Copyright © 2002, American Society for Microbiology. All Rights Reserved.
NOTES
Induction of Anti-Human Immunodeficiency Virus Type 1 (HIV-1)
CD8
⫹
and CD4
⫹
T-Cell Reactivity by Dendritic Cells Loaded with
HIV-1 X4-Infected Apoptotic Cells
Xiao-Qing Zhao,
1Xiao-Li Huang, Phalguni Gupta,
1,2Luann Borowski,
1Zheng Fan,
1Simon C. Watkins,
3Elaine K. Thomas,
4and Charles R. Rinaldo, Jr.
1,2*
Department of Infectious Diseases and Microbiology, Graduate School of Public Health,1and Departments of Pathology2and Cell
Biology and Physiology, School of Medicine,3University of Pittsburgh, Pittsburgh, Pennsylvania 15261, and
Immunex Corporation, Seattle, Washington 981014
Received 11 July 2001/Accepted 29 November 2001
T-cell responses to X4 strains of human immunodeficiency virus type 1 (HIV-1) are considered important in controlling progression of HIV-1 infection. We investigated the ability of dendritic cells (DC) and various forms of HIV-1 X4 antigen to induce anti-HIV-1 T-cell responses in autologous peripheral blood mononuclear cells
from HIV-1-infected persons. Immature DC loaded with HIV-1 IIIB-infected, autologous, apoptotic CD8ⴚcells
and matured with CD40 ligand induced gamma interferon production in autologous CD8ⴙand CD4ⴙT cells.
In contrast, mature DC loaded with HIV-1 IIIB-infected, necrotic cells or directly infected with cell-free HIV-1 IIIB were poorly immunogenic. Thus, HIV-1-infected cells undergoing apoptosis serve as a rich source of X4
antigen for CD8ⴙand CD4ⴙT cells by DC. This may be an important mechanism of HIV-1 immunogenicity
and provides a strategy for immunotherapy of HIV-1-infected patients on combination antiretroviral therapy.
Progression of human immunodeficiency virus type 1 (HIV-1) infection is related to a switch in predominance of macrophage-tropic strains that use the CCR5 coreceptor (termed R5 virus) to T-cell-tropic strains that use CXCR4 as their major coreceptor (termed X4 virus) (7, 27, 43, 44). Fail-ure of CD8⫹ and CD4⫹ T-cell responses to control HIV-1
infection may be a significant factor leading to unimpeded replication of X4 virus and the development of AIDS (48). Although the recent advent of combination antiretroviral ther-apy has resulted in dramatic improvements in control of HIV-1 replication in persons chronically infected with HIV-1 (18, 36), it does not completely restore anti-HIV-1 T-cell responses (12, 26, 35–38). Low levels of residual virus remain in such persons and increase when drug therapy is discontinued (13). Thus, therapeutic approaches are needed that enhance T-cell immu-nity to HIV-1 for more complete control of HIV-1 infection.
Dendritic cells (DC) are the most potent antigen-presenting cells for the induction of antiviral T-cell responses through their expression of high levels of major histocompatibility com-plex (MHC) class I and II molecules and costimulatory mole-cules, such as CD40, CD80, and CD86, and the production of immunomodulating cytokines such as interleukin-12 (IL-12) and IL-15 (6). Current evidence indicates that immature DC (iDC) are highly efficient at capturing and processing antigens (6). Subsequent maturation of the iDC by ligation of CD40 with CD40 ligand (CD40L or CD154) expressed on CD4⫹T
cells upregulates MHC class I and II molecules and costimu-latory molecules, greatly enhancing the presentation of antigen to T cells by these mature DC (mDC) (47). In the classic endogenous pathway, proteins produced during viral replica-tion in the antigen-presenting cells are proteolytically cleaved in the cytosol (34). The resulting peptides are transported to the endoplasmic reticulum, where they complex with MHC class I molecules and then travel through the Golgi to the cell surface. In the exogenous pathway, viral proteins are ingested from the extracytosolic space into endosomal vesicles. There, the proteins are digested and the viral peptides are complexed with MHC class II molecules before transport to the cell mem-brane.
Some viruses do not replicate efficiently in DC, suggesting that there are alternative mechanisms to the conventional, endogenous MHC class I pathway for the induction of CD8⫹
T-cell responses to these viral antigens (29, 50). This has been related to uptake by DC of exogenous antigen in the form of virus-infected, apoptotic, or necrotic cells, followed by process-ing through nonconventional pathways and cross-presentation of antigen in the context of MHC class I molecules to CD8⫹T
cells (2, 3, 20, 21, 29, 45, 46). In HIV-1 infection, iDC do not support efficient replication of X4 strains due to low expression of the CXCR4 coreceptor, whereas they express higher levels of CCR5 and more efficiently support R5 virus replication (11, 17, 29). Thus, induction of anti-HIV-1 CD8⫹T-cell responses
to X4 virus may at least in part be due to uptake of X4 antigens by iDC and cross-presentation by HLA class I molecules on mDC. These viral antigens could be derived from cells that have been productively infected by X4 strains and have under-gone apoptosis (4). A similar process of uptake of exogenous,
* Corresponding author. Mailing address: A427 Crabtree Hall, Uni-versity of Pittsburgh Graduate School of Public Health, 130 DeSoto St., Pittsburgh, PA 15261. Phone: (412) 624-3928. Fax: (412) 624-4953. E-mail: rinaldo@pitt.edu.
3007
on November 8, 2019 by guest
http://jvi.asm.org/
nonreplicating viral antigens by iDC, with processing through the HLA class II pathway, presumably leads to induction of anti-HIV-1 CD4⫹T-cell responses.
We therefore studied activation of anti-HIV-1 T-cell re-sponses in peripheral blood mononuclear cells (PBMC) of persons with chronic HIV-1 infection by autologous DC loaded in vitro with various forms of HIV-1 X4 antigen. The study subjects for the T-cell immunity experiments were eight HIV-1-seropositive homosexual men from the Pittsburgh, Pa., portion of the Multicenter AIDS Cohort Study who were chronically infected with HIV-1 (Table 1). One of these sub-jects (S5) was not receiving antiretroviral therapy, while the seven other subjects were being treated with combination an-tiretroviral drug therapy of a protease inhibitor and two re-verse transcriptase inhibitors. An additional HIV-1 chronically infected person on combination drug therapy was used for the DC phenotyping studies. Each subject gave written, informed consent approved by the University of Pittsburgh Institutional Review Board. Viral loads in plasma ranged from undetectable (⬍50 copies/ml) to⬎30,000 copies per/ml at the time of this study, and CD4⫹T-cell counts ranged from 339 to 1,215 per ml
of blood. Seven healthy HIV-1-negative persons served as con-trols.
For preparation of antigens, PBMC from the HIV-1-sero-positive subjects were separated from freshly donated, hepa-rinized blood by density centrifugation with Ficoll-Hypaque (Amersham Pharmacia Biotech, Piscataway, N.J.). CD8⫺cells
were obtained by treatment of the PBMC with
immunomag-netic beads specific for CD8 (BD Immunocytometry Systems, San Jose, Calif.). We noted in preliminary experiments that mDC loaded with HIV-1 IIIB-infected, CD8⫺or CD4⫹
neg-atively selected, apoptotic cell preparations induced compara-ble numbers of gamma interferon (IFN-␥)-producing cells (data not shown). Thus, we used CD8⫺cell preparations in
subsequent studies of anti-HIV-1 T-cell responses. The CD8⫺
cells were stimulated with anti-CD3 monoclonal antibody (MAb) (200 ng of OKT-3/ml; Ortho, Raritan, N.J.) and human recombinant IL- 2 (rIL-2; 100 U/ml; Chiron, Emeryville, Cal-if.) for 4 to 5 days in RPMI 1640 medium supplemented with 20% heat-inactivated fetal calf serum and antibiotics (Gibco, Grand Island, N.Y.). The CD8⫺cells were then treated with
Polybrene (5g per 106 cells; Sigma Biosciences, St. Louis,
Mo.) for 1 h, pelleted by centrifugation at 250⫻gfor 10 min, and superinfected with HIV-1 IIIB (5) for 2 h at 37°C (50 to 100 ng of p24 per 106 cells). The superinfected cells were
washed and adjusted to 106cells/ml and cultured in complete
RPMI 1640 medium supplemented with rIL-2 (100 U/ml) for another 4 to 5 days. Infection was monitored by production of HIV-1 p24 antigen in culture supernatant by a commercial p24 enzyme immunoassay (NEN Life Sciences, Boston, Mass.). Levels of p24 in the CD8⫺ cells from the HIV-1-infected
subjects ranged from 0.1 to 540 ng/ml for cells not superin-fected with HIV-1 IIIB compared to 6 to⬎1,160 ng/ml for cells superinfected with HIV-1 IIIB (Table 1).
To inactivate HIV-1 and induce apoptosis, the CD8⫺cells
were treated with psoralen [10g/ml; 4⬘-(aminomethyl)-4,5⬘
,8-TABLE 1. IFN-␥response of PBMC stimulated with autologous DC loaded with HIV-1 antigen
Subject Duration (yr)HIV positive CD4
⫹
T-cell count (cells/l)
Plasma HIV RNA (copies/ml) Therapy
a
p24 antigen (ng/ml)
in CD8⫺cells SFC/106PBMC
Without
IIIB With IIIB iDC iDC⫹AP iDC⫹AP⫺IIIB mDC mDC⫹AP mDC⫹AP⫺IIIB
HIV positive
S1 ⬎13.5 594 ⬍50 L, Z, I 12 172 10 90 250 240 70 2,040
S2 ⬎5.8 555 ⬍50 L, Z, N 0.1 6 4 10 0 65 1,765 1,715
S3 ⬎9.3 655 69 L, Z, I 540 ⬎1,160 0 100 80 190 460 880
S4 5.6 522 ⬍50 L, Z, I NDb ⬎1,000 0 20 90 470 210 2,320
S5 ⬎5.0 1,215 30,157 None 12 614 0 520 680 280 670 1,940
S6 ⬎14.5 588 ⬍50 L, D, I 23 80 0 0 0 60 120 880
S7 17.5 1,081 306 R, D, I 0.1 54 20 30 65 280 245 750
S8 ⬎12.9 601 ⬍50 d, D, E 0.1 ⬎10 0 260 120 20 20 1,280
Median ⬎11.1 597 58c 12 80 0d 60e 85f 215g 227h 1,498
HIV-1 negative
N1 ⬍0.01 7.5 65 25 25 80 30 40
N2 ⬍0.01 11 345 215 155 170 250 180
N3 ⬍0.01 5.5 0 0 0 150 80 150
Mediani 0 8 65 80 25 150 80 150
aL, lamivudine; Z, zidovudine; I, indinavir; N, nelfinavir; D, stavudine; R, ritonavir; d, didanosine; E, efavirenz. bND, not done.
cFor calculation of the median for viral load,⬍50 copies/ml were considered as 25 copies/ml.
dAnalyzed by Wilcoxon signed rank test for the HIV-1-positive group: iDC versus iDC⫹AP,P⫽0.022; iDC versus iDC⫹AP⫺IIIB,P⫽0.035; iDC versus mDC P⫽0.014.
eAnalyzed by Wilcoxon signed rank test for the HIV-1-positive group: iDC⫹AP versus iDC⫹AP⫺IIIb,P⫽not significant (ns); iDC⫹AP versus mDC⫹AP,P⫽
ns.
fAnalyzed by Wilcoxon signed rank test for the HIV-1-positive group: iDC⫹AP⫹IIIb versus mDC⫹AP⫺IIIb,P⫽0.014.
gAnalyzed by Wilcoxon signed rank test for the HIV-1-positive group: mDC versus mDC⫹AP,P⫽ns; mDC versus mDC⫹AP⫺IIIbP⫽0.014. hAnalyzed by Wilcoxon signed rank test for the HIV-1-positive group: mDC⫹AP versus mDC⫹AP⫺IIIb,P⫽0.021.
iAnalyzed by Wilcoxon signed rank test for the HIV-1-negative group: for all data comparisons,P⫽ns.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:2.587.43.542.85.305.2]trimethylpsoralen hydrochloride; Calbiochem, La Jolla, Calif.] and UV light irradiation (312-nm wavelength calibrated to provide 0.86 mJ/cm2/s for 6 min; Derma Control, Fisher
Sci-entific, Pittsburgh, Pa.) (referred to here as psoralen and UV irradiation [PUV] treatment). The cells were washed three times and cultured for an additional 6 h to allow time for apoptosis. Apoptosis was confirmed by flow cytometry (EPICS XL; Beckman Coulter, Fullerton, Calif.) by using the TUNEL (terminal deoxynucleotidyltransferase-mediated dUTP-biotin nick end labeling) assay (In Situ Cell Death Detection Kit; Boehringer Mannheim, Indianapolis, Ind.). Without PUV treatment, ca. 10% of the CD8⫺cell preparations not
super-infected with HIV-1 IIIB were apoptotic, compared to 25 to 30% apoptotic cells in CD8⫺cell preparations that were
super-infected with HIV-1 IIIB. Apoptosis in both of these cell prep-arations increased to 70% after PUV treatment (termed AP and AP-IIIB, respectively). Necrotic preparations of HIV-1 IIIB-superinfected (NE-IIIB) and nonsuperinfected (NE) CD8⫺ cells were made by three or four rapid freeze-thaw
cycles of the cells in dry ice-ethanol and 37°C water baths. DC were derived from autologous CD14⫹ cells that were
enriched by positive selection with anti-CD14 MAb-coated immunomagnetic beads (MACS Isolation Kit; Miltenyi Biotec, Auburn, Calif.). The purity of CD14⫹cells was⬎95%, based
on staining with MAb to CD14 (BD) and analysis by flow cytometry. The CD14⫹cells were cultured in 1,000 U of
gran-ulocyte-monocyte colony-stimulating factor/ml and 1,000 U of IL-4 (Schering-Plough, Kenilworth, N.J.)/ml in complete RPMI 1640 medium for 4 to 5 days to obtain iDC as previously described (14). To induce maturation, the iDC were treated with 1.0 to 2.5 g of trimeric CD40L (Immunex, Seattle, Wash.)/ml at day 4 or 5 for two additional days at 37°C in the cytokine-supplemented medium.
To determine uptake of the apoptotic and necrotic cell prep-arations by iDC or mDC, HIV-1 IIIB-infected and uninfected, apoptotic and necrotic CD8⫺ cells were stained with
mem-brane fluorophore PKH 26 as per the manufacturer’s instruc-tions (Sigma). They were then cocultured for 12 h at 37°C at a ratio of 2:1 with iDC or mDC that were stained with HLA-DR–fluorescein isothiocyanate (FITC). Flow cytometric anal-ysis was performed by gating on large cells with high side scatter, and double-positive cells were enumerated. As shown in Fig. 1A, the uptake of both the CD8⫺apoptotic and necrotic
cell preparations by iDC was much greater than by mDC (38% versus 6% for apoptotic cells and 23% versus 0.2% for necrotic cells). Furthermore, 30 to 50% of the iDC contained intracel-lular CD8⫺cells and cell debris as visualized by confocal laser
scanning microscopy (Leica Lasertechnik, Heidelberg, Ger-many) (Fig. 1B). Therefore, for the T-cell stimulation experi-ments, the apoptotic and necrotic CD8⫺cell preparations were
added to iDC at a 2-to-1 ratio (cell equivalent portions were used for the necrotic cell preparations) on day 4 or 5, incu-bated for 12 h, and then treated with either medium (to main-tain iDC) or CD40 ligand (CD40L) (to induce maturation) for 2 additional days before use in the T-cell studies. Additionally, iDC were infected on day 4 or 5 with 300 ng of p24 per 106cells
of replication-competent, cell-free HIV-1 IIIB or PUV-treated HIV-1 IIIB (prepared by PUV treatment as with the CD8⫺
cells and three sequential washes in medium for 1 h each at 22,000 ⫻ g at 4°C) for 2 h at 37°C and washed twice with
medium to remove unadsorbed virus. These HIV-1 IIIB-in-fected iDC were then cultured with medium alone or CD40L, as stated above, for the T-cell activation experiments.
Previous studies have shown increased expression of matu-ration markers on DC after treatment with either apoptotic or necrotic cells (15, 24, 40), which could relate to changes in the processing and presentation of antigen. Therefore, the expres-sion of relevant surface markers was assessed on iDC and mDC for possible effects of the antigen preparations on DC maturation. The iDC and mDC were analyzed by staining with
FIG. 1. (A) Uptake of apoptotic and necrotic, non-HIV-1 IIIB-infected, CD8⫺cells (subject N7) by iDC and mDC, as demonstrated
by flow cytometry. Similar results were obtained for HIV-1 IIIB-in-fected, CD8⫺cells. (B1) iDC ingested the apoptotic CD8⫺ cells, as
demonstrated by confocal laser scanning microscopy. (B2 to B5) Sec-tion series were collected from the apical to the basal surface of cells at 0.5-m section intervals at a resolution of 1,024⫻1,024 pixels.
on November 8, 2019 by guest
http://jvi.asm.org/
phycoerythrin (PE)-Cy5-conjugated anti-HLA-DR MAb (Im-munotech, Marseille, France) and PE-conjugated MAb to lin-eage markers CD3, CD14, CD16, CD19, and CD56 (BD). The cells were assayed by flow cytometry by gating on large cells with high side-scatter and analysis on a two-color histogram. Cells that were HLA-DR⫹ and CD3⫺, CD14⫺, CD16⫺,
CD19⫺, and CD56⫺ were defined as DC. These DC were
further analyzed after staining with PE-conjugated MAb spe-cific for CD80 (BD), CD83 (Immunotech), and HLA-ABC (BD) and FITC-conjugated MAb to CD86 (Ancell, Bayport, Minn.). Appropriate isotype-matched controls were used throughout the course of the study. The expression of the cell surface markers was assessed as the mean fluorescence inten-sity (MFI) and the percent positive cells.
The addition of CD40L to DC of three uninfected and three HIV-1-infected persons resulted in a similar pattern of surface marker changes, so the results were combined as shown in Fig.
2. The data show that there were significant increases in sur-face marker expression on iDC, i.e., HLA DR, CD80, and CD86, after loading with some of the antigen preparations (Fig. 2). Interestingly, all of these increases were noted with the iDC⫹AP-IIIB preparations. However, iDC⫹AP prepara-tions only increased the expression of CD86, and iDC⫹NE only increased the expression of HLA-DR and CD86. These changes were found in DC from both HIV-1-infected and uninfected subjects (data not shown). Thus, the modifications in surface marker expression were not dependent on HIV-1 infection in the antigen preparations or the study subjects.
The CD40L-treated mDC from HIV-1-infected and unin-fected persons had increases in surface expression of HLA-DR, HLA-ABC, CD80, CD86, and CD83 compared to iDC (Fig. 2), thus confirming our previous work (14) and that of other investigators (33). Furthermore, there was an increase in the number of mDC compared to iDC expressing CD80 (mean ⫾standard error: 56%⫾6% to 96%⫾1%,P⫽0.002; paired T test), CD86 (41%⫾11% to 98%⫾1%,P ⫽0.003), and CD83 (8%⫾3% to 66%⫾15%,P⫽0.01). The numbers of HLA-DR and HLA ABC positive cells were high in both iDC and mDC (95 to 99%), as is characteristic of these cells (6).
We next investigated the capacity of DC loaded with the apoptotic, necrotic, and cell-free X4 virus preparations to in-duce IFN-␥ production in autologous T cells. An enzyme-linked immunospot assay was used to assess the production of IFN-␥from single, antigen-specific T cells, as described else-where (14). A concentration of 104 antigen-loaded iDC or
mDC was added to 105autologous PBMC (i.e., a 1:10 ratio)
and incubated overnight at 37°C. The results were expressed as the number of spot-forming cells (SFC) per 106PBMC. This
assay had an average, within-sample variation of 12% among replicates of⬎100 PBMC samples tested in our laboratory.
In initial experiments with autologous PBMC from three HIV-1-infected subjects, we found that either CD8⫺cells,
PBMC, iDC, or mDC did not produce appreciable IFN-␥(0 to 5 SFC/105cells; data not shown). The iDC, or iDC loaded with
[image:4.587.50.280.73.406.2]the various antigens, induced low levels of IFN-␥in PBMC (Fig. 3). However, PBMC produced high levels of IFN-␥when stimulated by mDC loaded with HIV-1 IIIB-superinfected,
FIG. 2. Effect of CD40L and various antigen preparations on the expression of costimulatory and HLA class I and II molecules on DC. Data shown are the means (⫹the standard error) for the MFI of DC from three uninfected persons (N2, N3, and N4) and three HIV-1-infected subjects (S7, S8, and S9). ⴱ, Comparison of the effect of antigen loading on iDC expression of HLA-DR (iDC⫹AP-IIIB,P⫽
0.019; iDC⫹NE,P⫽0.047), CD80 (iDC⫹AP-IIIB,P⫽0.04), and CD86 (iDC⫹AP,P⫽0.032; iDC⫹AP-IIIB,P⫽0.001; iDC⫹NE,P⫽
0.029) (pairedttest);ⴱⴱ, comparison of the iDC with matched mDC (P
ⱕ 0.02, except for P ⫽ 0.038 and 0.044 for CD83 expression on NE-loaded and NE-IIIB-loaded DC, respectively). All other compar-isons wereP⬎0.05.
FIG. 3. IFN-␥production by PBMC from three subjects chronically infected with HIV-1 (S3, S7, and S8) induced by iDC or mDC loaded with the various antigen preparations.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.587.300.542.75.232.2]autologous, apoptotic CD8⫺ cell preparations (mDC⫹
AP-IIIB) compared to stimulation with mDC⫹AP (Fig. 3). This effect was dependent on the presence of the mDC, since only 0 to 1 SFC/105cells was detected when the CD8⫺cell
prepa-rations were added to autologous PBMC without mDC. The IFN-␥response was also dependent on the concentration of mDC⫹AP-IIIB added to the PBMC cultures, since ratios of 1:5 and 1:20 of DC to responder PBMC resulted in propor-tionally higher and lower numbers of IFN-␥-producing cells, respectively, than the standard 1:10 ratio (data not shown). In contrast to the effects of the apoptotic cell preparations, mDC loaded with either of the necrotic cell preparations (mDC⫹NE or mDC⫹NE-IIIB), or infected with cell-free, viable HIV-1 IIIB (Fig. 3) or PUV-treated HIV-1 IIIB (data not shown) did not induce appreciable IFN-␥production in the PBMC. Thus, mDC were superior to iDC at inducing IFN-␥production in autologous PBMC. Moreover, HIV-1 IIIB-superinfected apo-ptotic cells were more potent immunogens in these mDC than were HIV-1 IIIB-superinfected necrotic cell preparations or cell-free HIV-1 IIIB.
To determine whether the T-cell responses were mediated by HLA class I- and class II-restricted, CD8⫹ and CD4⫹ T
cells, highly enriched autologous CD8⫹and CD4⫹T cells were
obtained by negative selection with immunomagnetic beads specific for either CD4 or CD8 and CD19 and CD16 (Dynal, Lake Success, N.Y.) (15). Both CD8⫹and CD4⫹T cells
pro-duced IFN-␥in response to the mDC⫹AP-IIIB (Fig. 4A). In further experiments, the addition of either anti-HLA class I MAb or anti-HLA class II MAb to the DC-antigen prepara-tions (14) inhibited induction of IFN-␥production by autolo-gous mDC⫹AP-IIIB; the addition of both of these MAb com-pletely prevented induction of IFN-␥in the PBMC (Fig. 4B). Therefore, mDC⫹AP-IIIB induced IFN-␥production in au-tologous CD8⫹and CD4⫹T cells that was HLA class I and
class II restricted, respectively.
We next expanded these studies by examining the ability of autologous, HIV-1-infected, apoptotic cell preparations to
in-duce IFN-␥in PBMC cultures from eight HIV-1-infected per-sons. The individual and composite data confirmed our find-ings that the greatest number of IFN-␥-producing cells was induced by mDC⫹AP-IIIB (Table 1). The highest levels of IFN-␥were stimulated by mDC⫹AP-IIIB in PBMC cultures from seven of the eight HIV-1-infected persons compared to mDC⫹AP. HIV-1-specific T-cell responses were not induced by HIV-1 antigen-loaded iDC or mDC preparations in PBMC from three HIV-1-negative subjects.
The one HIV-1-infected subject (S2) who did not have greater numbers of IFN-␥-producing cells induced by the mDC⫹AP-IIIB had the highest response to mDC⫹AP. This same pattern of T-cell response to the apoptotic cell prepara-tions was confirmed in PBMC obtained 6 months later from this subject (data not shown).
Interestingly, HIV-1 p24 was detected in all of the AP prep-arations, although the levels were lower than in the AP-IIIB preparations (Table 1). This represents endogenous HIV-1 from naturally infected cells of the study subjects and could have resulted in stimulation of a portion of the IFN-␥by the mDC⫹AP preparations. However, induction of IFN-␥in the PBMC cultures was not significantly different when mDC⫹AP were used for antigen presentation compared to mDC alone. We next determined whether the T-cell response to the apoptotic cell preparation of X4 virus was consistent over time. We found that there were comparable levels of IFN-␥induced by autologous mDC⫹AP-IIIB over a⬎2-year period in one subject (S5) (Fig. 5). Very low levels of IFN-␥were induced by mDC alone or mDC⫹AP. Persistent production of IFN-␥by PBMC stimulated with mDC⫹AP-IIIB was confirmed at two separate time points 6 months apart in subject S3 (data not shown).
The data presented in Table 1 show that the level IFN-␥ production was not related to the amount of HIV-1 p24 in the AP or AP-IIIB antigen preparations loaded in the DC. Further studies indicated that very low levels of p24 were maintained from the baseline level through 60 h of culture of iDC or mDC
FIG. 4. (A) Both CD8⫹and CD4⫹T-cell subpopulations produced IFN-␥after treatment with mDC⫹AP-IIIB (subject S6). (B) Both HLA
class I- and class II-associated, anti-HIV-1 IFN-␥responses were induced in PBMC by mDC⫹AP-IIIB (subject S5). These data are representative of five experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
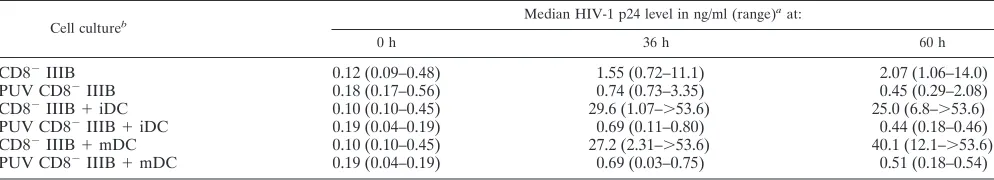
[image:5.587.67.519.78.258.2]that had been loaded with HIV-1 IIIB-infected, PUV-treated, apoptotic CD8⫺cell preparations (Table 2). These low levels
of p24 were similar to those in concurrent cultures of the HIV-1 IIIB-infected, PUV-treated, apoptotic CD8⫺cell
prep-arations without the DC. These results show that PUV treat-ment inactivates HIV-1 in the apoptotic CD8⫺cell
prepara-tions, with a resultant lack of replication of HIV-1 in DC loaded with them. However, both iDC and mDC loaded with non-PUV-treated, HIV-1 IIIB-infected CD8⫺cells supported
very high levels of replication of HIV-1 (Table 2). In contrast, iDC or mDC alone did not support efficient HIV-1 replication after infection with cell-free X4 virus (data not shown).
These results indicate that HIV-1-infected cells can serve as a source of antigen for efficient induction of HIV-1-specific CD8⫹ and CD4⫹ T cells by mDC from persons chronically
infected with HIV-1 who are receiving combination antiretro-viral therapy. This immunogenic property was promoted by driving the infected cells into apoptosis by PUV treatment in vitro. Other investigators have reported activation of T-cell responses specific for influenza A virus (2), human cytomega-lovirus (3), Epstein-Barr virus (31, 45), and cancer antigens (20, 22, 23, 25, 42) by DC loaded with antigen-expressing apoptotic cells. In our system, induction of programmed cell death may have led to enhanced recognition and uptake of the apoptotic cells by iDC (1). Cells infected with HIV-1 naturally undergo extensive apoptosis after more prolonged infection than that used in these studies (4). This could thus provide a
rich source of HIV-1 structural and nonstructural antigens for processing by iDC and presentation by mDC to CD8⫹and
CD4⫹T cells under conditions of natural infection.
Addition-ally, residual HIV-1-infected live cells in the AP-IIIB prepara-tions could be a source of antigen. In support of this, Harshyne et al. (19) have recently reported that living cells can transfer antigen to DC for activation of CD8⫹T cells. Further studies
are needed to compare the antigenicity of HIV-1-infected ap-optotic and viable cells in DC.
We found that HIV-1-infected necrotic cells were not a potent T-cell immunogen compared to HIV-1-infected apo-ptotic cells. This is in agreement with the findings of Albert et al. (2), who demonstrated that activation of influenza A virus-specific CD8⫹T cells by mDC loaded with influenza A
virus-infected apoptotic monocytes was superior to that induced by mDC containing virus-infected necrotic cell preparations. Moreover, the influenza virus antigen from the apoptotic cell preparations was processed through a nonclassical, alternative HLA class I pathway by the DC. The efficiency of presentation of antigens of HIV-1 and other viruses may reflect differences in the mechanisms of processing and presentation of apoptotic and necrotic cell preparations by DC. These are likely based on differences in viral proteins, such as the predominance of gly-cine-alanine repeats that can act on proteosomes (8), and differential activity of cellular components such as chaperone molecules that regulate antigen trafficking (29).
There was little or no HIV-1 replication demonstrable in either the iDC or the mDC that had been loaded with the X4 virus-infected, PUV-treated apoptotic CD8⫺cells. The mDC
loaded with this form of inactivated HIV-1 were, however, highly immunogenic for anti-HIV-1 CD8⫹T cells. This
sug-gests that the major processing of HIV-1 proteins in these DC for presentation to CD8⫹ T cells was not through a classic
HLA class I endogenous pathway. We hypothesize that, after the ingestion of HIV-1-infected apoptotic cells by the iDC, HIV-1 proteins in the apoptotic cells are processed through an alternative HLA class I pathway for cross-presentation by mDC to HIV-1-specific CD8⫹ T cells. HIV-1 peptides
pro-cessed by the infected CD8⫺cells may also serve as a source of
antigen for the mDC. Additional studies are required to ad-dress directly these hypotheses, including inhibitors of various steps in HIV-1 replication and processing of viral proteins by DC.
The iDC or mDC infected with replication-competent or PUV-treated cell-free HIV-1 IIIB did not activate an HIV-1-specific T-cell response in our study. Buseyne et al. (10)
[image:6.587.49.277.72.238.2]re-FIG. 5. T-cell responses to mDC⫹AP-IIIB are persistent over time in the same individual (subject S5). Cryopreserved PBMC were used as responder cells for this experiment.
TABLE 2. HIV-1 p24 levels in iDC and mDC loaded with untreated and PUV-treated, HIV-1 IIIB-infected CD8⫺autologous cells
Cell cultureb Median HIV-1 p24 level in ng/ml (range)
aat:
0 h 36 h 60 h
CD8⫺IIIB 0.12 (0.09–0.48) 1.55 (0.72–11.1) 2.07 (1.06–14.0)
PUV CD8⫺IIIB 0.18 (0.17–0.56) 0.74 (0.73–3.35) 0.45 (0.29–2.08)
CD8⫺IIIB⫹iDC 0.10 (0.10–0.45) 29.6 (1.07–⬎53.6) 25.0 (6.8–⬎53.6)
PUV CD8⫺IIIB⫹iDC 0.19 (0.04–0.19) 0.69 (0.11–0.80) 0.44 (0.18–0.46)
CD8⫺IIIB⫹mDC 0.10 (0.10–0.45) 27.2 (2.31–⬎53.6) 40.1 (12.1–⬎53.6)
PUV CD8⫺IIIB⫹mDC 0.19 (0.04–0.19) 0.69 (0.03–0.75) 0.51 (0.18–0.54)
aCD8⫺cells and DC were derived from PBMC of three normal, HIV-1-negative donors (N5, N6, and N7) and cultured for the designated times. bCD8⫺IIIB⫽HIV-1 IIIB-infected, autologous CD8⫺cells; PUV CD8⫺⫽PUV-treated, HIV-1 IIIB-infected, autologous CD8⫺cells.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.587.44.541.621.711.2]cently reported that iDC that had been infected with X4 virus could stimulate HIV-1-specific CD8⫹ T cells in vitro. This
required fusion and cytosolic processing of the virus by the iDC and was not related to endogenous virus replication in these cells. However, antigen presentation in that study was deter-mined by stimulation of HIV-1 peptide-specific CD8⫹T-cell
lines and clones by the virus-fused DC. Such cell lines and clones should have a much greater proportion of HIV-1-spe-cific CD8⫹T cells than the PBMC used in our work. Their
T-cell model may therefore detect lower amounts of HIV-1 peptide expressed in HLA class I molecules on the DC. Col-lectively, our studies suggest that DC have nonclassical, alter-native mechanisms for cross-presentation of X4 viral antigens to CD8⫹T cells.
Of importance is that maturation of DC induced by treat-ment with CD40L after loading of the iDC with the HIV-1 antigen preparations was essential for the efficient induction of anti-HIV-1 T-cell responses. This is likely related to enhanced expression of cell surface molecules CD80 and CD86 that are required for activation of T cells (6). CD40L also induces cytokines such as IL-12 (32) and IL-15 (28) that upregulate and prolong survival of antigen-specific T cells. In this regard, Os-trowski et al. (33) reported that CD40L treatment of HIV-1 peptide-treated DC enhances their activation of HIV-1-spe-cific cytotoxic T lymphocytes in vitro, which is associated with production of IL-15 by the DC. However, in terms of the natural HIV-1 infection, these immunity-enhancing effects of CD40L on DC should be viewed in the context of potential pathogenic effects due to upregulation of HIV-1 X4 replication in cocultures of CD40L-treated mDC and CD4⫹T cells (30).
Indeed, we found high levels of virus replication in both iDC and mDC cultures loaded with non-PUV-treated, HIV-1 IIIB-infected CD8⫺cells. Thus, there may be competing events of
uptake and presentation of HIV-1 X4-infected cells by mDC to T cells, with replication and cytopathic effects of this virus.
We (37) and others (26, 35, 36) have previously shown that CD8⫹T-cell reactivity to HIV-1 is only temporarily increased
in patients with long-term, chronic HIV-1 infection receiving combination antiretroviral drug therapy, even though the lev-els of circulating HIV-1 are greatly diminished. Additionally, there is usually poor recovery of anti-HIV-1 CD4⫹T-cell
ac-tivity in these persons (9, 12, 36, 38, 49). There do appear, however, to be residual T cells that are HIV-1 specific. This has been shown by reactivity of CD8⫹ T cells to HIV-1 during
treatment interruptions (16, 39, 41) and by in vitro stimulation with DC loaded with HIV-1 peptides (33) or proteins com-plexed with liposome (14). The present study indicates that HIV-1-specific CD8⫹T cells, as well as CD4⫹T cells, can be
readily activated to produce IFN-␥in vitro by a single round of stimulation with DC that have been loaded with autologous, HIV-1-infected apoptotic cells and matured with CD40L. This approach may be useful as a therapeutic vaccine to enhance T-cell immunity to a person’s own endogenous HIV-1 by au-tologous DC that have been loaded with apoptotic cells that contain strains of HIV-1 that are unique to the individual.
This work was supported in part by grants R01 AI41870, U01 AI37984, and U01 AI35041 from the National Institutes of Health.
We thank K. Picha of Immunex for assistance in providing the CD40L; S. Narula of Schering-Plough (Kenilworth, N.J.) for providing hIL-4 and hGM-CSF; W. Buchanan and B. Calhoun for clinical
assis-tance; R. Day, S. Barratt-Boyes, and L. Harshyne for technical advice; S. Alber, M. White, D. Damph, H. Li, C. Perfetti, C. Kalinyak, W. Jiang, M. Mather, and P. Zhang for technical assistance; and J. Malenka for administrative assistance. We give special thanks to the volunteers of the Pitt Men’s Study–MACS, whose dedication made this study possible.
REFERENCES
1.Albert, M. L., S. F. Pearce, L. M. Francisco, B. Sauter, P. Roy, R. L. Silverstein, and N. Bhardwaj.1998. Immature dendritic cells phagocytose apoptotic cells via␣v5and CD36, and cross-present antigens to cytotoxic T
lymphocytes. J. Exp. Med.188:1359–1368.
2.Albert, M. L., B. Sauter, and N. Bhardwaj.1998. Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature 392:86–89.
3.Arrode, G., C. Boccaccio, J. Lule, S. Allart, N. Moinard, J. P. Abastado, A. Alam, and C. Davrinche. 2000. Incoming human cytomegalovirus pp65 (UL83) contained in apoptotic infected fibroblasts is cross-presented to CD8⫹T cells by dendritic cells. J. Virol.74:10018–10024.
4.Badley, A. D., A. A. Pilon, A. Landay, and D. H. Lynch.2000. Mechanisms of HIV-associated lymphocyte apoptosis. Blood96:2951–2964.
5.Balachandran, R., P. Thampatty, A. Enrico, C. Rinaldo, and P. Gupta.1991. Human immunodeficiency virus isolates from asyptomatic homosexual men and from AIDS patients have distinct biologic and genetic properties. Vi-rology180:229–238.
6.Banchereau, J., F. Briere, C. Caux, J. Davoust, S. Lebecque, Y. J. Liu, B. Pulendran, and K. Palucka.2000. Immunobiology of dendritic cells. Annu. Rev. Immunol.18:767–811.
7.Bjorndal, A., H. Deng, M. Jansson, J. R. Fiore, C. Colognesi, A. Karlsson, J. Albert, G. Scarlatti, D. R. Littman, and E. M. Fenyo.1997. Coreceptor usage of primary human immunodeficiency virus type 1 isolates varies according to biological phenotype. J. Virol.71:7478–7487.
8.Blake, N., S. Lee, I. Redchenko, W. Thomas, N. Steven, A. Leese, P. Steiger-wald-Mullen, M. G. Kurilla, L. Frappier, and A. Rickinson.1997. Human CD8⫹T-cell responses to EBV EBNA1: HLA class I presentation of the (Gly-Ala)-containing protein requires exogenous processing. Immunity 7:791–802.
9.Blankson, J. N., J. E. Gallant, and R. F. Siliciano.2001. Proliferative re-sponses to human immunodeficiency virus type 1 (1) antigens in HIV-1-infected patients with immune reconstitution. J. Infect. Dis.183:657–661. 10.Buseyne, F., S. Le Gall, C. Boccaccio, J. P. Abastado, J. D. Lifson, L. O. Arthur, Y. Riviere, J. M. Heard, and O. Schwartz.2001. MHC-I-restricted presentation of HIV-1 virion antigens without viral replication. Nat. Med. 7:344–349.
11.Canque, B., Y. Bakri, S. Camus, M. Yagello, A. Benjouad, and J. C. Gluck-man.1999. The susceptibility to X4 and R5 human immunodeficiency virus-1 strains of dendritic cells derived in vitro from CD34⫹hematopoietic
pro-genitor cells is primarily determined by their maturation stage. Blood93: 3866–3875.
12.Carcelain, G., P. Debre, and B. Autran.2001. Reconstitution of CD4⫹T
lymphocytes in HIV-infected individuals following antiretroviral therapy. Curr. Opin. Immunol.13:483–488.
13.Chun, T. W., R. T. Davey, Jr., M. Ostrowski, J. Shawn Justement, D. Engel, J. I. Mullins, and A. S. Fauci.2000. Relationship between pre-existing viral reservoirs and the re-emergence of plasma viremia after discontinuation of highly active antiretroviral therapy. Nat. Med.6:757–761.
14.Fan, Z., Huang, X. L., Borowski, L., Mellors, J. W., and Rinaldo, C. R., Jr. 2001. Restoration of anti-human immunodeficiency virus type 1 (HIV-1) responses in CD8⫹T cells from late-stage patients on prolonged
antiretro-viral therapy by stimulation in vitro with HIV-1 protein-loaded dendritic cells. J. Virol.75:4413–4419.
15.Gallucci, S., M. Lolkema, and P. Matzinger.1999. Natural adjuvants: en-dogenous activators of dendritic cells. Nat. Med.5:1249–1255.
16.Garcia, F., M. Plana, G. M. Ortiz, S. Bonhoeffer, A. Soriano, C. Vidal, A. Cruceta, M. Arnedo, C. Gil, G. Pantaleo, T. Pumarola, T. Gallart, D. F. Nixon, J. M. Miro, and J. M. Gatell.2001. The virological and immunolog-ical consequences of structured treatment interruptions in chronic HIV-1 infection. AIDS15:F29–F40.
17.Granelli-Piperno, A., E. Delgado, V. Finkel, W. Paxton, and R. M. Steinman. 1998. Immature dendritic cells selectively replicate macrophage-tropic (M-tropic) human immunodeficiency virus type 1, while mature cells efficiently transmit both M- and T-tropic virus to T cells. J. Virol.72:2733–2737. 18.Gulick, R. M., J. W. Mellors, D. Havlir, J. J. Eron, A. Meibohm, J. H.
Condra, F. T. Valentine, D. McMahon, C. Gonzalez, L. Jonas, E. A. Emini, J. A. Chodakewitz, R. Isaacs, and D. D. Richman.2000. 3-Year suppression of HIV viremia with indinavir, zidovudine, and lamivudine. Ann. Intern. Med.133:35–39.
19.Harshyne, L. A., S. C. Watkins, A. Gambotto, and S. M. Barratt-Boyes.2001. Dendritic cells acquire antigens from live cells for cross-presentation to CTL. J. Immunol.166:3717–3723.
on November 8, 2019 by guest
http://jvi.asm.org/
20.Heath, W. R., and F. R. Carbone.2001. Cross-presentation, dendritic cells, tolerance and immunity. Annu. Rev. Immunol.19:47–64.
21.Herr, W., E. Ranieri, W. Olson, H. Zarour, L. Gesualdo, and W. J. Storkus. 2000. Mature dendritic cells pulsed with freeze-thaw cell lysates define an effective in vitro vaccine designed to elicit EBV-specific CD4⫹and CD8⫹T
lymphocyte responses. Blood96:1857–1864.
22.Hoffmann, T. K., N. Meidenbauer, G. Dworacki, H. Kanaya, and T. L. Whiteside.2000. Generation of tumor-specific T-lymphocytes by cross-prim-ing with human dendritic cells cross-prim-ingestcross-prim-ing apoptotic tumor cells. Cancer Res. 60:3542–3549.
23.Hoffmann, T. K., N. Meidenbauer, J. Muller-Berghaus, W. J. Storkus, and T. L. Whiteside.2001. Proinflammatory cytokines and CD40 ligand enhance cross-presentation and cross-priming capability of human dendritic cells internalizing apoptotic cancer cells. J. Immunother.24:162–171.
24.Ignatius, R., M. Marovich, E. Mehlhop, L. Villamide, K. Mahnke, W. I. Cox, F. Isdell, S. S. Frankel, J. R. Mascola, R. M. Steinman, and M. Pope.2000. Canarypox virus-induced maturation of dendritic cells is mediated by apo-ptotic cell death and tumor necrosis factor alpha secretion. J. Virol.74: 11329–11338.
25.Jenne, L., J. F. Arrighi, J. Jonuleit, J. H. Saurat, and C. Hauser.2000. Dendritic cells containing apoptotic melanoma cells prime human CD8⫹T
cells for efficient tumor cell lysis. Cancer Res.60:4446–4452.
26.Kalams, S. A., P. J. Goulder, A. K. Shea, N. G. Jones, A. K. Trocha, G. S. Ogg, and B. D. Walker.1999. Levels of human immunodeficiency virus type 1-specific cytotoxic T-lymphocyte effector and memory responses decline after suppression of viremia with highly active antiretroviral therapy. J. Virol. 73:6721–6728.
27.Koot, M., I. P. Keet, A. H. Vos, R. E. de Goede, M. T. Roos, R. A. Coutinho, F. Miedema, P. T. Schellekens, and M. Tersmette.1993. Prognostic value of HIV-1 syncytium-inducing phenotype for rate of CD4⫹cell depletion and
progression to AIDS. Ann. Intern. Med.118:681–688.
28.Kuniyoshi, J. S., C. J. Kuniyoshi, A. M. Lim, F. Y. Wang, E. R. Bade, R. Lau, E. K. Thomas, and J. S. Weber.1999. Dendritic cell secretion of IL-15 is induced by recombinant huCD40LT and augments the stimulation of anti-gen-specific cytolytic T cells. Cell. Immunol.193:48–58.
29.Larsson, M., J. F. Fonteneau, and N. Bhardwaj.2001. Dendritic cells res-urrect antigens from dead cells. Trends Immunol.22:141–148.
30.McDyer, J. F., M. Dybul, T. J. Goletz, A. L. Kinter, E. K. Thomas, J. A. Berzofsky, A. S. Fauci, and R. A. Seder.1999. Differential effects of CD40 ligand/trimer stimulation on the ability of dendritic cells to replicate and transmit HIV infection: evidence for CC-chemokine-dependent and -inde-pendent mechanisms. J. Immunol.162:3711–3717.
31.Metes, D., W. Storkus, A. Zeevi, K. Patterson, A. Logar, D. Rowe, M. A. Nalesnik, J. J. Fung, and A. S. Rao.2000. Ex vivo generation of effective Epstein-Barr virus (EBV)-specific CD8⫹cytotoxic T lymphocytes from the
peripheral blood of immunocompetent Epstein Barr virus-seronegative in-dividuals. Transplantation70:1507–1515.
32.Mosca, P. J., A. C. Hobeika, T. M. Clay, S. K. Nair, E. K. Thomas, M. A. Morse, and H. K. Lyerly.2000. A subset of human monocyte-derived den-dritic cells expresses high levels of interleukin-12 in response to combined CD40 ligand and interferon-gamma treatment. Blood96:3499–3504. 33.Ostrowski, M. A., S. J. Justement, L. Ehler, S. B. Mizell, S. Lui, J. Mican,
B. D. Walker, E. K. Thomas, R. Seder, and A. S. Fauci.2000. The role of CD4⫹T-cell help and CD40 ligand in the in vitro expansion of
HIV-1-specific memory cytotoxic CD8⫹T-cell responses. J. Immunol.165:6133–
6141.
34.Pamer, E., and Cresswell, P.1998. Mechanisms of MHC class I-restricted antigen processing. Annu. Rev. Immunol.16:323–358.
35.Pontesilli, O., S. Kerkhof-Garde, D. W. Notermans, N. A. Foudraine, M. T. Roos, M. R. Klein, S. A. Danner, J. M. Lange, and F. Miedema.1999. Functional T cell reconstitution and human immunodeficiency
virus-1-spe-cific cell-mediated immunity during highly active antiretroviral therapy. J. In-fect. Dis.180:76–86.
36.Richman, D. D.2001. HIV chemotherapy. Nature410:995–1001. 37.Rinaldo, C. R., Jr., X. L. Huang, Z. Fan, J. B. Margolick, L. Borowski, A.
Hoji, C. Kalinyak, D. K. McMahon, S. A. Riddler, W. H. Hildebrand, R. B. Day, and J. W. Mellors.2000. Anti-human immunodeficiency virus type 1 (HIV-1) CD8⫹T-lymphocyte reactivity during combination antiretroviral
therapy in HIV-1-infected patients with advanced immunodeficiency. J. Vi-rol.74:4127–4138.
38.Rinaldo, C. R., Jr., J. M. Liebmann, X. L. Huang, Z. Fan, Q. Al-Shboul, D. K. McMahon, R. D. Day, S. A. Riddler, and J. W. Mellors.1999. Prolonged suppression of human immunodeficiency virus type 1 (HIV-1) viremia in persons with advanced disease results in enhancement of CD4 T cell reac-tivity to microbial antigens but not to HIV-1 antigens. J. Infect. Dis.179: 329–336.
39.Rosenberg, E. S., M. Altfeld, S. H. Poon, M. N. Phillips, B. M. Wilkes, R. L. Eldridge, G. K. Robbins, R. T. D’Aquila, P. J. Goulder, and B. D. Walker. 2000. Immune control of HIV-1 after early treatment of acute infection. Nature407:523–526.
40.Rovere, P., C. Vallinoto, A. Bondanza, M. C. Crosti, M. Rescigno, P. Ric-ciardi-Castagnoli, C. Rugarli, and A. A. Manfredi.1998. Bystander apoptosis triggers dendritic cell maturation and antigen-presenting function. J. Immu-nol.161:4467–4471.
41.Ruiz, L., G. Carcelain, J. Martinez-Picado, S. Frost, S. Marfil, R. Paredes, J. Romeu, E. Ferrer, K. Morales-Lopetegi, B. Autran, and B. Clotet.2001. HIV dynamics and T-cell immunity after three structured treatment inter-ruptions in chronic HIV-1 infection. AIDS15:F19–F27.
42.Russo, V., S. Tanzarella, P. Dalerba, D. Rigatti, P. Rovere, A. Villa, C. Bordignon, and C. Traversari.2000. Dendritic cells acquire the MAGE-3 human tumor antigen from apoptotic cells and induce a class I-restricted T cell response. Proc. Natl. Acad. Sci. USA97:2185–2190.
43.Schuitemaker, H., M. Koot, N. A. Kootstra, M. W. Dercksen, R. E. de Goede, R. P. van Steenwijk, J. M. Lange, J. K. Schattenkerk, F. Miedema, and M. Tersmette.1992. Biological phenotype of human immunodeficiency virus type 1 clones at different stages of infection: progression of disease is asso-ciated with a shift from monocytotropic to T-cell-tropic virus population. J. Virol.66:1354–1360.
44.Shankarappa, R., J. B. Margolick, S. J. Gange, A. G. Rodrigo, D. Upchurch, H. Farzadegan, P. Gupta, C. R. Rinaldo, G. H. Learn, X. He, X. L. Huang, and J. I. Mullins.1999. Consistent viral evolutionary changes associated with the progression of human immunodeficiency virus type 1 infection. J. Virol. 73:10489–10502.
45.Subklewe, M., C. Paludan, M. L. Tsang, K. Mahnke, R. M. Steinman, and C. Munz.2001. Dendritic cells cross-present latency gene products from Ep-stein-Barr virus-transformed B cells and expand tumor-reactive CD8⫹killer
T cells. J. Exp. Med.193:405–411.
46.Tabi, Z., M. Moutaftsi, and L. K. Borysiewicz.2001. Human cytomegalovirus pp65- and immediate early 1 antigen-specific HLA class I-restricted cytotoxic T cell responses induced by cross-presentation of viral antigens. J. Immunol. 166:5695–5703.
47.van Kooten, C., and J. Banchereau.1997. Functional role of CD40 and its ligand. Int. Arch. Allergy Immunol.113:393–399.
48.Walker, B. D., E. S. Rosenberg, C. M. Hay, N. Basgoz, and O. O. Yang.1998. Immune control of HIV-1 replication. Adv. Exp. Med. Biol.452:159–167. 49.Watanabe, N., M. Tomizawa, A. Tachikawa-Kawana, M. Goto, A. Ajisawa, T.
Nakamura, and A. Iwamoto.2001. Quantitative and qualitative abnormali-ties in HIV-1-specific T cells. AIDS15:711–715.
50.Yewdell, J. W., C. C. Norbury, and J. R. Bennink.1999. Mechanisms of exogenous antigen presentation by MHC class I molecules in vitro and in vivo: implications for generating CD8⫹T cell responses to infectious agents,
tumors, transplants, and vaccines. Adv. Immunol.73:1–77.