Copyright©1976 AmericanSocietyforMicrobiology PrintedinU-SA.
Synthesis
of the
Adenovirus-Coded
DNA
Binding
Protein
in
Infected
Cells
ZVEE GILEAD,* KENJI SUGAWARA, G. SHANMUGAM, AND MAURICE GREEN
Institute for MolecularVirology,St. LouisUniversity School ofMedicine,St.Louis,Missouri63110
Receivedfor publication 25 November 1975
Synthesis of the 75K (75K indicates a molecularweight of70,000to 75,000)
DNAbinding protein, an early virus-coded proteininadenovirus 2-infected KB
cells, and its regulation were studied by using a radioimmune precipitation
inhibition assay. The protein was first detected at 4 h postinfection and
accumu-lated at an exponential rate. An arrest of further synthesis (accumulation)was
observed at 10 to 11 hpostinfection,coincidingwiththeonsetofsynthesisoflate
virion proteins. In contrast, when the infected cells were treated with25 ug of
arabinosyl cytosine per ml toblockviral DNAreplication, thesynthesisof 75K
protein did not cease but continuedfor up to 36 h postinfection. Thesynthesis of
75K protein incellsafter release from acycloheximide block (2to 9h
postinfec-tion) wasanalyzed. Increased amounts ofearlyadenovirus-specificmRNA
accu-mulate in infected cellsduringacycloheximideblock(Parsons andGreen,1971).
However, cycloheximide treatment did not produce increased levels of75K
pro-tein, and an abrupt arrest of 75K protein formation was again observed at the
time ofsynthesis of late virion proteins. Partition of the 75K protein between the
nuclear and cytoplasmic fractions during the course of infection was studied.
The 75K proteinappeared first in the cytoplasm andthen in the nucleus after a
slight lag.Accumulation of the 75K protein continued both in thecytoplasm and
nucleus, with higherlevelsbeingfoundinthe cytoplasm.
Several virus-specific polypeptides that are
not viron components are synthesized in
hu-mancells infected with adenovirus 2 or 5(Ad2,
Ad5). These new polypeptides were identified
by electrophoresis on sodium dodecyl
sulfate-polyacrylamidegels of extracts ofinfectedcells pulse labeled at early times after infection (1, 2,
9, 14, 15, 20).They range in size from 75,000 to
10,000daltons, based on their migration in gels.
Only one of these early polypeptides with a
molecular weight of 70,000 to 75,000 (referred to
as 75K protein) has been purified to date. It
binds to single-stranded but not double-stranded DNA and has been termed the "DNA
binding protein" (17, 18). The 75K proteins
codedby Ad2 andAd5 have similarproperties
andmolecularweights (13). Use of Ad5
temper-ature-sensitive mutants has indicated that the
75Kprotein is virus coded and isnecessary for
viral DNAreplication (19). The viral gene
cod-ing for ithas been mapped, by the use of
tem-perature-sensitive mutants, in the EcoRl B
fragment of the adenovirus genome (T.
Grod-zicker, C. W. Anderson, and J. Sambrook,
personal communication). Another polypeptide
ofmolecular weight 45,000 elutes from
DNA-cellulosecolumns together with the 75K protein
and appearstobeadegradation product of the
75Kprotein (13).
We have recently shown by radioimmune
precipitation and radioimmune precipitation
inhibition (RIPI) that the Ad2 75K protein is
virus specific (5). The method involved theuse
ofpurified 3H-labeled 75K protein and its
ho-mologous antibody present in sera from
ham-sters immunized with extracts of Ad2-simian
virus 40-induced tumors (4, 5). We have now
developed the RIPI method for the accurate
quantitation of the 75K protein in order to
study its synthesis. The presentcommunication
describes ourfindingson the synthesis of this
proteinduring infection and its modulationby
interferencewithDNAorprotein
syntheses.
MATERIALS AND METHODS
Isolation of labeled 75K protein. Labeled 75K protein was isolated from Ad2-infected KB cells and purified by DNA-cellulosechromatographyas previ-ously described (5, 17) withtwomodifications. We foundthat thetrailingedgeofthe0.6Mpeak elut-ing from the DNA-cellulose column contained only 75Kprotein, whereas the leading edge of the peak contained also the 45Kprotein. Fractions from the trailing edge of the peak only were pooled and used asthesourceof75K protein for the RIPI assay. In 454
on November 10, 2019 by guest
http://jvi.asm.org/
SYNTHESIS OF DNA BINDING PROTEIN 455
addition, labeling was performed with [35S]methio-nine instead of [3H]leucine, yielding preparations with higher specific radioactivity. Infected cells were incubated with 5 ,uCi of [35S]methionine per ml (210 to 466 Ci/mmol) in methionine-free Eagle minimal essential medium containing 5% horse se-rum.M5S-labeled75K protein wasprepared monthly tominimizeradioactive decay.
Experimental infection of KB cells and prepara-tion of cell extracts.Suspension cultures ofKBcells
wereinfected with purified Ad2 at a multiplicity of 200 PFU/cell aspreviouslydescribed (17). Infected
cellsweremaintainedat aconcentration of 3 x 105 cells/ml in Eagle minimal essential medium con-taining 5% horseserum. To prepare whole-cell ex-tractsfor assay of 75K protein, 9 x 106cells (30 ml) werewithdrawnatspecifiedtimesand washed with Ca2+- and Mg2+-free phosphate-buffered saline (PBS). The cells wereresuspended in 1 ml of PBS containing 1% Triton X-100 and stored at -5C. Before assay, suspensions weredisruptedfor 3 min atfull power inaRaytheon DF-101 sonicoscillator and clarified at 2,000 x g for 10 min at4C. The supernatant fractionswerecollected for assay. For the preparation ofcytoplasmicandnuclear extracts,
washed samplesof 9x 106 cellswereresuspendedin 1 ml ofPBS containing 1% Triton X-100 at room temperature, and after5minnuclei andcytoplasm
were separated by centrifugationat800 xgfor 10 min. Thenucleiweresonicated and clarifiedas de-scribed for whole-cellextracts.
To prepare extractsforcomplement fixation tests, 10-ml samplesofinfected cells (3 x 106cells)were washed withPBS,resuspendedin0.5mlof veronal buffer (16),sonicated for3min,and clarifiedat2,000
x g for 10 min. The supernatant fractions were collected for assay.
RIPI. The double-antibody method for radioim-mune precipitation oflabeledpurified 75Kprotein
and thesourceof antisera have beendescribed (5).
Inthe RIPI assay, 10-,ugaliquotsof immune ham-ster immunoglobulin G (IgG) (supplemented with 70 ,ugof "carrier" normal hamsterIgG)were
prein-cubated with various amountsofextracts of unla-beled infected cellssampledatvarioustimes post-infectionto blockantibody activity. Preincubation wasfor 1 h at 37 C in PBS containing 1% Triton X-100 (vol/vol) and 1% sodium deoxycholate (wt/
vol) inconical 0.75-ml centrifuge tubes (Sarstedt,
Princeton, N.J.).The final reaction volumewas200
A1l.
Labeled 75K protein (10,000 counts/min) was thenadded, and incubation wascontinued for1 h. Finally, 30Al of goatserum antihamsterIgG wasadded, and theprecipitatesformed after2 h of in-cubation at 37 C were centrifuged, washed, and counted as previously described (5). Background
precipitationvalues(obtained with80 Agof normal hamster IgG) were subtracted in each determina-tion. The degree of inhibition observed with ex-tractsof unlabeled infected cellswascalculated as percentage of controlby comparisonwith reactions substitutingPBSfor cellextracts.
Titration of virionproteinsin extractsof infected cells by the complement fixation. The
micro-com-plement fixation testofSever (16) was used for the titration of virion proteins as previously described (6). Eight units of goat antibody to purified Ad2 virions wasused with 2 U of complement.
RESULTS
Quantitation of75K protein in extracts of
infected cells by the RIPI assay. The RIPI
assay is one of the few methods available to
accurately measure early proteins present in relatively low concentrations in infected cells.
Wehave developed a RIPI assay for
quantitat-ing intracellular 75K protein usquantitat-ing 35S-labeled
75K proteinanditshomologousantibody from
hamsters immunized with extracts of tumors
induced by an Ad2-simian virus 40 hybrid. To
standardizethe assay, a precipitation reaction
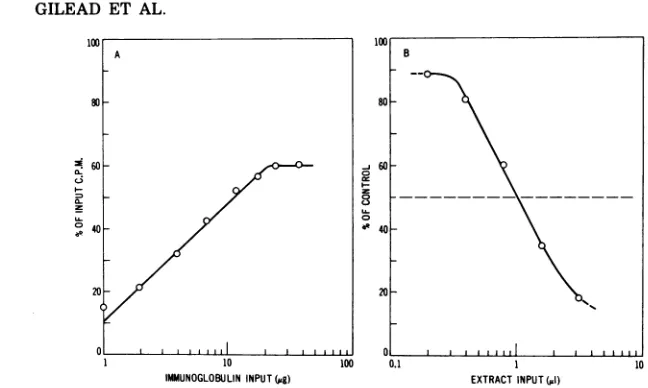
was performed first with increasing amounts of immune hamster IgG and 10,000 counts/min of 35S-labeled 75K protein (Fig. 1A). Ten
micro-grams of immune IgG, which precipitated.
about one-half of the 10,000-counts/min input
(Fig. 1A), was used subsequently in the RIPI
assays as follows.Aliquots of 10 ugof immune
IgG, supplemented with 70 ,ug of normalIgG,
were preincubated withincreasingvolumes of
unlabeled infected cellextractfor 1h at37 C.
35S-labeled 75K protein (10,000 counts/min)was
then added, thereaction mixtures werefurther
incubated, and the assay wascontinued as
de-scribed inMaterials and Methods. A represent-ative inhibition curve obtained with different
volumesof a 20-h infected cell extract is shown
in Fig. 1B. In this instance, 50% inhibition of
radioimmune precipitation was obtained with
1.03 ,tl of cell extract; the volume of extract
causing50%inhibition isdesignated
Vw.
Quan-titation ofthe 75K protein content in cell
ex-tracts is based upon the determination ofV50
from curves obtained for the individual ex-tracts, as describedin Fig. 1B, using different extractvolumes. Since the amount of 75K
pro-teinpresent in the extract isinverselyrelated
to V50, the 75K protein titer is therefore
ex-pressedininhibitoryunits as1/V50 x 100;
mul-tiplication by 100 is used to avoid fractional
values.
The fidelity ofthe RIPI assay was tested by
reconstruction reactions with known amounts
of unlabeled 75K protein. The 0.6 M eluate,
from a DNA-cellulose column, which is the source for labeled 75K protein, contains also
unlabeled hostcomponents (Sugawara, Gilead,
and Green, unpublished data). Unlabeled 75K
protein was therefore purified from the 0.6 M eluatetohomogeneity (Sugawara, Gilead, and
Green, manuscript in preparation). Known
amounts of pure 75Kproteinweremixed with
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
456 GILEAD ET AL.
~60 j60
0- L 8nVX _
040
"40-20 20
0 0 ii
1 10 100 0.1 1 10
IMMUNOGLOBULININPUT(,g) EXTRACTINPUT(,1)
FIG. 1. Quantitation of the RIPI assay. (A) Standard precipitation curve of35S-labeled 75Kprotein.
Aliquots of 75K protein (10,000counts/min) weremixedwithincreasing amountsofimmunoglobulin from
immunized hamsters (immuneIgG),supplemented withIgGfromnormal hamsters(normalIgG)togive 80
pgoftotal IgG, in reaction mixtures containing PBS, 1 % Triton X-100, and 1% sodiumdeoxycholate.After1h of incubation at 37 C, 30 4 of goat serum anti-hamster IgG was added and precipitates formed after 2 hof
incubation at 37 C werecentrifuged, washed twice with PBS containing 1% Triton X-100 and 1% sodium deoxycholate, dissolved in 0.25 M acetic acid, and counted in a scintillation spectrometer. Background radioactivity due tononspecificprecipitation, measured with 80pgofnormalIgGalone, was subtracted. (B) The RIPIassay. Increasing volumes of a 20-h infected cell extract were preincubated with 70pgof "carrier" normal IgG and 10pgof immuneIgG, shown in (A) to precipitate 50% of the10,000-counts/mininputof 35S-labeled 75K protein, for 1 h at 37 C in the buffer described in (A).35S-labeled75K protein (10,000counts/min)
wasadded, and incubation was continued for an additional 1 h at 37 C. Goat anti-hamster IgG (30pd) was
added,andthe precipitates formed after2 h at37 C weretreated wasdescribedin(A).Thebackground counts per minuteprecipitated with 80pgofnormal IgG was subtracted, and the percentage ofcontrol was calculated by comparison with a reactionmixtureinwhichPBSwassubstituted for the infected-cell extract.
extractsofuninfected KB cells (9x 106cells/ml) preparedasdescribed for infected cells. Aplot
of the concentration of 75K protein in the
ex-tracts versus the inhibitory titer measured in
the RIPItest (1/V5,, x 100) waslinear (Fig. 2), even at low 75K protein concentrations. The smallest detectable titer of 75Kprotein in in-fected cell extract is 0.5 inhibitory units. One unitcorrespondstoaconcentration of 1.4 Ag of
75Kprotein perml ofextract, accordingtothe
plotinFig. 2. However,wehavenotusedthis
plotto convertthe relativeinhibitorytiters into actual 75Kprotein concentrationsinthispaper
sincethe reconstructionmixturesarenot
iden-tical to extracts ofinfectedcells. Purified 75K
protein showsatendencyto aggregateandmay
be more prone to inactivation in the
recon-structedextractsthanintracellular75Kprotein
in extractsofinfectedcells.
Kinetics of synthesis of 75K protein in
in-fectedcells intheabsenceandpresenceof Ara
C. The 75Kprotein isanearlyprotein essential
for viral DNAreplication(19). Itissynthesized
beforetheonsetofviral DNAreplication, and also when viral DNA synthesis is inhibited
with arabinosyl cytosine (Ara C) (17, 18). The kinetics ofaccumulation ofthe 75K protein in
infected KB cellswereexamined(Fig. 3).Itwas
first detected at4hpostinfection, and its
syn-thesis (accumulation)wasexponentialupto 11
hpostinfection,atwhich time itstopped.There
was aslowdecline in theamountof 75Kprotein
after 11h, perhapsduetolysisofsomecellslate
duringproductive infectionortoinactivation of
75Kprotein molecules in vivo. In contrast, the initial rate ofaccumulation of 75K protein in Ara C-treated cellswas much slower, but did
notexhibitthe arrest observedinuninhibited
cultures (Fig. 3). The synthesis of 75K protein continued up to 36 h postinfection, at which time Ara C-treated cells accumulated four timesmore 75K protein than thehighestlevel
observed inuninhibitedcultures.
Synthesis of 75K protein in cells after
release fromacycloheximide block and effect of Ara C. Cells treated early after infection
with cycloheximide do not synthesize viral
DNA (18) and accumulate upto five times as
much early viral mRNAasuntreated infected
cells (12). It was of interest to determine whetherincreasedlevels of viral mRNA would result in an accelerated accumulation of 75K
protein after release from inhibition. Two pairs
of cultures, A and B, were infected. One A
A
1O
80
B
Inn, 1 rwn
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.79.403.56.250.2]SYNTHESIS OF DNA BINDING PROTEIN 457
Z50
200
C-,
I-L&J
-9 150
1oo[
50
n
140 120 100 80 60 40 20 0
[image:4.503.49.243.58.317.2]UNITS(1/V5ox100)
FIG. 2. Linearity of RIPI assay. A series of ex-tracts containing known concentrations of 75K pro-tein was prepared by adding different amounts of
purified unlabeled75Kprotein to an extract of
unin-fectedKBcells (9 x106 cells per ml of extract). The
reconstructedextracts were assayed by the RIPI as-say.The inhibitory titermeasuredinthe assay(1/VW
x100) was plotted against 75K proteinconcentration
(micrograms per milliliter of extract).
culturewastreated withcycloheximide(25
Ag/
ml) at2hpostinfection, and the other A culture
servedascontrol. At 9hpostinfection,both A
cultureswerewashedtwicewithwarmmedium
and incubatedinfresh medium lacking
inhibi-tor. One B culture received25 ,ugof
cyclohexi-mide per ml at 2 h postinfection, while the
other Bculture received25
,g
of Ara C per mlat 1 h postinfection. Both B cultures were
washed twice at 9 h postinfection with warm
mediumcontaining25
jig
of Ara C per ml andincubatedinmediumcontaining25 ug of AraC
per ml. Samples of 30 ml (9 x 106 cells) were
removedfrom AandBculturepairsthroughout
theexperiment foranalysisof 75Kprotein. The
A cultures (no Ara C) were sampled also for
assayof late virionproteinsbythecomplement
fixation test. The kinetics ofaccumulation of
75Kproteinin AculturesarepresentedinFig.
4A.Thesynthesis of75Kprotein inthecontrol
Aculture (not treatedwithcycloheximide)
ex-hibitedagain thearrestof 75Kprotein
synthe-sis.Afterreleasefrominhibition,the
cyclohexi-mide-treatedAculture exhibited a morerapid
rateofaccumulation of the75Kproteinas
com-pared with the control A culture. However, a
somewhatfaster onsetofarrestwas
observed,
resulting in a lower final titer. The time of
arrest in both A cultures coincided with the
onsetof synthesis of late virion proteins,
meas-ured by CF. Incontrast, noarrest insynthesis
of the 75K protein wasobserved in the B
cul-ture released from cycloheximide and then
treated with Ara C immediately after release.
Again, the released culture exhibited a
some-whatfasterinitial rate ofaccumulation as
com-paredwith theparallelBculture thatwas not
treated withcycloheximide.
Partition of the75K protein between
cyto-plasm and nucleus. The cytoplasmic fraction
from Ara C-treated infected cells harvestedat
24hpostinfection isroutinely used for the
puri-fication of the 75K protein. Unlike the nuclear
fraction, chromatography of the cytoplasmic
fraction on a DNA-cellulose columnyields
al-most pure labeled 75K protein (17). It was of
interest todetermine the kinetics of synthesis
and the partition of 75K proteininthe
cytoplas-mic and nuclear fractions. Infected cells were
sampled at various times after infection, and
10000
100
HOURS POSTINFECTION
FIG. 3. Synthesis of 75K protein inAd2-infected KB cells in thepresence and absence of Ara C. A suspension culture ofKB cells was infected with 200 PFUof Ad2per cell, divided into two portions, and maintained at 300,000 cellslml in Eagle minimal essential mediumcontaining5%horseserum. Ara C wasadded tofinal concentration of 25pg/mlat 1 h postinfection to one culture. Samples (30 ml) were removed at the indicated times,centrifuged,washed withPBS,andresuspendedin1mlofPBS contain-ing 1% Triton X-100. Cell suspensions were
soni-cated,clarified,andassayedfor75Kprotein bythe RIPIassay. Symbols: 0, infected culture,nodrug;
*,infected culture treated withAraC.
XrX
U- E 2
VOL. 18, 1976
I I A I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.257.446.317.543.2]458 GILEAD ET AL.
Z;
I-m
24 0 T
HOURS POST INFECTION
FIG. 4. Synthesis of 75K protein and virion proteins in Ad2-infected KB cells after release from a
cycloheximideblock in the absence(A)andthe presence(B)ofAraC.(A)AsuspensioncultureofKB cells was
infectedand divided into two. One culture received 25pg ofcycloheximidepermlat2hpostinfection,and the other culture served as control. Both cultures werecentrifugedat 9hpostinfection, washedtwicewithwarm
medium,andfurtherincubated at 37 C.Samples (30ml) wereremovedfromeach cultureatthe indicated times, and extracts wereprepared and assayed for 75K protein content by the RIPI assay. Ten-milliliter samples were removedalso for virion protein assay by the complement fixation test. (B) A suspension culture
ofKBcells wasinfected and divided into two portions. One culture received 25 pgof Ara C per mlat1 h postinfection, and thesecond received 25pgofcycloheximide per ml at 2 h postinfection. Both cultures were
centrifugedat 9 hpostinfection, washed twice with warm medium containing 25 pgofAraC perml,and further incubated in medium containing 25 pg ofAraC per ml.Samples(30ml) were removed andextracts were prepared and assayed for 75K protein by the RIPI assay. 75K protein titer: (A) 0, No drug; 0,
cycloheximide treated (2 to 9 h). (B)0,Ara C (1 to 24 h); *, cycloheximide (2 to 9 h), Ara C (9 to24h).Virion proteintiter: A, Nodrugs; A,cycloheximide treated.
thecytoplasmic andnuclear fractionswere
pre-pared as described in Materials and Methods.
Theaccumulation of 75K proteinincytoplasmic
and nuclear fractions of untreated infected cells
(Fig. 5A) and Ara C-treated cells (Fig. 5B)
showed the sameoverall kinetics observed for
whole cells (Fig. 3). Therewas somelaginthe
appearanceof the 75Kproteininthe nucleusas
compared with thecytoplasm, and theamount
of 75K inthe cytoplasmalways exceeded that
present inthenuclearfraction, except for late
timesinAra C-treated cultures.
DISCUSSION
Several investigations have identified early
and lateadenovirus-specific polypeptides in
in-fected human cells by using sodium dodecyl
sulfate-polyacrylamide gel electrophoresis of
extracts of infected cells labeled with [35S]methionine (1,2, 9,14, 15,20).Early
virus-specific protein synthesiswasdetectedstarting
at2.5 to4hpostinfection, andmaximum rates
ofsynthesis occurred at 6 to 12hpostinfection.
A gradual cessation was observed at later
times,anda"switch-over"tolate virionprotein
synthesis occurred at 12 to 16 hpostinfection.
The electrophoretic study of 75K protein,
termed ICSP3 or
E,
by Russel etal. (14, 15) orMaizel etal. (9, 20), respectively, was
compli-cated by the presence ofa host protein band
thatmigratedclosetothe 75K protein band(9,
20). The present study, measuring 75K protein
synthesis by a different method, is consistent
with the general pattern of early adenovirus
proteinsynthesis thatwaspreviously observed
by gel analyses. The methodusedin ourstudy
hastheadvantagethat it accurately measures
thenetaccumulation ofimmunologically active
proteinmolecules andis notinfluenced by
pos-sible variations in the equilibration and
con-centration of radioactiveamino acid pools and
by the background ofsimilarlymigratinghost
proteins. Byimmunological assay, we observed
anearly synthesis of 75K protein that is
appar-ently influenced by later events in the viral replication cycle. The synthesis of 75K protein
occurred from4 to 11 h postinfection, with an
arrestat 11 h thatcoincided withthe onset of virionprotein synthesis. However, the synthe-sis ofthe 75Kprotein continued without arrest
P
ac
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.78.402.60.280.2]SYNTHESIS OF DNA BINDING PROTEIN 459
if the synthesisof viral DNA and consequently
lateproteins wasinhibited. Some investigators
haveobservedby gel electrophoresis a residual
low level of synthesis of 75K protein at late times postinfection. This synthesis could be due
to acertain degree ofasynchrony in infection.
Alternatively, since the RIPI test measures net
accumulation ofimmunologically reactive
mol-ecules rather than the rate of synthesis, we
cannot completely rule out the existence of a
residual level ofsynthesis. The arrest in
accu-mulation of 75K protein observed in the present
studycan, therefore, reflect a steady-state level
whereby residual synthesis is masked by the
inactivation of some existing 75K protein.
Progeny viral DNA synthesis is blocked in
Ad2-infected cells treated with cycloheximide
early after infection (8), leading to the
accumu-lation of five times as muchearly viral mRNA
in uninhibited cells (12). We do not know
whether the viral mRNA that codes for the 75K
protein is increased in amount by this
proce-dure. Release from acycloheximide block
pro-duces an overall decrease in the rate of
synthe-sis of host polypeptides and increase in the
synthesis of several early adenovirus-specific
polypeptides,asmeasuredbygel
electrophore-sisof extractsofcellspulselabeledimmediately
after release from inhibition (M. Harter, G.
Shanmugam, and M. Green, in preparation).
By theRIPI assay, which measures the actual
quantity of 75K protein, we observed an
ini-tially accelerated accumulation of 75K
mole-cules after release fromcycloheximide,but the
accelerated rate was not maintained even in
cells treated with Ara C toprevent arrest.
Thegene coding for the 75K protein is located
in EcoRI restriction fragment B (Grodzicker,
Anderson, and Sambrook, personal
communi-cation). A 19 to20S early mRNA mapped in this
region (3) that is H-strand specific (W.
Buett-ner, S. Veres-Molnar, and M. Green,
manu-scriptinpreparation) has the appropriate
cod-ingcapacity for a75,000-dalton polypeptide and
is a class Iearly mRNA (3) by thenomenclature
of Lucasand Ginsberg (10); i.e., it is absent or
greatly reduced inamount at 18 h postinfection.
The arrest in 75K synthesis cannot be
ex-plainedbytheexpected normal rate of decay of
mRNA, but could be due to (i) more effective
competition for available ribosomes by abun-dant late mRNA species or (ii) inhibition by
late virionorvirus-coded proteins. Inhibition of
viral DNA synthesis by Ara C may allow
con-tinuedtranscription of the mRNA for75K
pro-tein and/or the inhibition ofsynthesis of late
virionproteins may preventcompetitionand/or
regulation.
Regulated synthesis of virus-specific early
proteins that is linked to DNA synthesis has
beendescribedinseveralother DNAvirus
sys-tems. Forexample, the synthesis ofphage
T4-1000
0
10
0 8 16 24 0 8 16 24
HOURS POSTINFECTION
FIG. 5. Partitionof75Kprotein incytoplasmicand nuclearfractions of Ad2-infectedcellsinthe absence (A)andpresence(B)ofAraC.AcultureofKBcellswasinfectedand divided intotwoportions.One culture received25pgofAraC per mlat1 hpostinfection,and thesecond culturewasuntreated.Samples(30ml) wereremovedattheindicatedtimes,washed,andresuspendedin PBScontaining1%Triton X-100. Nuclear and cytoplasmic fractions were separated and assayed for 75Kprotein by the RIPI assay. Symbols: 0,
Cytoplasmic fractions; 0,nuclearfraction. VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.99.392.402.612.2]codedearly enzymesceasesat10to12minafter
infection. When phage DNA synthesis is
in-hibited or delayed, such as by infection with
phage T4 amber mutants or UV-irradiated
phage, early enzyme synthesis continues well
beyond10to12min. Theamountof enzymesat
60min after infection may reach levels four to
five times greater than after infection with
wild-typeT4phage(21). Thesynthesisof
poxvi-rus-coded thymidine kinase, an "immediate"
early protein, is also abruptlyinhibited early
after infection. Studies using inhibitors have
suggested thataviralgeneproductsynthesized
afteruncoating of virus (perhaps a "delayed"
early protein)turns off thesynthesisof
thymi-dine kinase (11). Thesynthesis of herpesvirus
type1-specificpolypeptidesiscoordinately
reg-ulated andsequentially ordered. Three classes,
a, f3, andy, weredescribedaccordingto their
timesofsynthesis. Thesynthesisofone ormore
functional a polypeptides is necessary to
"switch on" thesynthesisofif orypolypeptides,
andone or more83 or ypolypeptides"switchoff'
thesynthesis ofapolypeptides (7).
The intracellular localization of virus-specific
earlyproteins is of interest since it may provide
cluesregarding function. Wehave shown that
the 75K protein is present in both the
cyto-plasm andnucleus, and the kinetics of
accumu-lation provide guidelines for the isolation of
maximumyields forfurther studies. However,
conclusionsregardingintracellular localization
are subject to the usual cautions concerning
leakage fromthenucleusduringcell
fractiona-tion. The lag in appearance of 75Kprotein in
thenucleus and theassociation of 75K protein
with rapidlysedimenting structuresinthe
cy-toplasmic andnuclearfractions (Gileadetal.,
unpublisheddata) suggestthatthe
intracellu-larlocalization deduced from biochemical
frac-tionation may be valid. Immunoflourescence
studies withfixed-cell preparationsusing
anti-body to purified 75K protein may resolve this
importantpoint.
ACKNOWLEDGMENTS
Wethank R. Gilden ofFlowLaboratories for generous gifts ofhamster sera, without which these studies could not havebeenundertaken.
Thisinvestigationwassupported by Public Health Ser-vicegrant AI-01725 from theNational Institute of Allergy andInfectious Diseases and by research contractNO1 CP 43359within the Virus Cancer Program of the National CancerInstitute. M.G. is a ResearchCareer Awardee of the NationalInstitutesofHealth(Public Health Service grant 5K6-A14739from theNationalInstitute of Allergy and In-fectiousDiseases).
LITERATURE CITED
1. Anderson, C. W., P. R. Baum, and R. F. Gesteland. 1973.Processingof adenovirus 2-inducedproteins.J.
Virol. 12:241-252.
2. Bablanian, R., and W. C. Russel. 1974. Adenovirus polypeptide synthesisin the presence of
non-replicat-ingpoliovirus.J.Gen.Virol. 24:261-279.
3. Craig,E.A.,S.Zimmer,and H. J.Raskas.1975. Anal-ysisofearlyadenovirus2RNAusingEcoRAR1viral DNAfragments.J.Virol. 15:1202-1213.
4. Gilden,R.V.,J.Kern,A.E.,Freeman,C. E.Martin,
R.C. McAllister,H.C.Turner,and R. J.Huebner. 1968. T andtumourantigensofadenovirusgroup C-infected and transformed cells. Nature (London)
219:517-518.
5. Gilead, Z.,M.Q. Arens,S.Bhaduri, G.Shanmugam,
and M.Green. 1975. Tumourantigen specificityofa
DNAbindingproteinfromcellsinfected with adeno-virus2. Nature(London)254:533-536.
6. Gilead, Z.,and H. S.Ginsberg.1965.Characterization ofatumorlikeantigenintype12andtype 18 adeno-virus-infectedcells.J.Bacteriol. 90:120-125. 7. Honess, R. W., and B. Roizman. 1975. Regulationof
herpesvirus macromolecular synthesis: sequential
transition of polypeptide synthesis requires func-tional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A.72:1276-1280.
8. Horwitz, M. S., C. Brayton, and S. G. Baum. 1973.
Synthesisoftype2adenovirusDNA inthe presence ofcycloheximide.J.Virol.11:544-551.
9. Ishibashi,M.,andJ. V.Maizel,Jr.1974.The
polypep-tides ofadenovirus.VI. Earlyandlate glycopolypep-tides.Virology58:345-361.
10. Lucas,J. J., and H. S. Ginsberg. 1971. Synthesisof
virus-specific ribonucleic acid in KB cells infected withtype2adenovirus.J.Virol.8:203-213. 11. McAuslan,B.R. 1963.The induction andrepressionof
thymidinekinase in thepoxvirus-infectedHeLa cell. Virology21:383-389.
12. Parsons,J.T.,and M.Green.1971.Biochemical studies
on adenovirus multiplication. XVIII. Resolution of early virus-specificRNAspeciesinAd2infectedand transformed cells.Virology45:154-162.
13. Rosenwirth,B.,K. Shiroki,A.J.Levine,and H. Shi-mojo.1975.Isolationandcharacterizationof
adenovi-rustype12DNAbindingproteins. Virology67:14-23. 14. Russel,W.C.,andJ.J.Skehel.1972.Thepolypeptides
ofadenovirus-infectedcells. J.Gen. Virol. 15:45-57. 15. Russel,W.C.,J.J.Skehel,and J. F.Williams.1974. Characterizationof temperature-sensitive mutants of adenovirus type 5: synthesis ofpolypeptides in in-fected cells. J. Gen. Virol.24:247-259.
16. Sever,J. L. 1962. Applicationofa microtechniqueto
viralserologicalinvestigations.J.Immunol. 88:320-329.
17. Shanmugam, G.,S.Bhaduri,M.Arens, and M. Green. 1975.DNAbindingproteinsinthe cytoplasm and ina
nuclearmembranecomplexisolated from uninfected andadenovirus 2 infected cells.Biochemistry 14:332-337.
18. Van derVliet, P. C., andA. J. Levine. 1973. DNA-bindingproteins specificforcells infectedby
adenovi-rus.Nature(London)New Biol. 246:170-174. 19. Van derVliet,P.C.,A. J.Levine, J. M. Ensinger, and
H. S. Ginsberg. 1975. Thermolabile DNA binding proteinsfrom cellsinfected witha temperature-sensi-tive mutant ofadenovirus defective in viral DNA synthesis.J.Virol. 15:348-354.
20. Walter, G.,and J. V.Maizel,Jr.1974.Thepolypeptides ofadenovirus. IV. Detection ofearlyand late virus-inducedpolypeptidesandtheirdistribution in subcel-lularfractions. Virology57:402-408.
21. Wiberg, J. S., M. L. Dirksen, R. H. Epstein, S. E. Luria, and J. M. Buchanan. 1962. Early enzyme synthesis and its control in E. coli infected with
someambermutantsofbacteriophage T4.Proc. Natl. Acad. Sci. U.S.A. 48:293-302.
on November 10, 2019 by guest
http://jvi.asm.org/