DI-72 plus-strand repRNA [(⫹)repRNA] or NoV repRNA relative to the 18S rRNA level (61). Replication was induced by adding 50M CuSO4to the
medium, and yeast cells were then cultivated for 24 h at different temper-atures for TBSV and for 48 h at 29°C for NoV.
In vitroreplication assay using whole-cell extract.The yeast strain (BY4741) used for cell-free extract (CFE) preparation was grown in yeast extract-peptone-dextrose (YPD) medium at 29°C for 6 h to a final optical density (OD) of 1.5, followed by 30 min at 37°C before harvesting, and was prepared as described previously (66). The reaction mixture (24l) con-tained 2l of whole CFE; 0.5g of DI-72 (⫹)repRNA transcript; 700 ng of purified MBP-p33; 1.4g of purified MBP-p92pol(both recombinant
proteins were purified fromE. coli); 30 mM HEPES-KOH (pH 7.4); 150 mM potassium acetate; 5 mM magnesium acetate; 0.13 M sorbitol; 0.4l of actinomycin D (5 mg/ml); 2l of 150 mM creatine phosphate; 0.2l of 10 mg/ml creatine kinase; 0.2l of RNase inhibitor; 0.2l of 1 M dithio-threitol (DTT); 2l of 10 mM ATP, CTP, and GTP; 0.25 mM UTP; and 0.1l of [32P]UTP (56,66). In addition to the recombinant viral
replica-tion proteins MBP-p33 and MBP-p92pol, different amounts (16, 32, and
64mol) of the recombinant protein GST, Sec17, Pex5, GST-Sti1, or GST-Cns1 were also included in the assay mixture. The CFE assay was performed at 25°C for 3 h and terminated by adding 100l stop buffer (1% SDS and 0.05 M EDTA [pH 8.0]) to the mixture, followed by extrac-tion with 100l phenol-chloroform, isopropanol-ammonium acetate precipitation overnight at⫺20°C, and a washing step with 70% ethanol (56, 66). The newly synthesized 32P-labeled repRNA products were
treated at 85°C for 5 min and analyzed by using electrophoresis in a 5% polyacrylamide gel containing 0.5⫻Tris-borate-EDTA (TBE) buffer with 8 M urea. Signals were detected by using a Typhoon 9400 imaging scanner (Amersham) and quantified by using imageQuant software. The CFE as-say without MBP-p92polwas used as a negative control. To detect
on November 7, 2019 by guest
http://jvi.asm.org/
stranded repRNA/double-stranded repRNA, the newly synthesized32
P-labeled RNA products without heat treatment were directly separated by electrophoresis (56,66).
Replicase purification from yeast andin vitroRdRp assay.Yeast strains were cotransformed with plasmids pGAD-ADH-HFp92, pYC-GAL-DI72, and pHis-GBK-ADH-HFp33 expressing His6- and
Flag-tagged tombusvirus p33 and p92. The procedure was described in detail previously (56). Briefly, initially, we grew yeast transformants in 10 ml of Ura⫺/Leu⫺/His⫺selective medium overnight at 30°C, and the cultures were then transferred into fresh 200 ml Ura⫺/Leu⫺/His⫺medium to an OD at 600 nm (OD600) of 0.2 and grown at 23°C until the OD600reached
1.5. Copurification was done according to a previously described proce-dure (35), with the following modifications. Yeast cells (2 g) were resus-pended and homogenized in TG buffer (50 mM Tris-HCl [pH 7.5], 10% glycerol, 15 mM MgCl2, 10 mM KCl, 0.5 M NaCl, 0.5% Triton, and 1%
[vol/vol] Ypic) with glass beads by using a FastPrep homogenizer (MP Biomedicals). The yeast cell lysate was cleared by centrifugation at 500⫻
gfor 5 min at 4°C to remove unbroken cells and debris. The membrane fraction containing the viral replicase complex was collected by centrifu-gation at 35,000⫻gfor 15 min at 4°C and then solubilized in 1 ml TG buffer with a buffer containing 2% Triton–1% (vol/vol) Ypic via gentle rotation for 3 h at 4°C. The solubilized membrane fraction was centri-fuged at 35,000⫻gfor 15 min at 4°C, and the supernatant was incubated with 100l anti-Flag M2-agarose affinity resin (Sigma) preequilibrated with 1 ml TG buffer. After 3 h of gentle rotation at 4°C, the column was washed 5 times with TG buffer containing 0.5% Triton. The resin-bound replicase complex was eluted in 700l elution buffer (50 mM Tris-HCl [pH 7.5], 10% glycerol, 15 mM MgCl2, 10 mM KCl, 0.05 M NaCl, 0.5%
Triton, 1% Ypic, and 0.15 mg/ml Flag peptide [Sigma]) following over-night rotation at 4°C.
Thein vitroRdRp activity assay was performed by using DI-72 minus-strand RNA [(⫺)RNA] templates obtained by T7 transcriptionin vitro
(26,52).
In vitroviral RNA recruitment assay.Thein vitroRNA recruitment reaction was performed according methods described previously (56,66), except that32P-labeled DI72 (⫹)repRNA was used and rCTP, rUTP,32
P-labeled UTP, and actinomycin D were omitted from the reaction mixture. Following a 2-h incubation at 25°C, 1 ml reaction buffer was added to the
in vitroreaction mixture and left on ice for 10 min before being centri-fuged at 35,000⫻ g for 45 min. The membrane-bound 32P-labeled
repRNA was extracted from the pellet by the addition of 0.1 ml stop buffer and 0.1 ml phenol-chloroform, followed by brief vortexing on a high setting and centrifugation at 27,000⫻gfor 5 min. The supernatants from each reaction were precipitated with isopropanol-ammonium acetate overnight at⫺20°C. The RNA samples were analyzed by denaturing PAGE.
Protein purification fromE. coli.pMAL-p33 and pMAL-p92 were transformed separately intoE. colistrain BL21(DE3)CodonPlus. Protein expression and amylose affinity purification were performed as described previously (56,63,66). Protein fractions used for the CFE-based replica-tion assays were 95% pure, as determined by 12% SDS-PAGE.
Northern and Western blot analyses. Total RNA isolation and Northern blot analysis were performed as described previously (27). Pro-tein analysis was performed as described previously, using an anti-His6
antibody as the primary antibody for the detection of His-p33 and zz-Cns1 (26). Detection of Flag-Cns1 was carried out by using primary anti-Flag antibody (56). The secondary antibody for both primary antibodies was alkaline phosphatase-conjugated anti-mouse immunoglobulin G (Sigma).
RESULTS
Identification of host proteins with TPR domains that inhibit
TBSV replication in a cell-free replication assay.
To test if some
TPR-containing yeast proteins can directly affect the activity of
the tombusvirus replicase, we purified GST-tagged Cns1p, Sti1p,
Pex5p, and Sec17p yeast proteins from
E. coli
. Cns1p and Sti1p are
cochaperones of heat shock protein 70 (Hsp70) and Hsp90 (
67
),
and Pex5p is a peroxisomal membrane signal receptor for
perox-isomal matrix proteins (
68
), while Sec17p is a membrane protein
required for vesicular transport between the endoplasmic
reticu-lum (ER) and Golgi membrane (
69
). Although these host proteins
have diverse cellular functions, they share similar TPR-like
se-quences. Therefore, we studied their effects on TBSV replication
based on CFE prepared from wild-type (wt) yeast (
Fig. 1
) (
66
).
The CFE-based TBSV replication assay utilizes TBSV (⫹)repRNA
and purified recombinant p33 and p92
polobtained from
E. coli
to
assemble the viral replicase
in vitro
, followed by a single cycle of
complete TBSV replication, resulting in both minus-stranded and
plus-stranded repRNA progeny (
51
,
66
).
Using the CFE-based TBSV replication assay, we found that
Cns1p was a strong inhibitor of TBSV replication (
Fig. 1A
, lanes
15 to 17), while Sec17p and Sti1p had moderate (
Fig. 1A
, lanes 6 to
8 and 12 to 14) and Pex5p had lesser (
Fig. 1A
, lanes 9 to 11 versus
lanes 3 to 5) inhibitory effects under the same conditions. Due to
the remarkable inhibitory effect of the Cns1p cochaperone on
TBSV replication, we decided to further study the role of Cns1p.
Yeast-based studies reveal that Cns1p cochaperone inhibits
TBSV replication.
To test if Cns1p can also act as an inhibitor
in
vivo
, we overexpressed C-terminally tagged Cns1p in yeast
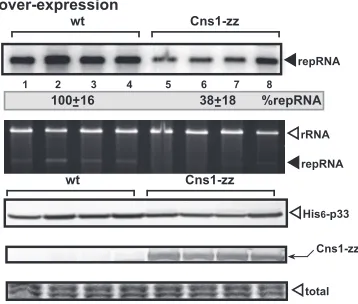
sup-porting TBSV repRNA accumulation. Interestingly, we found that
Cns1p was able to inhibit TBSV RNA accumulation by 60% in
yeast (
Fig. 2
). Since Cns1p is an essential yeast gene, we used a
temperature-sensitive (TS) mutant of Cns1p (
31
,
65
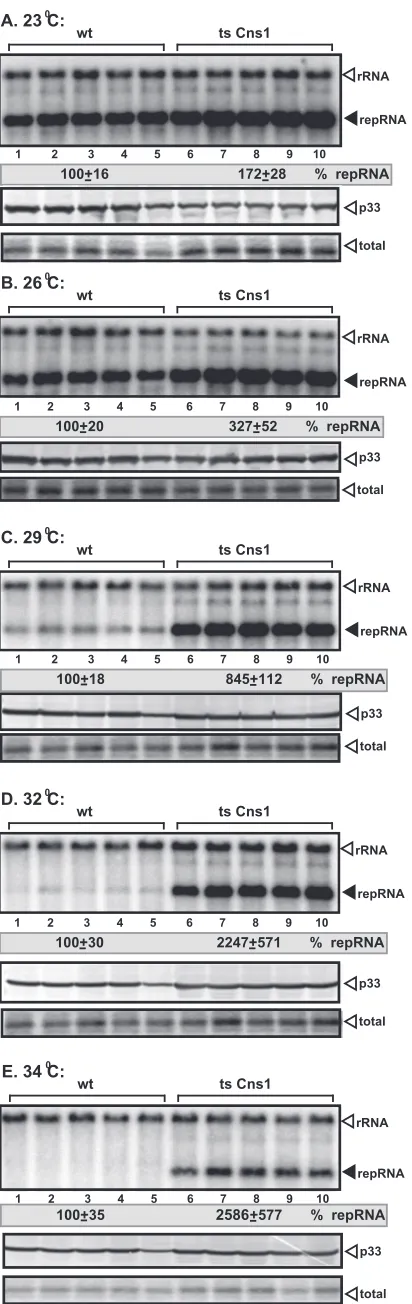
). Yeast cells
expressing TS Cns1p supported TBSV repRNA accumulation at a
⬎20-fold-higher level at semipermissive temperatures than did wt
yeast cells (32°C and 34°C) (
Fig. 3D
and
E
, lanes 6 to 10 versus
lanes 1 to 5), which support TBSV replication poorly at elevated
temperatures. The accumulation of TBSV repRNA was also higher
at lower temperatures (23°C, 26°C, and 29°C) (
Fig. 3A
to
C
) than
in wt yeast. Apparently, partial inactivation of TS Cns1p by
semi-permissive temperatures helps TBSV to partly overcome the
re-striction of replication caused by the higher temperature.
Never-theless, these data with yeast expressing TS Cns1p confirmed that
wt Cns1p is a strong inhibitor of TBSV replication in yeast.
Cns1p interacts with tombusvirus p33 and p92 replication
proteins.
To test if Cns1p can interact with the p33 and p92
rep-lication proteins, we first used the split-ubiquitin-based
two-hy-brid assay (
70
,
71
). We observed strong interactions between
Cns1p and p33 (
Fig. 4A
) and between Cns1p and p92 (
Fig. 4B
).
We confirmed the interaction between Cns1p and the p33
repli-cation protein using a pulldown assay with recombinant Cns1 and
p33 (
Fig. 4C
).
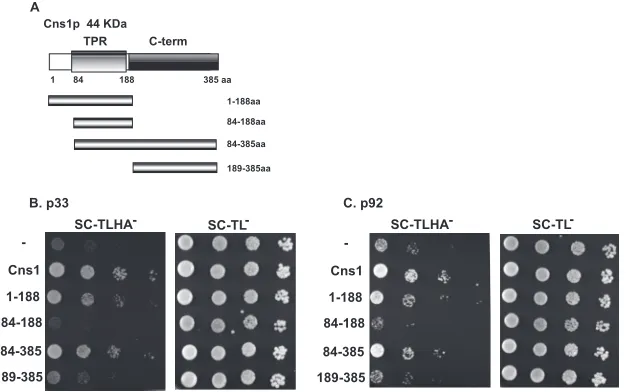
Interestingly, the N-terminal region including the TPR
do-main of Cns1p interacted with p33 although less efficiently than
the full-length protein (amino acids [aa] 1 to 188) (
Fig. 5B
).
Ad-ditional mutants of Cns1p revealed that the C-terminal segment,
when fused with the TPR domain, also contributed to the
interac-tion with p33. However, the C-terminal region (aa 189 to 385) of
Cns1p alone could not interact with p33, suggesting that the TPR
domain is required for the interaction. Interestingly, the minimal
TPR domain (aa 84 to 188) (
Fig. 5A
) of Cns1p did not interact
with either p33 or p92 (
Fig. 5B
and
C
), suggesting that the
inter-action requires sequences outside the minimal TPR domain.
Al-together, the results with p92 were comparable to those with p33
(
Fig. 5C
). Based on these mutants, we propose that the TPR
on November 7, 2019 by guest
http://jvi.asm.org/
main together with flanking N- and C-terminal sequences is
re-quired for binding to p33 and p92 replication proteins.
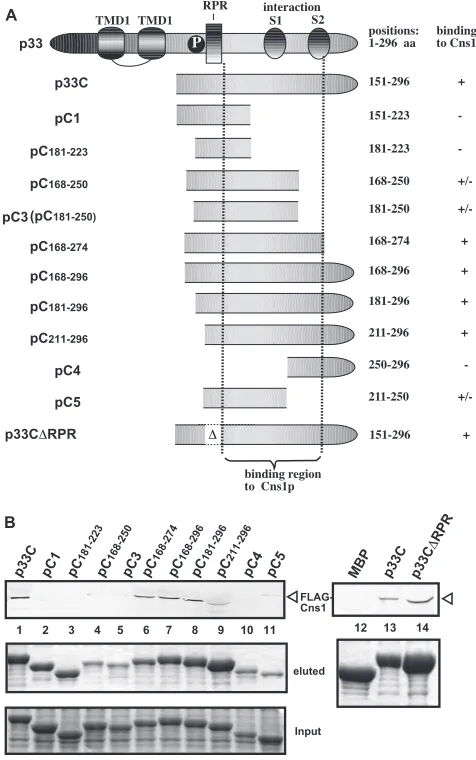
To map the Cns1p binding site in the overlapping TBSV
rep-lication proteins, we used pulldown experiments with
immobi-lized MBP-p33 and its truncation derivatives (
Fig. 6A
) from yeast
extracts containing Flag-tagged Cns1p. These experiments
re-vealed that Cns1p binds to the C-terminal region containing the
p33-p33/p92 interaction sequences. Deletion of the
arginine-pro-line-rich RNA binding domain (RPR) motif, involved in RNA
binding, did not inhibit p33 binding to Cns1p (p33C
⌬
RPR) (
Fig.
6B
, lane 14). The RPR motif in the p33 and p92 replication
pro-teins is required for specific viral RNA recruitment and replicase
assembly, and it also binds to Cpr7p (
56
,
66
,
72
,
73
). Thus, there is
a difference between Cns1p and Cpr7p in binding to p33 and p92,
suggesting that the mechanism of inhibition by Cns1p could be
different from that of Cpr7p in tombusvirus replication.
Cns1p blocks the assembly of the tombusvirus replicase
in
vitro
.
To study which step(s) of the replication process is inhibited
by Cns1p, we first tested plus- and minus-strand synthesis in the
CFE-based assay. Addition of recombinant Cns1p decreased the
production of both double-stranded RNA (dsRNA), which
repre-sents the product of minus-strand RNA synthesis, and
single-stranded RNA (ssRNA), which exclusively consists of new
(⫹)RNAs (
Fig. 7
, lanes 8 to 10 versus lanes 5 to 7) (
51
,
66
). Based
on these data, we suggest that plus- and minus-strand synthesis is
reduced by Cns1p, possibly through inhibition of the replicase
assembly step.
To test if the tombusvirus replicase assembly is affected by
Cns1p
in vitro
, we used a two-step CFE-based assay (
Fig. 8A
) (
66
),
which is based on the assembly of the tombusvirus replicase from
the recombinant replication proteins and host proteins (present
in the CFE) in the presence of ATP/GTP in the first step. This is
followed by a second step where the soluble fraction of CFE and
the unbound viral replication proteins are removed to prevent
additional assembly of new replicase complexes. Moreover, ATP/
CTP/GTP and [
32P]UTP were added to facilitate RNA synthesis
(
66
).
When purified recombinant Cns1p was added to the assay
mixture during the first step,
in vitro
viral RNA synthesis was
strongly inhibited (
Fig. 8B
, lanes 8 to 10 versus lanes 5 to 7). On the
contrary, the addition of Cns1p during the second step of the CFE
assay did not inhibit viral RNA synthesis
in vitro
(
Fig. 8C
, lanes 8
FIG 1Yeast CFE-based TBSV replication assay to identify TPR domain-containing yeast proteins inhibiting TBSV replicationin vitro. (A) Denaturing PAGE analysis of the32P-labeled TBSV repRNA products obtained in the TBSV replication assay. Purified recombinant p33 and p92polreplication proteins of TBSV and in vitro-transcribed TBSV DI-72 (⫹)repRNA as well as the indicated yeast proteins affinity purified as GST-tagged proteins (16, 32, and 64mol) fromE. coli
were added to the whole-cell extract prepared from the BY4741 yeast strain. Each experiment was repeated three times. (B) Coomassie blue-stained SDS-PAGE gel of various recombinant proteins after affinity purification. MW, molecular weight (in thousands).
FIG 2Overexpression of Cns1p inhibits TBSV repRNA accumulation in wt yeast. (Top) Northern blot analysis of TBSV repRNA accumulation in wt yeast and yeast overproducing zz-tagged Cns1p. (Middle) Ethidium bromide stain-ing of an agarose gel demonstratstain-ing comparable levels of rRNA loadstain-ing. (Bot-tom) Detection of overproduced Cns1p in yeast. His6-p33 and zz-tagged (and
also His6-tagged) Cns1p were detected by Western blotting, while the total
protein level in each sample was analyzed by SDS-PAGE and Coomassie blue staining.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.136.452.70.294.2] [image:4.585.74.253.497.649.2]to 10 versus lanes 5 to 7). These experiments demonstrated that
Cns1p mostly inhibits tombusvirus replicase assembly, while RNA
synthesis was not affected when Cns1p was added to the
preas-sembled replicase. This could be due to an inaccessibility of the
preassembled membrane-bound tombusvirus replicase to Cns1p.
To test directly if Cns1p could inhibit the viral RNA synthesis
step, we added purified recombinant Cns1p to an affinity-purified
tombusvirus replicase preparation derived from a solubilized
yeast membrane preparation (
26
,
52
). The purified replicase
de-pends on the added RNA for activity (
Fig. 9A
, lane 1 versus lane 2).
Unlike GST, the addition of Cns1p reduced cRNA synthesis by the
tombusvirus replicase by 40 to 60% (
Fig. 9A
, lanes 8 to 10 versus
lanes 5 to 7), demonstrating that Cns1p could inhibit viral RNA
synthesis if it has access to the viral replicase.
To study if Cns1p inhibits specific viral RNA recruitment by
p33, we added purified recombinant Cns1p to the RNA
recruit-ment assay mixture containing the CFE. Collection of the
mem-brane fraction of the CFE with the bound p33 showed that viral
repRNA recruitment by p33 was comparable in the presence and
in the absence of Cns1p (
Fig. 9B
). Therefore, we suggest that
Cns1p is unlikely to affect the recruitment of the viral RNA to the
membrane-bound replicase.
Cns1p interacts with Nodamura virus and other viral
repli-cation proteins.
To test if Cns1p could interact with replication
proteins of other related and unrelated viruses, we used the
split-ubiquitin assay with the TBSV p33-related turnip crinkle virus
(TCV) p28 and red clover necrotic mosaic virus (RCNMV) p27
replication proteins and the unrelated Nodamura virus (NoV)
protein A and tobacco mosaic virus (TMV) 130K replication
pro-teins. Interestingly, all these replication proteins interacted with
Cns1p in the split-ubiquitin assay (
Fig. 10A
to
D
).
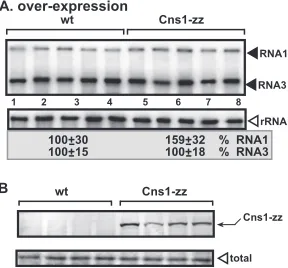
To examine if Cns1p affects the replication of NoV RNA1,
which can also replicate in yeast (
45
,
61
), we overexpressed
zz-tagged Cns1p in yeast. Replication of RNA1 or subgenomic RNA3
made from RNA1 during replication was not affected by
overex-pression of Cns1p (
Fig. 11
, lanes 6 to 10). Thus, NoV replication is
not regulated by Cns1p at a detectable level in yeast.
DISCUSSION
Identification of novel host factors affecting TBSV replication
via a conserved domain search.
Although we have performed
several genome-wide screens to identify host factors affecting
TBSV replication in yeast and performed global proteomics
ap-proaches to find host proteins interacting with viral components
(
15
,
17
,
28–31
,
34–37
), we think that the
⬃
500 host proteins
al-ready identified do not yet represent the complete list of factors.
An alternative approach to find novel host factors is based on
similarity searches for host proteins based on previously identified
2586
FIG 3Stimulatory effect of the TS mutant of Cns1p on TBSV replication in yeast. Shown is the accumulation of TBSV repRNA in the TS Cns1 or wt yeast strain at permissive (23°C and 26°C) (A and B) or semipermissive (29°C, 32°C, and 34°C) (C to E) temperatures. We launched TBSV repRNA replication by expressing His6-p33 and His6-p92 from theCUP1promoter (copper
induc-ible) and DI-72 (⫹)repRNA from the galactose-inducibleGAL1promoter in the TS Cns1 and parental (BY4741) yeast strains. Top panels show Northern blot analysis used to detect DI-72 (⫹)repRNA accumulation. Note that the data were normalized based on 18S rRNA. Each experiment was repeated three times. Middle panels show Western blot analysis of p33 accumulation by anti-His antibody. Bottom panels show Coomassie blue-stained SDS-PAGE gels for total protein levels in the samples.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.62.268.63.715.2]host factors using either entire protein sequences or conserved
functional domains. In this paper, we used the TPR domain-based
approach to further test the function of this domain in TBSV
replication. The TPR domain of Cpr7p cyclophilin was identified
previously to interact with the p33 and p92 replication proteins
and to restrict TBSV replication (
56
). Testing of four TPR
do-main-containing cytosolic yeast proteins revealed that three of
them acted as inhibitors of TBSV replication in the CFE assay (
Fig.
1
). Thus, this approach seems to be rather effective in the
identi-fication of new host restriction factors. Indeed, we also used a
similar approach with WW motif-containing host proteins, such
as the Rsp5p E3 ubiquitin ligase, to identify additional yeast and
plant proteins carrying WW motifs that function as TBSV
restric-tion factors (
54
,
55
). Another powerful approach is to study all the
members of a whole protein family for which one or more
com-ponents have been identified in high-throughput screens.
Accord-ingly, we used this strategy to test the functional roles of the
mul-timember ESCRT family (
43
) and the large cyclophilin family and
related peptidyl-prolyl
cis-trans
isomerases in TBSV replication
(
34
,
56
). Altogether, the combination of several strategies likely
increases the completeness of the catalog of host factors involved
in TBSV replication.
Cns1p cochaperone restricts TBSV replication through
blocking the assembly of the replicase.
The essential yeast protein
Cns1p is a TPR domain-containing cochaperone that interacts
with the Hsp70 chaperone, although its cellular function is
cur-rently unknown (
57
). We found that Cns1p strongly interacts
with the TBSV p33 and p92 replication proteins via its extended
TPR domain (
Fig. 4
and
5
). The interaction between Cns1p and
the viral replication proteins seems to be blocking the assembly of
FIG 4Interaction between Cns1p and the p33 and p92 replication proteins. (A and B) A split-ubiquitin membrane yeast two-hybrid assay was used to test binding between p33 or p92 and full-length yeast Cns1p. The bait protein p33 or p92 was coexpressed with the prey protein Cns1p in yeast. SSA1 (Hsp70 chaperone) and the empty prey vector (NubG) were used as positive and negative controls, respectively. Yeast cells were grown on a TLHA⫺plate to test protein interactions, and a control plate showing growth of the yeast transformants on nonselective TL⫺medium was included. SC, synthetic complete medium. (C) Affinity binding (pulldown) assay to detect interactions between Flag-Cns1p and the MBP-tagged viral p33 protein (the C-terminal portion). The MBP-tagged viral protein or the MBP control produced inE. coliwas immobilized on amylose affinity columns. Flag-Cns1p expressed in BY4741 yeast cells was then passed through the amylose affinity columns with immobilized MBP-tagged proteins. The affinity-bound proteins were eluted from the columns with maltose. The eluted proteins were analyzed by Western blotting with anti-Flag antibody to detect the amount of Flag-Cns1p specifically bound to MBP-tagged viral protein.
FIG 5Sequence of the interaction between Cns1p and the p33 and p92 replication proteins. (A) Schematic representation of the yeast Cns1p cochaperone and its derivatives used in the interaction assay. The various domains include the TPR domain (consisting of three TPR motifs) involved in interactions with Hsp70s and Hsp90s and the C-terminal domain of unknown function. (B and C) A membrane yeast two-hybrid assay was used to define the interaction of the domains in yeast Cns1p with full-length TBSV p33 or p92. See the legend ofFig. 4for further details.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.126.479.67.202.2] [image:6.585.139.454.488.684.2]the tombusvirus replicase, based on a CFE assay (
Fig. 8
). Cns1p
also inhibited viral RNA synthesis with the purified replicase
in
vitro
(
Fig. 9A
). Therefore, the decreased level of replicase assembly
and the inhibition of RNA synthesis in the presence of Cns1p are
the likely reasons why (i) Cns1p inhibits both (
⫺
)RNA and
(⫹)RNA synthesis
in vitro
(
Fig. 7
), (ii) overexpression of Cns1p
leads to decreased TBSV repRNA accumulation in yeast (
Fig. 2
),
and (iii) expression of TS Cns1p (as the only copy of Cns1p in
yeast) increased TBSV repRNA accumulation at semipermissive
temperatures (
Fig. 3
).
Cns1p does not seem to affect the recruitment of the viral RNA
to membranes performed by the p33 replication protein (
Fig. 9B
).
Also, Cns1p could not inhibit RNA synthesis with the
preas-sembled tombusvirus replicase (
Fig. 8C
), although it did inhibit
RNA synthesis by the purified tombusvirus replicase
in vitro
(
Fig.
9A
). These data suggest that Cns1p was incapable of entering the
fully assembled membrane-bound replicase complex once the
VRC was formed. In spite of these limitations, Cns1p seems to be
a strong inhibitor of TBSV replication in yeast.
There are two possible scenarios for Cns1p-mediated
inhibi-tion of VRC assembly. The first one is based on direct inhibiinhibi-tion of
p33-p33 and p33-p92
polinteractions via binding of Cns1p to the
p33-p33/p92 interaction domain present in both the p33 and
p92
polreplication proteins. The interactions between the two viral
replication proteins are required for VRC assembly, as shown
pre-viously using p33 and p92
polmutants in CFE-based replication
and VRC assembly assays (
49
,
74
). Accordingly, the observed
in-hibition of the
in vitro
RdRp activity of the purified replicase by the
addition of Cns1p (
Fig. 9A
) could be due to direct binding of
Cns1p to p92
polleading to reduced RdRp activity. Indeed, we have
shown previously that p92
polrequires the p33-p33/p92
interac-tion domain for RdRp activity
in vitro
(
75
). In the second scenario,
it is also possible that Cns1p affects the activity of the subverted
cellular Hsp70 molecules, which are residents of the VRCs. Cns1p
has been shown to stimulate the ATPase activity of the yeast Ssa1
and Ssa2 Hsp70s (
57
), which in turn could interfere with the
func-tion of Hsp70s to support VRC assembly. The Ssa1/Ssa2 Hsp70s
FIG 6Sequence within the TBSV p33 protein needed for binding to Cns1pin vitro. (A) Schematic representation of TBSV p33 and its truncated derivatives used in the binding assay. TMD, transmembrane domain; RPR, arginine-pro-line-rich RNA binding domain (RPR); P, phosphorylated serine and threo-nine; S1 and S2, subdomains involved in the p33-p33/p92 interaction. (B) Affinity binding (pulldown) assay to detect interactions between Flag-Cns1p and the MBP-tagged viral p33 protein derivatives. The MBP-tagged viral pro-teins produced inE. coliwere immobilized on amylose affinity columns. Flag-Cns1p expressed in BY4741 yeast cells was then passed through the amylose affinity columns with immobilized MBP-tagged proteins. The affinity-bound proteins were eluted from the columns with maltose (shown in the middle image). The top image shows the eluted proteins analyzed by Western blotting with anti-Flag antibody to detect the amount of Flag-Cns1p specifically bound to MBP-tagged viral proteins. Note that the binding of three p33-derived pro-teins to Cns1p is indicated by “⫹/⫺,” possibly due to the low level of recovery of these proteins in the eluted fraction. The bottom image shows SDS-PAGE analysis of the totalE. coliproteins prior to immobilization of MBP-tagged proteins on the amylose affinity columns.
FIG 7Cns1p inhibits both minus- and plus-strand synthesis in the CFE-based TBSV replication assay. Purified recombinant p33 and p92polreplication
pro-teins of TBSV andin vitro-transcribed TBSV DI-72 (⫹)repRNA were added to the whole-cell extract prepared from yeast. Nondenaturing PAGE analysis of the32P-labeled TBSV repRNA products obtained in the TBSV replication
as-say based on the CFE is shown. The ssRNA product, representing the newly made (⫹)RNA, and the dsRNA product, representing the replication interme-diate with annealed minus and plus strands, are depicted with arrowheads. The dsRNA product made without added recombinant GST-Cns1p (lane 1) was chosen as 100%. Each experiment was repeated three times.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.312.526.66.282.2] [image:7.585.42.280.67.447.2]are known to assist in the intracellular localization and proper
membrane insertion of both the p33 and p92
polreplication
pro-teins. These chaperones are also required for the activation of the
RdRp function of p92
poland the assembly of the VRCs (
9
,
37
,
41
,
42
,
66
). By affecting the activity of Hsp70s, Cns1p could inhibit
any of the above-described steps, resulting in inhibition of VRC
assembly.
We do not yet know if Cns1p could inhibit the replication of
other viruses. Based on interaction, it seems that several viral
rep-lication proteins, both related and unrelated to the tombusvirus
replication proteins, interact strongly with Cns1p in yeast cells.
This finding is similar to those reported previously for Cpr7p
cy-clophilin (
56
). However, while Cpr7p inhibited both TBSV and
nodavirus replication (
56
), Cns1p seems to be a specific restriction
factor for TBSV, and it did not influence NoV RNA accumulation
in yeast (
Fig. 11
). We do not know the reasons for this difference in
the ability of Cns1p to act as a viral restriction factor. These viruses
replicate in different microenvironments, as TBSV recruits
perox-isomal and, occasionally, ER membranes (
47
,
49
,
76
), while
noda-FIG 9Cns1p inhibits cRNA synthesis by the affinity-purified tombusvirus replicase. (A) Representative denaturing gel of32P-labeled RNA products
syn-thesized by the purified tombusvirus replicasein vitroin the presence of 16, 32, and 64mol purified recombinant Cns1p or GST. Thein vitroassay mixtures were programmed with DI-72 (⫺)repRNA, and they also contained rATP/ rCTP/rGTP and [32P]rUTP. All the components were added at the same time.
The level of cRNA synthesis producing the full-length cRNA product (made via initiation from the 3=-terminal promoter) is marked as “de novorepRNA.” The quantification was based on this product, which was chosen as 100% for the control sample (lane 2). Note that this replicase preparation also synthe-sizes internal initiation products (“ii”) and 3=-terminal extension products (“3=TEX”). Each experiment was repeated three times. Note that lanes 3 and 4 contain the purified GST and GST-Cns1p proteins, respectively, in combina-tion with the template RNA to exclude that contaminating terminal trans-ferase activity is present in these purified preparations. (B) Cns1p does not affect the recruitment of the viral (⫹)repRNA to membranesin vitro.32
P-labeled TBSV (⫹)repRNA transcripts were added to the CFE in the presence of the p33 and p92 replication proteins, unlabeled ribonucleotides, and purified GST or GST-Cns1p, as shown. The denaturing PAGE gel shows the repRNA associated with the membranes from the CFE. Note that the (⫹)repRNA binds inefficiently to the membrane in the absence of p33/p92, likely due to an association with a membrane-bound cellular protein(s).
FIG 8An inhibitory role for Cns1p in the assembly of the TBSV VRC is supported by a stepwise CFE-based TBSV replication assay. (A) Scheme of the CFE-based TBSV replicase assembly assay. In step 1, purified recombinant p33 and p92polreplication proteins of TBSV andin vitro-transcribed TBSV DI-72
(⫹)repRNA were added to CFE prepared from the BY4741 yeast strain. The assay mixture either contained or lacked purified recombinant Cns1p (16, 32, and 64mol) or GST during step 1. Note that the assay was performed in the presence of rATP/rGTP to facilitate TBSV VRC assembly but prevent RNA synthesis in step 1. Step 2 was performed after step 1 and centrifugation to collect the membrane fraction of the CFE, in the presence of rATP/rCTP/rGTP and32P-rUTP to allow TBSV RNA replication. (B) Denaturing PAGE analysis
of the32P-labeled TBSV repRNA products obtained in the CFE-based assays
when Cns1p was added during the first step. See the legend ofFig. 1for further details. (C) Denaturing PAGE analysis of the32P-labeled TBSV repRNA
prod-ucts obtained in the CFE-based assays when Cns1p was added at the second step. Three repeats of each experiment were performed.
FIG 10Interactions between Cns1p and various viral replication proteins. A split-ubiquitin membrane yeast two-hybrid assay was used to test binding between full-length yeast Cns1p and NoV protein A (protA), (A) TCV p28 (B), RCNMV p27 (C), and TMV 130K (D) replication proteins. The bait viral replication pro-teins were coexpressed with the prey Cns1p protein in yeast. SSA1 (Hsp70 chap-erone) and the empty prey vector (NubG) were used as positive and negative controls, respectively. See the legend ofFig. 4for further details.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.335.508.71.255.2] [image:8.585.62.265.80.387.2] [image:8.585.353.490.543.657.2]viruses replicate on mitochondrial membranes (
77
). Thus, it is
possible that the accessibility of Cns1p to these viruses may be
different and may affect the inhibitory function of Cns1p. It is also
possible that binding of Cns1p to the region of the replication
proteins might be critical. Indeed, we have shown that the
differ-ence in the abilities of Cpr7p and Cpr6p cyclophilins to inhibit
TBSV replication was likely due to differences in binding to target
sequences in the p33 and p92 replication proteins (
56
).
Interest-ingly, the inhibitory Cpr7p cyclophilin bound to the RNA binding
region of p33/p92 (
56
), while Cns1p bound to the C-terminal
p33-p33/p92 interaction domain in p33/p92 (
Fig. 6
). Thus, how
these proteins affect the functions of viral replication proteins to
restrict viral replication may be different.
Cns1p cochaperone plays a role similar to that of the Cpr7p
Cyp40-like cyclophilin in TBSV replication.
Comparison of the
effects of TPR-containing Cpr7p and Cns1p on TBSV replication
shows many similarities between the viral restriction functions of
these cellular proteins. In addition to the strong inhibitory
func-tions by both proteins on TBSV replication in yeast (
56
), Cns1p
and Cpr7p were shown to block the assembly of the tombusvirus
VRC and inhibit RNA synthesis by the viral replicase
in vitro
. Also,
similar to Cns1p, the TPR domain of Cpr7p interacts with p33 and
p92
pol(
56
). Thus, the emerging picture is that Cpr7p and Cns1p
have similar inhibitory functions during TBSV replication,
al-though some of the details of the mechanisms are different. For
example, Cpr7p efficiently inhibits the recruitment of the viral
RNA for replication (
56
), while Cns1p seems ineffective in this
step. Also, the two proteins recognize different domains in the p33
and p92
polreplication proteins. Based on the binding of Cpr7p to
the RNA binding domains in p33 and p92
pol, we proposed that
Cpr7p inhibits TBSV replication by preventing the proper
associ-ation between the viral (
⫹
)RNA and the replication proteins. This
leads to the inhibition of viral (⫹)RNA recruitment for
replica-tion and also blocking of VRC assembly, which utilizes the viral
(⫹)RNA as a platform in the assembly process (
48
,
52
,
66
,
72
). On
the other hand, Cns1p seems to inhibit VRC assembly via
inhib-iting the interaction between p33 and p92
polmolecules, which is
essential for assembly (
74
). It is also possible that Cns1p might
inhibit the essential function of coopted Hsp70 during VRC
as-sembly, as discussed above. It is also possible that Cns1p and
Cpr7p viral restriction factors could work together to inhibit
TBSV replication, since their interaction in yeast has been
dem-onstrated (
59
,
78
).
Overall, our observations with Cns1p, Cpr7p, and two
addi-tional TPR domain proteins, namely, Sec17p and Sti1p (
Fig. 1
),
indicate that potentially more members of the large
TPR-contain-ing protein family could act as virus restriction factors. However,
based on yeast Cpr6p (
56
), we already know that not all
TPR-containing proteins are inhibitory factors of TBSV replication in
spite of their abilities to interact with the p33 and p92 replication
proteins. Since a large number of TPR-containing proteins are
predicted to exist in all eukaryotes, including
⬃
25 in
S. cerevisiae
,
⬃80 in
Caenorhabditis elegans
,
⬃200 in humans, and
⬃235 in
Arabidopsis
(36 of which are predicted to bind to Hsp90/Hsp70
chaperones) (
79
,
80
), we do not yet know which and how many
members of the TPR domain proteins are actual restriction factors
during TBSV replication.
ACKNOWLEDGMENTS
We thank Daniel Barajas, Judit Pogany, and Reddisiva Prasanth for criti-cal reading of the manuscript and for very helpful suggestions.
This work was supported by the NIAID (grant 5R21AI096323-02).
REFERENCES
1.Nagy PD.2008. Yeast as a model host to explore plant virus-host inter-actions. Annu. Rev. Phytopathol.46:217–242.
2.Nagy PD, Pogany J.2008. Multiple roles of viral replication proteins in plant RNA virus replication. Methods Mol. Biol.451:55– 68.
3.Nagy PD, Wang RY, Pogany J, Hafren A, Makinen K.2011. Emerging picture of host chaperone and cyclophilin roles in RNA virus replication. Virology411:374 –382.
4.Li Z, Nagy PD.2011. Diverse roles of host RNA binding proteins in RNA virus replication. RNA Biol.8:305–315.
5.den Boon JA, Diaz A, Ahlquist P.2010. Cytoplasmic viral replication complexes. Cell Host Microbe8:77– 85.
6.Salonen A, Ahola T, Kaariainen L.2005. Viral RNA replication in asso-ciation with cellular membranes. Curr. Top. Microbiol. Immunol.285: 139 –173.
7.Miller S, Krijnse-Locker J.2008. Modification of intracellular membrane structures for virus replication. Nat. Rev. Microbiol.6:363–374. 8.Fernandez-Garcia MD, Mazzon M, Jacobs M, Amara A.2009.
Patho-genesis of flavivirus infections: using and abusing the host cell. Cell Host Microbe5:318 –328.
9.Nagy PD, Pogany J.2011. The dependence of viral RNA replication on co-opted host factors. Nat. Rev. Microbiol.10:137–149.
10. Fischl W, Bartenschlager R.2011. Exploitation of cellular pathways by dengue virus. Curr. Opin. Microbiol.14:470 – 475.
11. Diamond MS, Gale M, Jr.2012. Cell-intrinsic innate immune control of West Nile virus infection. Trends Immunol.33:522–530.
12. Aoshi T, Koyama S, Kobiyama K, Akira S, Ishii KJ.2011. Innate and adaptive immune responses to viral infection and vaccination. Curr. Opin. Virol.1:226 –232.
13. Jensen S, Thomsen AR.2012. Sensing of RNA viruses: a review of innate immune receptors involved in recognizing RNA virus invasion. J. Virol. 86:2900 –2910.
14. Ding SW.2010. RNA-based antiviral immunity. Nat. Rev. Immunol. 10:632– 644.
15. Panavas T, Serviene E, Brasher J, Nagy PD.2005. Yeast genome-wide screen reveals dissimilar sets of host genes affecting replication of RNA viruses. Proc. Natl. Acad. Sci. U. S. A.102:7326 –7331.
16. Cherry S, Doukas T, Armknecht S, Whelan S, Wang H, Sarnow P, Perrimon N.2005. Genome-wide RNAi screen reveals a specific sensitiv-ity of IRES-containing RNA viruses to host translation inhibition. Genes Dev.19:445– 452.
17. Jiang Y, Serviene E, Gal J, Panavas T, Nagy PD.2006. Identification of essential host factors affecting tombusvirus RNA replication based on the yeast Tet promoters Hughes Collection. J. Virol.80:7394 –7404. FIG 11Unaltered level of NoV RNA accumulation in yeast overexpressing
Cns1p. To launch NoV RNA1 replication, we expressed NoV RNA1 from the copper-inducibleCUP1promoter in the parental BY4741 yeast strain. The yeast cells were cultured for 48 h at 29°C. (A) Northern blot analysis was used to detect NoV RNA1 and subgenomic RNA3 accumulation. The accumulation level of NoV RNAs was normalized based on the 18S rRNA level. Each exper-iment was repeated. (B) zz-tagged (and also His6-tagged) Cns1p was detected
by Western blotting.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.92.236.68.202.2]18. Kushner DB, Lindenbach BD, Grdzelishvili VZ, Noueiry AO, Paul SM, Ahlquist P.2003. Systematic, genome-wide identification of host genes affecting replication of a positive-strand RNA virus. Proc. Natl. Acad. Sci. U. S. A.100:15764 –15769.
19. Krishnan MN, Ng A, Sukumaran B, Gilfoy FD, Uchil PD, Sultana H, Brass AL, Adametz R, Tsui M, Qian F, Montgomery RR, Lev S, Mason PW, Koski RA, Elledge SJ, Xavier RJ, Agaisse H, Fikrig E.2008. RNA interference screen for human genes associated with West Nile virus in-fection. Nature455:242–245.
20. Li Q, Brass AL, Ng A, Hu Z, Xavier RJ, Liang TJ, Elledge SJ.2009. A genome-wide genetic screen for host factors required for hepatitis C virus propagation. Proc. Natl. Acad. Sci. U. S. A.106:16410 –16415.
21. Randall G, Panis M, Cooper JD, Tellinghuisen TL, Sukhodolets KE, Pfeffer S, Landthaler M, Landgraf P, Kan S, Lindenbach BD, Chien M, Weir DB, Russo JJ, Ju J, Brownstein MJ, Sheridan R, Sander C, Zavolan M, Tuschl T, Rice CM.2007. Cellular cofactors affecting hepatitis C virus infection and replication. Proc. Natl. Acad. Sci. U. S. A.104:12884 –12889. 22. Sessions OM, Barrows NJ, Souza-Neto JA, Robinson TJ, Hershey CL, Rodgers MA, Ramirez JL, Dimopoulos G, Yang PL, Pearson JL, Garcia-Blanco MA.2009. Discovery of insect and human dengue virus host factors. Nature458:1047–1050.
23. Tai AW, Benita Y, Peng LF, Kim SS, Sakamoto N, Xavier RJ, Chung RT. 2009. A functional genomic screen identifies cellular cofactors of hepatitis C virus replication. Cell Host Microbe5:298 –307.
24. Nagy PD, Pogany J.2006. Yeast as a model host to dissect functions of viral and host factors in tombusvirus replication. Virology344:211–220. 25. White KA, Nagy PD.2004. Advances in the molecular biology of
tom-busviruses: gene expression, genome replication, and recombination. Prog. Nucleic Acid Res. Mol. Biol.78:187–226.
26. Panaviene Z, Panavas T, Serva S, Nagy PD.2004. Purification of the cucumber necrosis virus replicase from yeast cells: role of coexpressed viral RNA in stimulation of replicase activity. J. Virol.78:8254 – 8263. 27. Panavas T, Nagy PD.2003. Yeast as a model host to study replication and
recombination of defective interfering RNA of Tomato bushy stunt virus. Virology314:315–325.
28. Serviene E, Shapka N, Cheng CP, Panavas T, Phuangrat B, Baker J, Nagy PD.2005. Genome-wide screen identifies host genes affecting viral RNA recombination. Proc. Natl. Acad. Sci. U. S. A.102:10545–10550. 29. Serviene E, Jiang Y, Cheng CP, Baker J, Nagy PD.2006. Screening of the
yeast yTHC collection identifies essential host factors affecting tombusvi-rus RNA recombination. J. Virol.80:1231–1241.
30. Shah Nawaz-Ul-Rehman M, Martinez-Ochoa N, Pascal H, Sasvari Z, Herbst C, Xu K, Baker J, Sharma M, Herbst A, Nagy PD. 2012. Proteome-wide overexpression of host proteins for identification of fac-tors affecting tombusvirus RNA replication: an inhibitory role of protein kinase C. J. Virol.86:9384 –9395.
31. Shah Nawaz-Ul-Rehman M, Reddisiva Prasanth K, Baker J, Nagy PD. 2013. Yeast screens for host factors in positive-strand RNA virus replication based on a library of temperature-sensitive mutants. Methods59:207–216. 32. Nagy PD.2011. The roles of host factors in tombusvirus RNA
recombi-nation. Adv. Virus Res.81:63– 84.
33. Nagy PD, Pogany J.2010. Global genomics and proteomics approaches to identify host factors as targets to induce resistance against Tomato bushy stunt virus. Adv. Virus Res.76:123–177.
34. Mendu V, Chiu M, Barajas D, Li Z, Nagy PD.2010. Cpr1 cyclophilin and Ess1 parvulin prolyl isomerases interact with the tombusvirus repli-cation protein and inhibit viral replirepli-cation in yeast model host. Virology 406:342–351.
35. Li Z, Pogany J, Panavas T, Xu K, Esposito AM, Kinzy TG, Nagy PD. 2009. Translation elongation factor 1A is a component of the tombusvirus replicase complex and affects the stability of the p33 replication co-factor. Virology385:245–260.
36. Li Z, Barajas D, Panavas T, Herbst DA, Nagy PD.2008. Cdc34p ubiquitin-conjugating enzyme is a component of the tombusvirus replicase complex and ubiquitinates p33 replication protein. J. Virol.82:6911– 6926. 37. Serva S, Nagy PD.2006. Proteomics analysis of the tombusvirus replicase:
Hsp70 molecular chaperone is associated with the replicase and enhances viral RNA replication. J. Virol.80:2162–2169.
38. Nagy PD, Barajas D, Pogany J.2012. Host factors with regulatory roles in tombusvirus replication. Curr. Opin. Virol.2:691– 698.
39. Wang RY, Nagy PD.2008. Tomato bushy stunt virus co-opts the RNA-binding function of a host metabolic enzyme for viral genomic RNA syn-thesis. Cell Host Microbe3:178 –187.
40. Li Z, Pogany J, Tupman S, Esposito AM, Kinzy TG, Nagy PD.2010. Translation elongation factor 1A facilitates the assembly of the tombusvi-rus replicase and stimulates minus-strand synthesis. PLoS Pathog. 6:e1001175. doi:10.1371/journal.ppat.1001175.
41. Wang RY, Stork J, Pogany J, Nagy PD.2009. A temperature sensitive mutant of heat shock protein 70 reveals an essential role during the early steps of tombusvirus replication. Virology394:28 –38.
42. Wang RY, Stork J, Nagy PD.2009. A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to in-tracellular membranes. J. Virol.83:3276 –3287.
43. Barajas D, Jiang Y, Nagy PD.2009. A unique role for the host ESCRT proteins in replication of Tomato bushy stunt virus. PLoS Pathog. 5:e1000705. doi:10.1371/journal.ppat.1000705.
44. Huang TS, Nagy PD.2011. Direct inhibition of tombusvirus plus-strand RNA synthesis by a dominant negative mutant of a host metabolic en-zyme, glyceraldehyde-3-phosphate dehydrogenase, in yeast and plants. J. Virol.85:9090 –9102.
45. Kovalev N, Pogany J, Nagy PD. 2012. A co-opted DEAD-box RNA helicase enhances tombusvirus plus-strand synthesis. PLoS Pathog. 8:e1002537. doi:10.1371/journal.ppat.1002537.
46. Sasvari Z, Izotova L, Kinzy TG, Nagy PD.2011. Synergistic roles of eukaryotic translation elongation factors 1Bgamma and 1A in stimulation of tombusvirus minus-strand synthesis. PLoS Pathog.7:e1002438. doi:10 .1371/journal.ppat.1002438.
47. Jonczyk M, Pathak KB, Sharma M, Nagy PD.2007. Exploiting alterna-tive subcellular location for replication: tombusvirus replication switches to the endoplasmic reticulum in the absence of peroxisomes. Virology 362:320 –330.
48. Pogany J, White KA, Nagy PD.2005. Specific binding of tombusvirus replication protein p33 to an internal replication element in the viral RNA is essential for replication. J. Virol.79:4859 – 4869.
49. Panavas T, Hawkins CM, Panaviene Z, Nagy PD.2005. The role of the p33:p33/p92 interaction domain in RNA replication and intracellular lo-calization of p33 and p92 proteins of Cucumber necrosis tombusvirus. Virology338:81–95.
50. Stork J, Kovalev N, Sasvari Z, Nagy PD.2011. RNA chaperone activity of the tombusviral p33 replication protein facilitates initiation of RNA syn-thesis by the viral RdRp in vitro. Virology409:338 –347.
51. Pogany J, Nagy PD.2008. Authentic replication and recombination of Tomato bushy stunt virus RNA in a cell-free extract from yeast. J. Virol. 82:5967–5980.
52. Panaviene Z, Panavas T, Nagy PD.2005. Role of an internal and two 3=-terminal RNA elements in assembly of tombusvirus replicase. J. Virol. 79:10608 –10618.
53. Jiang Y, Li Z, Nagy PD.2010. Nucleolin/Nsr1p binds to the 3= non-coding region of the tombusvirus RNA and inhibits replication. Virol-ogy396:10 –20.
54. Barajas D, Li Z, Nagy PD.2009. The Nedd4-type Rsp5p ubiquitin ligase inhibits tombusvirus replication by regulating degradation of the p92 rep-lication protein and decreasing the activity of the tombusvirus replicase. J. Virol.83:11751–11764.
55. Qin J, Barajas D, Nagy PD.2012. An inhibitory function of WW domain-containing host proteins in RNA virus replication. Virology426:106 –119. 56. Lin JY, Mendu V, Pogany J, Qin J, Nagy PD.2012. The TPR domain in the host Cyp40-like cyclophilin binds to the viral replication protein and inhibits the assembly of the tombusviral replicase. PLoS Pathog. 8:e1002491. doi:10.1371/journal.ppat.1002491.
57. Hainzl O, Wegele H, Richter K, Buchner J.2004. Cns1 is an activator of the Ssa1 ATPase activity. J. Biol. Chem.279:23267–23273.
58. Marsh JA, Kalton HM, Gaber RF.1998. Cns1 is an essential protein associated with the hsp90 chaperone complex in Saccharomyces cerevisiae that can restore cyclophilin 40-dependent functions in cpr7Delta cells. Mol. Cell. Biol.18:7353–7359.
59. Dolinski KJ, Cardenas ME, Heitman J.1998. CNS1 encodes an essential p60/Sti1 homolog in Saccharomyces cerevisiae that suppresses cyclophilin 40 mutations and interacts with Hsp90. Mol. Cell. Biol.18:7344 –7352. 60. Jaag HM, Stork J, Nagy PD.2007. Host transcription factor Rpb11p
affects tombusvirus replication and recombination via regulating the ac-cumulation of viral replication proteins. Virology368:388 – 404. 61. Pogany J, Panavas T, Serviene E, Nawaz-Ul-Rehman MS, Nagy PD.
2010. A high-throughput approach for studying virus replication in yeast. Curr. Protoc. Microbiol. Chapter 16:Unit 16J. doi:10.1002 /9780471729259.mc16j01s19.
on November 7, 2019 by guest
http://jvi.asm.org/
62. Barajas D, Nagy PD.2010. Ubiquitination of tombusvirus p33 replica-tion protein plays a role in virus replicareplica-tion and binding to the host Vps23p ESCRT protein. Virology397:358 –368.
63. Rajendran KS, Nagy PD.2003. Characterization of the RNA-binding domains in the replicase proteins of tomato bushy stunt virus. J. Virol. 77:9244 –9258.
64. Rajendran KS, Nagy PD.2006. Kinetics and functional studies on inter-action between the replicase proteins of Tomato bushy stunt virus: re-quirement of p33:p92 interaction for replicase assembly. Virology345: 270 –279.
65. Li Z, Vizeacoumar FJ, Bahr S, Li J, Warringer J, Vizeacoumar FS, Min R, Vandersluis B, Bellay J, Devit M, Fleming JA, Stephens A, Haase J, Lin ZY, Baryshnikova A, Lu H, Yan Z, Jin K, Barker S, Datti A, Giaever G, Nislow C, Bulawa C, Myers CL, Costanzo M, Gingras AC, Zhang Z, Blomberg A, Bloom K, Andrews B, Boone C.2011. Systematic explora-tion of essential yeast gene funcexplora-tion with temperature-sensitive mutants. Nat. Biotechnol.29:361–367.
66. Pogany J, Stork J, Li Z, Nagy PD.2008. In vitro assembly of the Tomato bushy stunt virus replicase requires the host heat shock protein 70. Proc. Natl. Acad. Sci. U. S. A.105:19956 –19961.
67. Li J, Richter K, Buchner J.2011. Mixed Hsp90-cochaperone complexes are important for the progression of the reaction cycle. Nat. Struct. Mol. Biol.18:61– 66.
68. Brown LA, Baker A.2003. Peroxisome biogenesis and the role of protein import. J. Cell. Mol. Med.7:388 – 400.
69. Bonifacino JS, Glick BS.2004. The mechanisms of vesicle budding and fusion. Cell116:153–166.
70. Iyer K, Burkle L, Auerbach D, Thaminy S, Dinkel M, Engels K, Stagljar I.2005. Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins. Sci. STKE2005:pl3. doi:10.1126/stke.2752005pl3.
71. Kittanakom S, Chuk M, Wong V, Snyder J, Edmonds D, Lydakis A, Zhang Z, Auerbach D, Stagljar I.2009. Analysis of membrane protein
complexes using the split-ubiquitin membrane yeast two-hybrid (MYTH) system. Methods Mol. Biol.548:247–271.
72. Pathak KB, Pogany J, Xu K, White KA, Nagy PD.2012. Defining the roles of cis-acting RNA elements in tombusvirus replicase assembly in vitro. J. Virol.86:156 –171.
73. Pathak KB, Pogany J, Nagy PD.2011. Non-template functions of the viral RNA in plant RNA virus replication. Curr. Opin. Virol.1:332–338. 74. Pathak KB, Jiang Z, Ochanine V, Sharma M, Pogany J, Nagy PD.2013.
Characterization of dominant-negative and temperature-sensitive mu-tants of tombusvirus replication proteins affecting replicase assembly. Vi-rology437:48 – 61.
75. Pogany J, Nagy PD.2012. p33-independent activation of a truncated p92 RNA-dependent RNA polymerase of Tomato bushy stunt virus in yeast cell-free extract. J. Virol.86:12025–12038.
76. McCartney AW, Greenwood JS, Fabian MR, White KA, Mullen RT. 2005. Localization of the tomato bushy stunt virus replication protein p33 reveals a peroxisome-to-endoplasmic reticulum sorting pathway. Plant Cell17:3513–3531.
77. Kopek BG, Perkins G, Miller DJ, Ellisman MH, Ahlquist P. 2007. Three-dimensional analysis of a viral RNA replication complex reveals a virus-induced mini-organelle. PLoS Biol. 5:e220. doi:10.1371/journal .pbio.0050220.
78. Tesic M, Marsh JA, Cullinan SB, Gaber RF.2003. Functional interac-tions between Hsp90 and the co-chaperones Cns1 and Cpr7 in Saccharo-myces cerevisiae. J. Biol. Chem.278:32692–32701.
79. Haslbeck V, Eckl JM, Kaiser CJ, Papsdorf K, Hessling M, Richter K. 2013. Chaperone-interacting TPR proteins in Caenorhabditis elegans. J. Mol. Biol.425:2922–2939.
80. Prasad BD, Goel S, Krishna P.2010. In silico identification of carboxylate clamp type tetratricopeptide repeat proteins in Arabidopsis and rice as putative co-chaperones of Hsp90/Hsp70. PLoS One5:e12761. doi:10.1371 /journal.pone.0012761.
on November 7, 2019 by guest
http://jvi.asm.org/