JOURNAL OFVIROLOGY,Aug. 1993,p. 4543-4548 0022-538X/93/084543-06$02.00/0
Copyright © 1993, AmericanSociety forMicrobiology
Entry of Poliovirus into Cells
Does
Not
Require
a
Low-pH Step
LUIS PEREZANDLUIS CARRASCO*Centro deBiologia Molecular "Severo Ochoa"(Consejo Superior de Investigaciones
Cientificas-UniversidadAut6noma deMadrid), Universidad Aut6noma de Madrid,
CantoBlanco, 28049 Madrid, Spain Received 11 January 1993/Accepted 11 May 1993
The requirementofalow-pHstepduringpoliovirusentrywasinvestigated by using the macrolide antibiotic bafilomycin Al,which isapowerfulandselectiveinhibitor of thevacuolarproton-ATPases. Thus, virusessuch as Semliki Forest virus and vesicular stomatitis virus that enter cells through endosomes and need their acidification, are potently inhibited by bafilomycin Al, whereas poliovirus infection is not affected by the antibiotic. The presence oflysosomotropic agents such as chloroquine, amantadine, dansylcadaverine, and
monensinduringpoliovirusentrydidnotinhibitinfection,furthersupportingtheideathatpoliovirus doesnot depend on a low-pH step to enter the cytoplasm. The effect ofbafilomycin Al on other members of the Picornaviridaefamilywasalsoassayed.Encephalomyocarditisvirus entryinto HeLa cellswasnotaffected by the macrolide antibiotic, whereas rhinoviruswas sensitive. Coentry of toxins, such as ax-sarcin, with viral particles was potently inhibited by bafilomycin Al, indicating that an active vacuolar proton-ATPase is
necessaryfor theearlymembrane permeabilization (coentryofa-sarcin) induced bypoliovirustotakeplace. Poliovirus has a virion particle 30 nm in diameter with
icosahedralsymmetrythatiscomposedof 60copieseachof four structural proteins, VP1, VP2, VP3, and VP4, that surround the 7.5-kb genome covalently linked to
genome-bound protein VPg (15, 25). Thepoliovirus infective cycle
commences by interaction ofpoliovirus particles with
spe-cific cell surface receptors (20). Then,virus internalization proceeds by receptor-mediated endocytosis (31). Entry of poliovirus particlesdirectly through theplasma membrane, after binding to its receptor, has also been suggested as a portal of virus entry (8). Once the viral particles are in endosomes, theymustexitthisorganelleto starttranslation of thegenomicRNA in thecytoplasm. Itis stillamatterof debate to what extent poliovirus requires acidification of endosomes to allow passage of the genome through the endosomal membrane (16, 18, 31) or whether poliovirus accomplishes this task in a pH-independent fashion (12). Indirect experiments in which poliovirus particles were
complexedwithneutral red and incubatedwithweak amines
or monensin for several minutes and then incubated for a verylongtime(18to40h) suggestedthatpoliovirus requires a low-pH step for infectivity (16-18). Virus entry was
evaluated by measuring the cytopathic effect caused by poliovirus complexedwith neutralred after several rounds ofviralreplication (16-18). High concentrationsofamines, i.e.,0.3 mM chloroquineor70 mMNH4Cl (31), have been
used in some of these studies, in which poliovirus poly-merase production was strongly inhibited even when the drugswereadded 1 or2 h after virus entry(31). Certainly, thelysosomotropicagentsemployedhaveanumber of side effects that influence a wealth of enzymes and functions apartfromraisingthe endosomalpH (13, 27). Thus,itis not surprising that during the long incubation times, during which several rounds of viral replication take place, there
was inhibition ofpoliovirus, particularly if the drugs used
were irreversible (18). Shorter incubationtimes, testingthe
*Correspondingauthor.
inhibition ofpoliovirusby chloroquine ormonensin during
theuncoating period, ledto the idea thatpoliovirusentryis pH independent (12).
Studies onthe requirementofa low-pHstep forentryof other members of the Picomaviridae family indicate that foot-and-mouth disease virus replication is inhibited by lysosomotropic compounds (2, 6, 7). However, a direct
actionof theseagentsinthe initialstepsof virus infection is still uncertain, because chloroquinewas still active in the inhibition offoot-and-mouth disease virus replication even
when added 2.5 h after infection (2), whereas monensin blockedtheinternalizationoflabeled rhinovirus particlesin
HeLa cells (22). It has been claimed that this ionophore might even increasethe infectionof cells by
encephalomy-ocarditis (EMC)virus whenlow-pH medium is used during viruspenetration (18).
Endosomes,likeother organelles of thevesicularsystem,
areacidified bythe actionof the vacuolarproton-ATPases (21, 26). Theseenzymespumpprotonsinto the organellesat
the expense of ATP hydrolysis in an electrogenic fashion (21, 26). The effect of specific inhibition of the vacuolar proton-ATPase on animal virus infectivity has not been examined.Onlythe action of theunspecific ATPase inhibitor N,N'-dicyclohexylcarbodiimide on poliovirus has been as-sayed (17, 18). This compound was effective only when several rounds of viral replication occurred, but it had no
effect on poliovirus entry when the amount of virus was
increased(17, 18). Thetwomajor classes of inhibitors used raise the endosomal pH by accumulation ofweak amines (NH4Cl, chloroquine, andamantadine, etc.) orby dissipat-ingthe proton gradient of endosomesby ionophores (mon-ensin and nigericin) (27). These two classes ofcompounds leave the endosomal proton-ATPase untouched, and in principle, this enzyme continues functioning even in their
presence. BafilomycinAl (BFLA1)isapowerfuland
selec-tive inhibitor of the vacuolar proton-ATPases that has re-centlybeendiscovered(3, 21, 32).Toelucidate the require-mentsofalow-pHstepforpoliovirus infectivity,wedecided to test the effect this new antibiotic, and also those ofthe
4543
Vol. 67,No. 8
on November 9, 2019 by guest
http://jvi.asm.org/
other two classes ofinhibitorsdescribed above, on poliovi-rus entry. Our presentresults suggest that although poliovi-rus enters cells via an endosomal pathway, it does not require a low-pH step to infect cells.
MATERLILS ANDMETHODS
Cell lines,viruses, and media.Poliovirustype1 (Mahoney
strain)and EMC virus werepropagated,grown,andtitrated
by plaque assay in humanepithelioid carcinoma(HeLa)cells and mousefibroblasts (Lcells), respectively. Baby hamster kidney(BHK-21) cells were used for growth and titration of
Semliki Forest virus (SFV) and vesicular stomatitis virus
(VSV; Indianastrain). Humanrhinovirus(serotype 14)was
kindly provided by M. G. Rossmann (Purdue University).
BHK-21and African greenmonkey kidney(CV2)cells were grown in Dulbecco modified Eagle medium supplemented
with 8% fetal calf serum. HeLa andAfrican green monkey
kidney(Vero) cellswere grownin DulbeccomodifiedEagle
mediumsupplemented with 10%newborn calfserum. Inhibitors. BFLA1 wasprovided by K. Altendorf
(Univer-sity of Osnabruck, Osnabruck, Germany). a-Sarcin was a
generousgiftfrom D. M. Shuurmans(Department of Public
Health; Lansing, Mich.). Monensin, chloroquine, and dan-sylcadaverine were purchased from SigmaChemical Co., St.
Louis, Mo.
Analysis
of proteins by sodium dodecyl sulfate(SDS)-poly-acrylamide gel electrophoresis (PAGE). Cells grown in 24-well plates were infected at a multiplicity of infection (MOI) of 50 PFU per cell. After virus adsorption (min 30
postin-fection), the cells were incubated in Dulbecco modified
Eagle medium plus 2% calf serum. Protein labeling was
performed with 20 ,uCi of
[35S]methionine
per ml (1.45Ci/mmol;
Amersham International, Amersham, UnitedKingdom)inmethionine-free medium. Theradiolabeledcell
monolayers were dissolvedin samplebuffer (62.5mM Tris
[pH 6.8], 2% SDS, 0.1 M dithiothreitol, 17% glycerol,
0.024% bromophenol blue [as anindicator]). Samples were
heated at 90°C for 5 min and electrophoresed on a 15%
polyacrylamide gel overnight at 80 V. Fluorography was
carried outin 1 M sodium salicylate. Thegelswerefinally
dried andexposedtoAgfa X-ray films.
Measurement ofradioactivity incorporated into trichloro-aceticacid-precipitable material. The cellswereincubated for 1 h with the virus and a-sarcin (50 ,g/ml) in Dulbecco modified Eagle medium plus 2% calf serum, washed, and
pulse-labeledfor 1 hwith 2 ,uCiof
[35S]methionine
perml inmethionine- and serum-free medium. Radioactive medium
was removed, and the cellswere washed with
phosphate-buffered saline, treated with 5% trichloroacetic acid, and washed twicewith ethanol. The cellmonolayerwasallowed todrybeforeaddition of 0.1 MNaOH-1%SDS.Thesamples
weredissolved inliquidscintillation counting cocktail (For-mula 989; Du Pont, Boston, Mass.), and radioactivity was
quantitated inaliquid scintillation counter (1219 Rackbeta;
LKB, Bromma, Sweden).
RESULTS
Effects ofBFILA1 onentry of poliovirus. Comparison with enveloped RNA-containing viruses. To test the effect of BFLA1 on poliovirus entry, HeLa cells were treated with the compound in three different ways: (i) BFLA1 was presentcontinuously, from virus addition until the cells were labeledwith[35S]methionine; (ii) the compound was present
only duringthefirst 20 min of infection, and then the excess
POLIO
°1
BFLw o
=:
u a b cSFV
°- BFL
C°
u a b c
C
ED,.bA P
vsv
L& BFL
C:
° a b cG _
-NS(P)Iw
-N
...*I 5 2
-2C
_ __-.
C-rn 40 M-m 1.aI4
-mm-1C
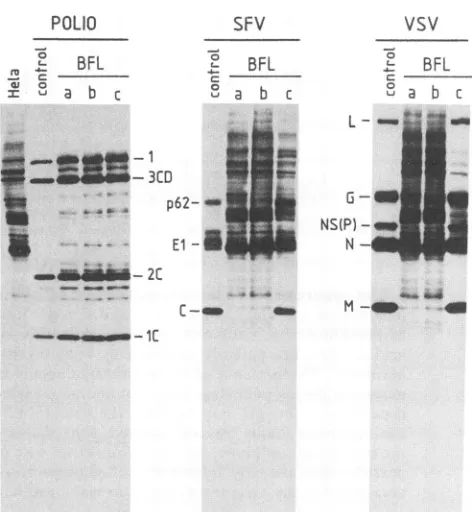
FIG. 1. Effects ofBFLA1addedatdifferent timesonpoliovirus (POLIO), SFV, and VSV. The MOI was 50 PFU per cellforthe three viruses.Labeling of infectedHeLacellswascarriedoutfrom 3.5 to4.5 h postinfection, asdescribedin Materials andMethods. Except in the controllanes,1,uMBFLA1 waspresent from addition of the virusto thecells(0hpostinfection) until the labelingperiod (lanesa), during only the first 20minofinfection(lanes b),orfrom 20 min postinfection (lanes c) until the labeling period. Lanes: control, infected cellsnottreatedwith thecompound; HeLa,protein synthesis inuninfected cells not treated with BFLA1.
virus and the inhibitor were removed; or (iii) BFLA1 was added 20 min after the virus and left throughout infection.
Figure 1 shows that BFLA1 did not prevent infection of
HeLacellsby poliovirusunder any of the threeconditions
tested. In contrast, SFV and VSVwerepotentlyinhibitedby
BFLA1, but onlywhen the compound was present
during
virusentry(i.e., conditionsi andii). Additionof BFLA1 20
minafter the virus
(condition
iii)didnotblock SFVorVSV infection, indicatingthat this agent doesnot inhibit subse-quent stepsof viralinfectionthat takeplaceafter the first 20minof thereplication cycle.WetestedtheinhibitionofSFV
attachment and internalization with radioactively labeled virions and found no inhibition by BFLA1 (results not
shown).
Thereare two additional points inFig. 1 that should be
emphasized. One is that BFLA1 does not inhibit cellular
translation, evenafterlongincubationtimes (seelaneafor
both SFV and VSV). After 6 h of incubation at 1 ,uM
BFLA1, cellularproteinsynthesiswas 100%of thecontrol value (datanotshown). The second aspect is the very low concentrations of BFLA1used, i.e., 1 ,uM, whichis about 100-fold lower than the concentrations ofchloroquine
nor-mally used to block SFV replication. The concentration
dependence of the effect of BFLA1on SFV andpoliovirus
replicationis shown inFig.2. Concentrationsof the
antibi-oticaslowas0.5,uM inhibitedSFVreplication,whereasan
almost 10-foldhigherconcentration of BFLA1(4 ,uM)hadno effectonpoliovirus.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.314.550.71.327.2]POLIOVIRUS ENTRY 4545
SFV
;
BFL
[iM]
0 0.5 1p107-
-p62- a.
E1,2-
&C-_.-.
POLIO
0 0.5
1
2
4
BFL[tM]
-____- - 3CD
2C
a
(mM)
, BFL
i (pM) AMAN CLO D-CAD MON
.!
A2 .^C, A-. 1^
mM)
BFL
:> (uM AMAN CLO DOAD MON
U,- - .. -.
11111,....
*g.,, _-- loop-a C1FIG. 2. Effects of different concentrations of BFLA1 on SFV
andpoliovirus (POLIO) infection of HeLa cells. Cellmonolayers
werepretreatedwith the indicatedconcentrations of BFLA1 15min
before infection with the virus(0hpostinfection; MOI,50 PFUper
cell). After 1 h of incubation with the virus and BFLA1, the monolayerswerewashed and incubated with fresh medium for 5 h postinfection. Labeling with [35S]methionine was carried out as
indicated in Materials and Methods. Proteinswerelabeled from 5to
6 hpostinfection with 15 pCiof[35S]methionine perml in
methi-onine-free medium andanalyzed bySDS-PAGE.
Comparison of BFLA1 and otherlysosomotropically active compounds. Although BFLAl potently blocked replication of the enveloped RNA-containing viruses tested, it was possible apriori that, for unknownreasons, thiscompound
was active only on these viruses but noton poliovirus. It could bespeculated thatpoliovirus enters cells inan endo-somal subpopulationwhere thevacuolar proton-ATPase is not susceptible to BFLAL. To assay the effects of other agents known toraise thepHinendosomes, wetested the aminesamantadine,chloroquine, anddansylcadaverineand the ionophore monensin against poliovirus. Amines are thoughttoaccumulate inendosomes and otherorganelles of the vesicular system, wheretheybecomeprotonatedandare
thus lesspermeable, leadingtoanincreaseintheendosomal pH (27). On the otherhand,monensin isan ionophorethat exchanges Na+/H+ in favor ofan H+ gradient, thus dissi-pating the ApH createdby the proton-ATPase pump (27). Since there isnonetmovementofcharges by monensin, AtI is not affected by the ionophore. These compounds were assayedover awiderangeofconcentrations.Figure3shows the results obtainedwith the lowest andhighest concentra-tions tested on poliovirus. None of these compounds blockedpoliovirus replication.Thehighestconcentrations of amantadine andchloroquineanalyzed, andalso dansylcada-verine, decrease the synthesis ofpoliovirus proteins, most likely because of the toxic effects that these agents have under those conditions (27). In no case was there even a slightincrease of cellularprotein synthesisthat wouldbean
FIG. 3. Effects ofseveralinhibitorsoninfection of HeLacellsby poliovirus(POLIO) (a)orSFV (b). Each compound was added to cellsattheconcentrationsindicatedat15 minbefore virus infection andweretreated asdescribedin the legend to Fig. 2. Synthesis of late viral proteinswasanalyzedby SDS-PAGE as described in the legendtoFig. 2. Proteinsynthesiswas measuredin cells infected withpoliovirusorSFV andtreated with the following compounds: BFL, BFLA1; AMAN, amantadine; CLQ, chloroquine; D-CAD: dansylcadaverine; MON, monensin.
indication that the entry ofpoliovirus was compromised by thesecompounds. As a control, the effects of these agents on SFV infection were alsotested (Fig. 3). All of them hadan inhibitory effect on SFV infection, although amantadine and dansylcadaverine were less effective than the other inhibi-tors at the concentrations tested (compare levels of viral peptideC). Therefore, entry of poliovirus is not affected by agents whichincrease the endosomal pH by three different
mechanisms, i.e., (i) blockade of the vacuolar
proton-ATPase with BFLA1, (ii) buffering of H+ by weak amines, and(iii) dissipation of the ApH by monensin.
Inprevious studies, HeLa S3 cells were used to analyze
therequirement for a low-pHstep forpoliovirus entry into
cells (16-18). Therefore, we decided to test the effects of BFLA1 onpoliovirus entry in different cell lines. Figure 4 shows that BFLA1 did not reduce poliovirus infection of HeLa S3, CV2, or Vero cells, suggesting that the uptake mechanismsarelikelytobesimilar inall of these cell types.
Coentry of a-sarcin with poliovirus. Effect of BFLA1.
Poliovirus is able to cointemalize and efficiently deliver protein toxins during entry into cells (1, 4). The exact mechanismbywhich this occurs is not fully understood, but virionuncoating is necessary for this process to occur (1). It has beenspeculatedthatdelivery of protein toxins by animal virusesinvolves disruptionofendosomes (5, 10), but other more selective mechanisms are possible. a-Sarcin had no effectonprotein synthesis when addedtouninfected HeLa cells, withor without BFLA1 (Fig. 5c), whereas in
agree-ment with previous findings, the toxin efficiently blocked
translation when added togetherwith virion particles (Fig. 5a).Thiscointernalization of theproteintoxinby poliovirus
waspotentlyblockedby BFLA1. These results suggest that
anactive vacuolar proton-ATPase is necessary for efficient
delivery of ax-sarcin to the cytoplasm. In addition, they
constitute indirect evidence that poliovirus entry occurs
through the endosomal pathway, because the coentry of
VOL.67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.315.561.77.267.2] [image:3.612.63.301.79.334.2]CV2
BFL[pM] 5 0 2
4' _ _ -1
-_ -3CD
- 2C
-1C
Vero
_ze
M 0 2
i.
.-._-1 '-- -3CD..-GM-2C
-lom-1C
FIG. 4. Poliovirus infection of different cell lii BFLA1.CV2, Vero, and HeLa S3 cellswereinfecte rusatanMOIof50 PFUpercell. The cellswerepr
2)ornot(lanes 0) at15min before infection with 2 indicated and incubated in thepresence ofthecom
virus for 1 h. At that time (1 hpostinfection), the r
washed and then incubated with fresh medium fo postinfection, protein synthesisinthe infectedcellsv
asdescribedin thelegendtoFig.2.
a-sarcin induced by poliovirus does notoccuu ence ofan inhibitorofan endosomal enzyme
coentry of alpha-sarcin induced by Sendai blockedatallby BFLA1,because this virusen fusion with the plasma membrane (14, 19) shown).Theseresults leadtotheconclusionthE all,thetoxindeliveredin thecytoplasm bypol
comesfromendosomes,becausepoisoningofI
a-sarcinwastotallyprevented byinhibition of proton-ATPase with BFLAL. For a model f entry illustrating the coentry of a-sarcin an
action of theinhibitors, seeFig. 7.
Effect ofBFIA1 on replication of other me
Picornaviridae family. The Picornaviridae fami into four genera: the enteroviruses, (poliovii
ruses,cardioviruses(EMC virus),andaftovirur mouth disease virus). The enteroviruses and
arecloselyrelatedbyanumber ofcharacteris the othertwogeneraare moredistant inevolui (24).Wetested theaction of BFLA1againstEl rhinovirus and compared its effect against ti RNA-containing virus VSV (Fig. 6). Infection4
by EMCvirus didnot seemtobe blocked byt antibiotic (e.g., compareviralpeptide alpha w outBFLA1), as occurswithpoliovirus, where or VSV infection was inhibited by BFLA1.
treatment ofEMC virus-infected cells with I
feredwith theinhibition of hosttranslation byI
did not investigate this effect. Figure 6 als( a-sarcinefficientlyentered thecytoplasminth the three virus particles. BFLA1 efficiently coentrymediatedbyanyof thethree viruses.A detailedanalyseswitheach of these virusesare
more firm conclusion regarding their mode (
HeLa S3
U
0
m 0 2
a-(1)0
I-u
0
z_
U) o
, 0
(L
LCi
2 Helai< -S
Iii
Ac
-POLIO
a-S + - + BFL
1 ..-
-3CL--_
-2C
-iin_ -2C
--1 C z t:°
X i
[E3FL.LIMs
nes. Effect of FIG. 5. a-Sarcin andpoliovirusentryinto HeLa cells. Effect of d withpoliovi- BFLA1.(a)Permeabilization of HeLa cells for a-sarcin at different -etreated(lanes MOIswith poliovirus. Symbols: El, poliovirus-infected cells incu-,uMBFLA1 as bated with 50,ugof a-sarcinpermlduringvirusentry(60min) bythe ipound and the protocol described in Materials andMethods; 0,cells treated with monolayerwas 1 FM BFLA1 during incubation with the virus and a-sarcin. Ir 4 h. At 5 h [35S]methionine incorporation into mock-infected cells (6.0 x 105 vasdetermined cpm)was 100% of the control value. Infection with the virus or treatmentwithBFLA1 ora-sarcininuninfected cellsyielded values of over85% ofthe control.(b) Effects of different concentrations of BFLA1 on thepermeabilization ofpoliovirus-infected HeLacells (MOI,100PFUpercell) fora-sarcin.BFLA1wasaddedtothe cells
rin the pres- 15 min before at-sarcin and virus infection. a-Sarcin entry was . In fact, the determined as inhibition of protein synthesis measured as described virus is not for panel a. (c) Electrophoretic analysis of[35S]methionine-labeled
terus
bydiret
proteins in cellsinfected withpoliovirus (POLIO)
in thepresenceof itersby
direct ot-sarcin and BFLA1. Mock-infected or poliovirus-infected HeLa(results not cells were incubated with orwithout 50
p,g
ofa-sarcin(a-s) per mlat most,ifnot with (+) or without(-) and 1 ,uM BFLA1 for 1 h. The cells were
liovirus entry washedand incubated with fresh medium until labeling of proteins
HeLa cellsby from 5to6hpostinfectionasdescribedinthelegendtoFig. 2. Ac, the vacuolar cellularproteinactin.
.or poliovirus
d the site of
results certainly suggest that only rhinovirus requires a Xmbers of the low-pH step forefficiententryintocells.
iily is divided
rus), rhinovi- DISCUSSION
ses
(foot-and-rhinoviruses Themechanisms usedbyanimalvirusesto crosscellular
,tics,whereas membranes and delivertheirgenomesinto thecytoplasmto
tionaryterms start infection is a matter of intensive research (14, 19).
MC virus and Animal viruses containing lipid membranes
accomplish
thishe enveloped task by fusion of the external envelope, either with the of HeLa cells plasma membrane, as occurswithSendaivirus,herpesvirus
the macrolide orvacciniavirus,orwith the endosomalmembrane,aswith
rith
and with- SFV, VSV, or influenza virus (14, 19). In the latter case, asrhinovirus fusion is promoted by conformational modifications of theRemarkably, viralglycoproteins after receptorbindingandacidification of
BFLA1 inter- endosomes (28, 29). Prevention of this acidification by
the virus. We lysosomotropic agents leads to inhibition of infection by D shows that SFV,VSV,orinfluenza virus(14). Modelstoaccountforthe
epresenceof entryofnonenveloped viruses,suchaspoliovirusor adeno-blocked this virus, are less well developed. We support the idea that
Ithoughmore attachment of poliovirus and internalization in endosomes
-needed fora leads toconformational changes in the virion particle,
par-of entry, our ticularlyinVP1 (11),whichcaninteract with the membrane
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.312.552.76.279.2] [image:4.612.62.297.77.279.2]POLIOVIRUS ENTRY 4547 EMCV
sarcin
BFL - + - +
E--I
EY- -_ _
HRV sarcn
p92-
p75-VPO .
vpI-
VP3-vsv
sarcin
mock
'a c C
u_
0 %1
X-FIG. 6. Action ofBFLA1 onthe entry ofseveralpicornaviruses into HeLa cells. Effectona-sarcin coentry. Monolayers of HeLa cells (Ohio strain) were mock infected (mock) or infected with human rhinovirus serotype14(HRV)at anMOI of 100 PFU per cell orwith EMC virusorVSVat anMOIof 25 PFU per cell. Lanes: +, treatmentof cells with 2 ILM BFLA1 (BFL) as described in the legend to Fig. 2; -, cellsnot treated with BFLA1. As indicated (sarcin), 50 p1g of a-sarcin per mlwasaddedtothe mediumatthe same time asthe virus. After incubation for 1 h, the cells were washed and incubatedwith fresh medium. [35SJmethionine-labeled
viralproteins wereanalyzed by PAGE. Samples containing equal amountsof totalproteinwereloaded in all of the lanes. Thecells werelabeledat5.5 hpostinfection for VSV and EMC virus andat 7.5hpostinfection for rhinovirus. Ac,cellularprotein actin.
and aid the insertion of
myristoylated
proteinVP4 into themembrane (11). This insertion not only destabilizes the
particle butmayalso allow interaction ofsomemoieties of
other viral
proteins
with themembrane(11).
Such aninser-tion couldopenaporein the membrane that allowspassage
of thegenome tothe
cytoplasm.
In thismodel, proteins
ofthe virus
particle
actsimilarly
to the B chains of someprotein toxins (30). In fact, we have reported that animal
virions, including poliovirusparticles,areabletoreplacethe
Bchains ofprotein toxins and promote the entry of the A
chain or effector
moiety
of the molecule(5,
9,23).
Werecently showed that for thisco-entryoftoxinswith
polio-virus, uncoatingof theparticleisrequired,in agreement with
themodel describedabove
(1).
Ourpresentfindings
ontheinvolvement of the vacuolar proton ATPase inthis
phenom-enonare of interest in
understanding
notonly
the entryofnonenvelopedvirusesbutalso thecoentry ofproteintoxins
inside cells.Thus,wecannotdissociatepoliovirusuncoating
from the coentry mechanisms,
arguing against
previousmodelsthatsuggestedthe rupture ofendosomes
during
virusentry
(1,
4, 5,10). Therefore, poliovirus
enters fromendo-somes into the
cytoplasm
and maintains the endosomalmembraneintact,because this rupture would lead toleakage
of the endosomalcontent,
including
oa-sarcin, into thecyto-plasm. Thus, after orduring poliovirus entry, the
modifica-tion introduced in the cellular membrane is usedbya-sarcin totraversethe membrane.Our present results favor the view that the force that pushes a-sarcin through the membrane may be the proton gradient generated by the vacuolar
proton-ATPase. Therefore, instead ofasimple mechanistic
model ofcreation ofaporethroughwhich molecules pass, other forces
(proton
motive,
electrical)
may be involved incX-sACINH
IBAFILOHYCINA
[image:5.612.327.564.67.246.2]N a-SARCIN H
a-SUrCIN H
FIG. 7. Schematic model of two potential mechanisms of polio-virus entry into cells. Poliopolio-virusinternalizationthrough endosomes ordirectcrossing of the plasma membrane would lead to insertion of structuralproteins intothe membrane. These proteins, together with the protongradient between endosomes and the cytoplasm gener-atedby theH'-ATPase,wouldtranslocate the toxin a-sarcin to the cell interior.
protein translocation.Inconclusion, the virus capsid protein
would be able tocouplethese forces to thetranslocation of
theproteinspresentinendosomes.
Two major questions must be answered to explain the
mechanism ofpoliovirus entry into cells.One iswhetherthe virus is internalizedthrough endosomes, and the second is
whetherendosomes need to beacidified for the virus to enter
cells. Webelieve that thesecondquestionhas beenclarified
by the present work. Since none of the agents known to
increase the endosomal pH prevent poliovirus infection
when they are present during entry, we conclude that a
low-pHstepisnotrequiredforpoliovirustoinfect cells. We
emphasize theunique mode of action ofoneoftheinhibitors
used in ourwork,i.e., BFLAL. Since this agentselectively
blocks thevacuolar proton-ATPases (3, 21, 32) and shows
activity at micromolar concentrations, we suggest that it
should replace other compoundswidely employed to
ana-lyze the low-pH requirements for animal virusentry(14, 19).
However,ourresults donotprovideadefinitiveanswer to
thefirstquestionraisedabove,i.e., whetherpoliovirus gains
access tothecytoplasmvia endosomes. We believethatthe
two possible mechanisms of poliovirus entry, i.e., direct
entry throughthe plasma membrane (8) and internalization
in endosomes (31), are still open. It is even possible that
poliovirususesboth of them.Althoughpoliovirus
permeabi-lizescells fora-sarcinandthisevent occurs inendosomes, it
is possible a priori that the virus has already released its
genomeintothe cytoplasm afterbindingtothe cellsurface
receptor and the virion proteins present in the membrane
andinternalized in endosomesarethe ones thatpermeabilize
the membrane for a-sarcin once the ApH has been created
(model in Fig. 7). Alternatively, it is possible that the
poliovirus particle can traverse the cell membrane only
during the internalization process. Recent electron
micro-scopic evidence illustrating entry of poliovirus into cells
favors the second mechanism(31). Furtherexperiments are neededtoclarifythis issue andgive us abetteridea of how the genome of anonenveloped animal virus is ableto cross
alipidmembrane to start infection.
VOL.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.69.309.70.251.2]ACKNOWLEDGMENTS
LuisPerez is the holder of a Comunidad Aut6noma de Madrid fellowship. Plan Nacional project BIO 92-0715, DGICYT project PB90-0177, and theFundaci6n Ram6nAreces areacknowledged for financialsupport.
REFERENCES
1. Almela, M. J., M. E. Gonzilez, and L. Carrasco. 1991.Inhibitors ofuncoating efficientlyblock the earlypermeabilization induced by poliovirus. J. Virol. 65:2572-2577.
2. Baxt, B. 1987. Effect of lysosomotropic compounds on early eventsin foot-and-mouth diseasevirus replication. VirusRes. 7:257-271.
3. Bowman, E. J., A. Siebers, and K. Altendorf. 1988. Bafilomy-cins:aclass of inhibitors of membraneATPasesfrom microor-ganisms, animal cells, and plant cells. Proc. Natl.Acad. Sci. USA85:7972-7976.
4. Carrasco, L. 1981. Modification of membrane permeability induced by animal viruses earlyininfection.Virology 113:623-629.
5. Carrasco,L., M. J.Otero, andJ. L. Castrillo. 1989. Modifica-tion of membrane permeabilitybyanimal viruses. Pharmacol. Ther. 40:171-212.
6. Carrillo,E. C., C. Giachetti, and R. Campos.1985. Earlysteps in FMDVreplication: further analysis on the effects of chloro-quine. Virology147:118-125.
7. Camillo,E.C., C. Giachetti, and R. H. Campos. 1984. Effect of lysosomotropic agents on the foot-and-mouth disease virus replication. Virology135:542-545.
8. Dunnebacke, T. H., J. D. Levinthal, and R. C.Williams. 1969. Entryand release ofpoliovirusasobserved byelectron micros-copyofcultured cells.J.Virol. 4:505-513.
9. Fernandez-Puentes, C., and L.Carrasco. 1980. Viral infection permeabilizes mammalian cellstoprotein toxins. Cell 20:769-775.
10. FitzGerald, D. J., R. Padmanabhan, I. Pastan, and M. C. Willingham. 1983. Adenovirus-induced release of epidermal growth factorandpseudomonas toxin intothe cytosol ofKB cells during receptor-mediated endocytosis. Cell 32:607-617. 11. Fricks, C. E., and J. M. Hogle. 1990. Cell-induced
conforma-tional changeinpoliovirus: externalization of the amino
termi-nus of VP1 is responsible for liposome binding. J. Virol. 64:1934-1945.
12. Gromeier, M., and K. Wetz. 1990. Kineticsof poliovirus un-coating in HeLa cells in a nonacidic environment. J. Virol. 64:3590-3597.
13. Hiebsch, R. R., T. J. Raub, and B. W. Wattenberg. 1991. Primaquine blocks transport by inhibiting the formation of functional transport vesicles. Studies in a cell-free assay of protein transportthrough theGolgi apparatus. J. Biol. Chem. 266:20323-20328.
14. Hoekstra, D., and J. W. Kok. 1989. Entry mechanisms of enveloped viruses. Implications for fusion of intracellular
mem-branes.Biosci.Rep. 9:273-305.
15. Hogle, J. M., M. Chow, and D. J. Filman. 1985. Three-dimen-sional structure of poliovirus at 2.9 A resolution. Science 229:1358-1365.
16. Madshus,I.H.,S. Olsnes, and K. Sandvig. 1984.DifferentpH requirementsforentryof thetwopicornaviruses, human rhino-virus 2 and murineencephalomyocarditis virus. Virology 139: 346-357.
17. Madshus, I. H., S.Olsnes,and K.Sandvig.1984. Requirements for entry ofpoliovirus RNA into cells at lowpH. EMBO J. 3:1945-1950.
18. Madshus, I. H.,S.Olsnes,and K.Sandvig. 1984. Mechanism of entryinto thecytosol of poliovirustype 1:requirement for low pH. J.Cell Biol.98:1194-1200.
19. Marsh, M., and A. Helenius.1989.Virusentryinto animalcells. Adv. VirusRes.36:107-151.
20. Mendelsohn, C. L., E. Wimmer, and V. R. Racaniello. 1989. Cellularreceptorforpoliovirus: molecular cloning, nucleotide sequence,andexpression ofa newmemberof the immunoglob-ulinsuperfamily. Cell 56:855-865.
21. Nelson, N., and L. Taiz.1989. Theevolution ofH+-ATPases. Trends Biochem. Sci. 14:113-116.
22. Neubauer, C., L. Frasel, E. Kuechler, and D. Blaas. 1987. Mechanism of entryof human rhinovirus 2 into HeLa cells. Virology 158:255-258.
23. Otero, M. J., and L.Carrasco.1987.Proteinsarecointernalized with virionparticles duringearlyinfection.Virology 160:75-80. 24. Palmemberg, A. 1990. Proteolytic processing of picomaviral
polyprotein.Annu.Rev.Microbiol. 44:602-623.
25. Rossmann, M.G., andJ. E. Johnson. 1989. Icosahedral RNA virusstructure.Annu. Rev.Biochem.58:533-573.
26. Schneider, D. L.1987. Theproton pumpATPaseoflysosomes and related organelles of the vacuolar apparatus. Biochim. Biophys. Acta 895:1-10.
27. Seglen, P. 0. 1983. Inhibitors oflysosomalfunction. Methods Enzymol. 96:737-764.
28. White, J. M.1990.Viralandcellular membrane fusionproteins. Annu. Rev.Physiol. 52:675-697.
29. Wiley, D.C.,andJ. J.Skehel. 1987.Thestructureand function of thehemagglutinin membrane glycoprotein of influenza virus. Annu.Rev.Biochem. 56:365-394.
30. Wilson,B. A., and R. J. Collier. 1992. Diphtheria toxin and Pseudomonasaeruginosa exotoxin A: active-sitestructureand enzymic mechanism. Curr. Top. Microbiol. Immunol. 175:27-41.
31. Zeichhardt,H., K. Wetz, P.Willingmann,and K.0.Habermehl. 1985.Entry ofpoliovirustype 1andmouseElberfeld(ME)virus intoHEp-2cells:receptor-mediated endocytosis and endosomal
orlysosomaluncoating.J. Gen.Virol.66:483-492.
32. Zeuzem, S.,P.Feick,P.Zimmermann,W.Haase,R.A.Kahn, andI. Schulz. 1992. Intravesicular acidification correlates with binding ofADP-ribosylation factortomicrosomal membranes. Proc.Natl. Acad. Sci. USA 89:6619-6623.

![FIG. 5.batedwithwashedMOIsBFLA1.protocolofcpm)forproteinstreatmentBFLA1determinedot-sarcincells(MOI,[35S]methioninecellular1from15 FM over min a-Sarcin and poliovirus entry into HeLa cells](https://thumb-us.123doks.com/thumbv2/123dok_us/1299364.83018/4.612.62.297.77.279/batedwithwashedmoisbfla-protocolofcpm-forproteinstreatmentbfla-determinedot-sarcincells-methioninecellular-sarcin-poliovirus.webp)

