CopyrightC 1976 American Society for Microbiology Printed inVol. 20, No. 3U.S.A.
RNA
Synthesis in Cells Infected with Herpes Simplex Virus
XIII. Differences in the Methylation Patterns of Viral RNA During the
Reproductive
Cycle
MICHAEL BARTKOSKI AND BERNARD ROIZMAN* Committee on Virology, University ofChicago, Chicago, Illinois 60637
Received for publication 20 July 1976
Herpessimplex virus 1 (HSV-1) RNA labeled with
[methyl-3H]methionine
atvarioustimes during the infectious cycle and purified by hybridization to viral
DNAwasanalyzed for the presence of methylated nucleotides. The data indicate
the following.
(i)
RNA labeled from 0 to 14 h postinfection and accumulating inthe
cytoplasm contained internal base-methylated nucleotides and terminaloligonucleotides
consistentwith thestructure7mG(5')ppp-(5')XmpYmpNp. Simi-lar methylatednucleotides and oligonucleotides were also found in viral RNA accumulatinginthe cytoplasm of cells treated with cycloheximide from thetime ofinfection.Previousstudies(M.Kozak andB. Roizman, 1974)have shownthat,
whereas the RNA accumulating in the 14-h infected cells contains all ofthe
sequences functioningasmRNA throughout infection, theRNA accumulating
inthe cytoplasm of cycloheximide-treated cells isassociated with polyribosomes
synthesizing
the earliest (a) groupofpolypeptides specified by the virus. (ii) Cytoplasmic viral RNA from cells labeled11to 14hpostinfectionaswellasthe total adenylated RNAinthe cytoplasm and polyribosomes labeledinthesame fashion contained the terminal oligonucleotide butnotthe internalbase-methyl-atednucleotide.
Previous reports from this laboratory have
shownthatinhumancellsinfected with human herpesvirus1 (herpessimplexvirus 1 [HSV-1]) viral polypeptides form at least three groups whose
synthesis
iscoordinately
regulated
andsequentially
ordered (13, 14, 21).Thus,
apoly-peptides
are made first and reach maximumratesofsynthesis between2and4h
postinfec-tion. They induce the
synthesis
of thesecond
or,8
groupmadeatmaximumratesbetween5and7hpostinfection.
These
shut off thesynthesis of
a
polypeptides and induce
thesynthesis
of the thirdory group.Thisgroup,consisting
largely
of viral structural
polypeptides (12),
in turn shuts off thesynthesis of
the/8
group.Several lines of evidence indicate that the a and
J8
mRNA's,
arerelatively
stable and that the shutoff ofsynthesis
of thesepolypeptides
is aspecific
functionofone or morepandypoly-peptides,
respectively.
Thus,
apolypeptide
syn-thesiscontinuesformanyhoursifthesynthesis
of 3
polypeptides
isblocked eitherby
rendering
a polypeptides nonfunctional (14) or
by
inter-fering
with thetranscription of(8
mRNA after amRNA
had accumulated inthe infected cell(13, 21). However, a
polypeptide
synthesis
de-cays at a
relatively
rapid
rate oncefunctional(8
polypeptides
aremade (13, 21). One expectationofasystem inwhich the
polypeptide
synthesisiscoordinatelyregulatedinacascade fashionis that the mRNA's of eachgroupshare structural features thatwould permit differential regula-tion at the level of translation (21). In the search forsuch differential features, webegan studies on the presence and distribution of methylated nucleotidesinviral RNA extracted from
the
cytoplasm
of infected cells.Methylated
nucleotides have beendemon-strated in mRNA of
eukaryotic
cells (1, 4, 20) and virus (17, 24, 30), butnotall viruses infect-ing eukaryotic cellsspecify methylated
infor-mational RNAmolecules (7). Themethylated
nucleotides occur
internally,
usually in the form of6-methyl
A, and atthe5' terminusas anoligonucleotide
of the structurem7G(5')-ppp(5')XmpYmpNp or
m7G(5')ppp(5')XmpNp,
where Xm and ym representthe 2'-O-methyl-ated derivatives of all four ribonucleosides (1). Although it. has been recently demonstrated that thepresence of m7Gatthe5' terminus is needed for the effective translation ofglobin,
vesicularstomatitis virus, and reovirus mRNA's
(2,
19), theexact functionof thesemethylatedsequences isstillunknown.
583
on November 10, 2019 by guest
http://jvi.asm.org/
In designing the experiments described in
this paper we took advantage of two
observa-tions. First, only a polypeptides are made in
cellsimmediatelyafter withdrawal of
cyclohex-imidepresent inthe medium from the time of
infection (13) or inthe presence of amino acid
analogues addedatthe time of infection(14). In
bothinstances transcripts arisingfromonly 10
to 14%of the DNAaccumulate in the cytoplasm
(16,21;N.Frenkel, H. Locker,and B.
Roizman,
manuscript in preparation). We have defined
thesetranscriptsasamRNA. Second, afterthe
onset of assembly and maturation of
viruses,
the transcripts accumulating in the infected
cell cytoplasm are homologous to 43% of the
viral DNA, and molecularhybridization
stud-iesshowthatthis RNAincludesalltranscripts
translocated into the cytoplasm, includingthe a mRNA sequences, even though the a
poly-peptidesare nolonger made (16, 21, 25).
MATERIALS AND METHODS
Cells and virus. The procedures for the
propaga-tionof HEp-2 (humanepidermoid carcinomano. 2) cells and the assay, production, andpertinent prop-ertiesoftheF strainof HSV-1 weredescribed else-where (6, 8, 15, 22).Inthesestudies, confluent
mon-olayers of HEp-2 cellswereinfected with HSV-1 (F)
at amultiplicity of10PFU/cell.
Labeling ofRNA. Infected cells were labeled at
the times specified inthetext with20to 25 mCiof
[methyl-3H]methionine
(Amersham/Searle Co.) perml ofmaintenance medium consisting ofmixture 199containing one-fifththe normal methionine
con-centrations, butsupplemented with 1%dialyzed calf
serum, 20 ,Meachadenosine and guanosine, and20 mM sodium formate toinhibit the incorporation of
[methyl-3H]methioninegroups intothepurinerings
(18).
Cell fractionation. Cells were scraped and har-vested in phosphate-buffered saline (5), washed
once,and resuspendedinlysing buffer(50 mM Tris-hydrochloride [pH 7.4]-50 mM KCl-1 mM MgCl2-0.5%Nonidet P-40 IBDHChemicalsLtd.]). After10 to 20min onice,the nucleiwereseparated from the cytoplasmicextractbycentrifugation. Sodium dode-cyl sulfate (SDS)wasaddedto afinalconcentration
of0.5%tothe cytoplasmic fraction.
ExtractionofRNA. RNA wasextracted from the
cytoplasmic fraction with phenol followedby reex-traction ofthenonaqueous residuewith Tris buffer, pH 9.0 (3). Theaqueous phaseswere combined and reextractedwithphenol-chloroform(1:1)followedby
chloroform-4% isoamyl alcohol. Inthe finalstep of
purification the RNA in a solventcontaining 0.025
M ammonium acetate was precipitated with ethanol.
FractionationofpolyadenylatedRNA. The bind-ing and elution of adenylated RNA from poly(U) immobilized on glass-fiber columns were done as
previouslydescribed (26).
Enzymatic digestionofRNA. RNA was digested
withT2RNase(30Ag/ml;SigmaChemicalCo.) in 10
mM sodiumacetate, pH4.6. P1 nuclease
digestion
(200 .g/ml; Yamasa, Ltd., Japan) wasdone in 10 mM acetate buffer, pH 6.0. For alkaline
phospha-tasedigestion(10 Ag/ml;SigmaChemicalCo.)after
P1 digestion, thepHwasraisedto8.2with Tris.
DEAE-Sephadex chromatography. The nuclease
digestion products were analyzed by chromatogra-phyonDEAE-Sephadex(A25)columns
equilibrated
with 20 mM Tris-hydrochloride (pH 7.4), 0.05 M
NaCl,and 7 Murea(29).Thedigestwaselutedfrom the columns withlineargradients of0.05 to 0.4 M NaCl in theTris-urea bufferandclearedcompletely
with1 MNaCl.
The eluent fractionswerepooledanddesaltedby
thebariumprecipitation proceduredescribedbyRoy
andBishop(23). The elution ofnucleotidesand
oli-gonucleotides withcharges rangingfrom -2 to -7
wasstandardizedwiththe aid ofpancreaticRNase
digestofEscherichiacolitRNA.
DNA-RNAhybridization. TheDNA-RNA
hybrid-izationswerecarriedout byoneoftwoprocedures.
Inthefirst, DNAwasimmobilizedonSepharoseas
described by Gilboa et al. (11). The hybridization
wasdone in 5x SSC(1XSSC contains 0.15 MNaCl plus0.015 Msodiumcitrate)and 50% formamideat
50°C. Afterhybridization, the DNA-Sepharose
col-umn was washed extensively with hybridization buffer. The
hybridized
RNA wasthen eluted with98% formamide-10mMTris(pH7.4) followedbya1
M NaCl wash. The second procedure involved hy-bridization ofRNA to viral DNA fixed to
nitro-cellulose filtersasdescribedby Kieffetal. (15).The
hybridizations were done at 50°C for 48 h in 0.8 ml ofhybridization buffer containing0.75 M NaCl,
0.1 MTris(pH7.4), 0.5 mMEDTA,0.1% SDS, and 50%formamide. Afterhybridizationthefilterswere
soaked in 1 x SSC containing 0.5% SDS, reincu-bated for 1h,and then rinsed in 2 xSSC. The RNA
wasthen released by incubatingthe filters in95% formamide-30mM Tris (pH8.2)-0.1%SDS for1hat
50°C. Released RNA was precipitated with 2
vol-umesofethanol and stored at -20°C.
RESULTS
The studiesdescribedinthispaperconsisted
ofthree series of experiments. In the first we
examined the methylated nucleotides in
cyto-plasmic virus-specific RNA labeled with
[methyl-:H]methionine from the time of
expo-sure of cells to virus until 14 h postinfection.
Previous studies from this laboratory have
shown thatthecytoplasm of 14-h infectedcells
contains RNA sequences homologousto 43%of
DNA. This RNA represents all informational
sequences present incytoplasmthroughout
in-fection even though only a polypeptides are
madeatthattime (16, 21). In this seriesHEp-2
cells were infected with HSV-1 (F) at a
multi-plicityof 10PFU/cell and labeled with
[methyl-IHimethionine from 0 to 14 h postinfection as
described above. Atthattimetheinfectedcells
were harvested, and nuclear and cytoplasmic
fractions were separated as described. Viral
J.
on November 10, 2019 by guest
http://jvi.asm.org/
RNA SYNTHESIS IN CELLS INFECTED WITH HSV 585
RNA was purified by hybridization to viral
DNA immobilized on Sepharose (11), eluted,
and then analyzed for thepresence of
methyl-ated nucleotides. In these analyses the T2
RNase digest of purified RNA was
chromato-graphedonDEAE-Sephadex in thepresenceof
7Murea(29). This procedureseparates
nucleo-tides and oligonucleonucleo-tides on the basis of the
number of negatively charged phosphates.
Thus, each internal phosphate contributesa-1
charge, whereas each free terminal phosphate contributesa-2charge.Themethyl-3Hlabel in
theHSV-1(F) RNAdigest chromatographedas
two peaks, which eluted at positions
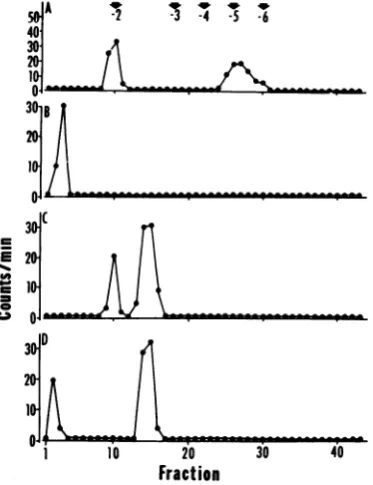
corre-spondingto-2and -5to -6charge (Fig. 1A).
Peak 1, containing approximately 37% of the
totalelutedactivity(Table1),consisted of
base-methylated nucleotides. This conclusion is basedonitscharge(-2)andontheobservation
that subsequenttreatmentwith alkaline phos-phatase convertedittoanucleoside that didnot
absorb to the DEAE-Sephadex column (Fig.
1B). The material elutinginpeak1corresponds
in its properties to internal base-methylated
nucleotides (1, 24, 30). The -5 to -6 charged material inpeak 2 (Fig. 1A) wasdesalted,
di-gested with P1 nuclease, which degrades RNA to5'-nucleotides(9),and splitintotwoportions.
Oneportion, designated as the P1 digest, was
chromatographed on DEAE-Sephadex in 7 M urea. The otherportion,designatedasP1
alka-line phosphatase digest, was redigested with
alkaline phosphatase and then
rechromato-graphed asabove. The P1 digest eluted in two
peaks (Fig. 10). Thefirstpeak, containing
ap-proximately one-third of the label, eluted at a
position corresponding to a mononucleotide
witha -2charge.The secondpeak,containing two-thirds of the label,elutedataposition (-2
to -3) asexpected for a structure of the type m7G(5')ppp(5')Xm (10). The P1 alkaline phos-phatase digestion (Fig. 1D) did not, however,
affect the elution of the peak2 ofFig. 1C. We
interpretthese results to indicate that P1
nu-clease digestion released the twoterminal
nu-cleotides (Ymp, Np) of which the methylated nucleotideeluted in peak1 (Fig.1Cand D) and
that the remaining dinucleotides of the type
m7G(5')ppp(5')Xm elutedinpeak2(Fig.1C and
D). The results of these analysesareconsistent
54
40
301
210
1le
E
m 0
30
10
A X l;5 6
.
D
ilo
i20
30 40Fraction
FIG. 1. Elution profile of HSV-l-specificRNA
di-gest on DEAE-Sephadex columns. The RNA was
labeled with [methyl-3H]methionine from 0to 14 h postinfection. (A) T2RNasedigest ofHSV-1 RNA.
(B) Rechromatography of peak1 of (A) after
desalt-ingandalkaline phosphatase digestion. (C)
Rechro-matography of peak 2 of (A) after desalting and
digestion with P1 nuclease. (D) P1 digest of (C) further digestedwith alkalinephosphatase. The
po-sitionofcharges-2to-7wasstandardized, usinga
pancreaticRNasedigest ofE. coli tRNA.
TABLE 1. Distributionofmethyl-3H-labeled nucleotidesinselected RNAsfromHSV-1-infectedcells [methyl- [methyl-3H]methionine-la-
3H]methionine-la-Labeling beledmononucleo- beled oligonucleotides Infected cell RNAdigested withT2RNase afterinfec- tidesindigest(-2 indigest(-5to-6
tion(h) charge) charge)
cpm %of total cpm % of total
Cytoplasmic HSV-1 specificRNAa 0-14 60 37 99 63
Cytoplasmic HSV-1specificRNAb 0-8 25 55 20 45
Adenylated polyribosomal HSV-1specific 11-14 2 5 48 95
Adenylated totalcytoplasmic 11-14 80 1 6,880 99
Adenylated polyribosomal 11-14 50 1 3,550 99
aHSV-1-specific RNA referstoRNAselectedbyhybridizationtoHSV-1DNAasdescribedintext. bFromcellsinfected and maintainedinthepresenceofcycloheximide.
04
a
VOL. 20, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.496.248.435.152.395.2] [image:3.496.43.439.521.631.2]with the hypothesis that HSV-1 RNA
accumu-lating in the cytoplasm has a modified 5'
terminus predominantly of the type
m7G(5')-ppp(5')XmpYmpNp, designated as cap 2,
al-though we cannot exclude the possibility that
modified5' terminiof thetype
m7G(5')ppp(5')-XmpNp, designated as cap 1, arealso present.
This conclusion is based on a comparison of
labelinpeaks1and2of Fig. 1C. Assumingthat each nucleotide has a single methylation, the predicted ratio oflabel in peak 1 to peak 2 of
Fig. 1C is 1:2for cap2. P1nucleasedigestionof
acap1structureshould yieldno countsinpeak
1. Since we are observing the maximum ratio we could expect for pure cap 2, the extent of contamination with cap 1 structures is prob-ablynotvery high.
Inthesecond seriesof experiments we
exam-ined RNA accumulating in the cytoplasm of
cells treated withcycloheximidefromthe time
ofinfection. As indicated in the introduction,
the RNA extracted frompolyribosomesorfrom
total cytoplasm of cells treated with
cyclohexi-mide is homologous to 10to 12% of viral DNA
(16, 21). On withdrawal of the drug, the cells
make only a polypeptides (13, 21). In these
experiments HEp-2 cells were infected with
HSV-1 (F) at amultiplicityof10PFU/cells, in
the presence ofcycloheximide (50 ,ug/ml), and
thenmaintainedinthe presence of thedrugfor
8h.The cells werethenfractionated, andRNA
wasextracted from the cytoplasm asdescribed
above. Cytoplasmic viral RNA purified by
hy-bridization toviral DNA wasdigested with T2
RNase and chromatographed as above. The methyl-3H-labeled material from
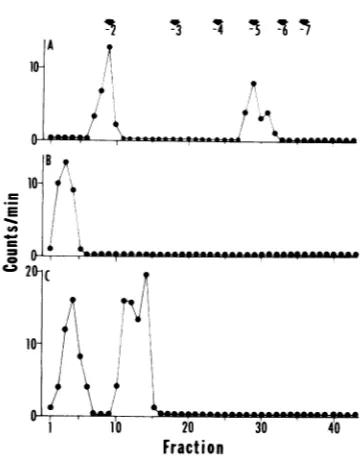
HSV-1-spe-cific RNA eluted from the columnin two peaks
(Fig. 2A and Table 1) in a pattern similar to
that ofthedigestof HSV-1 RNAlabeledfor0 to
14hpostinfection (Fig. 1A). The twopeaks with
-2 and -5 to -6 charges, respectively, were
collected and desalted for further analysis.
Peak 1 (Fig. 2A) was shown to consist of a
mononucleotide because subsequent digestion
with alkaline phosphatase converted it to a
nucleoside that did not absorb to the column
(Fig. 2B). Peak2(Fig. 2A) was digestedwithP1
nuclease and alkaline phosphatase. The
prod-ucts werethenchromatographedasabove(Fig.
2C). Approximately one-third of the methyl-3H
label activity was converted tonucleosides that
didnotabsorbtothe column (peak 2, Fig. 2C).
However, the remainingmethyl-3H-labeled
ma-terial retaineda -2 toa -3charge (peak 2, Fig.
2C), corresponding in its elution properties to a structure ofthe type m7G(5')ppp(5')Xm (10). It
appears from this analysis that HSV-1 RNA
madeinthe presence of cycloheximide contains
10-E
U,
a A
I
1 I
L ----F---
E----20
i1
i 10 20
Fraction
30 40
FIG. 2. Elution profile of HSV-1-specificRNA di-gest on DEAE-Sephadex columns. [3IH-methylimethionine from0to 8hpostinfection in the presenceofcycloheximide (50
Mig/ml)
asdescribed in the text. (A) T2 RNase digest ofHSV-1-specific RNA. (B)Rechromatography of peak 1 of (A) after desalting and alkaline phosphatase digestion. (C) Peak2of(A) rechromatographedafterdesalting and digestion with P1 and alkaline phosphatase.both terminal and internal methylated nucleo-tides.
The objective of the third series of
experi-ments was to examine the mRNA specifying
the last or y group of viral polypeptides.
How-ever, because (i) the RNA sequences associated
with polyribosomes of infected cells making a
polypeptides are conserved and cosediment
with polyribosomes even after a polypeptide
synthesis ceases (16, 21) and (ii) current evi-dence strongly suggests that the templates for the three groups of polypeptides are intermixed
rather than clustered (P. Jones, G. S.
Hay-ward, and B. Roizman, J. Virol., in press),
itwasnot feasible to obtain purified y mRNA.
We have therefore examined only the RNA
labeled and entering into the cytoplasm late
in infection without prejudice as to whether
this RNA is capable of specifying all classes
or only y polypeptides. In designing this ex-periment we took advantage of the
observa-tionthatall or most species of viral RNA
accu-mulatinginthecytoplasm of infected cells are
adenylated and bind to poly(U)bound to
glass-fiber filters (26). Inone experiment the
(poly)A-containing RNA was hybridized to viral DNA, B
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.272.454.58.287.2]RNA SYNTHESIS IN CELLS INFECTED WITH HSV
587
eluted,
and thendige
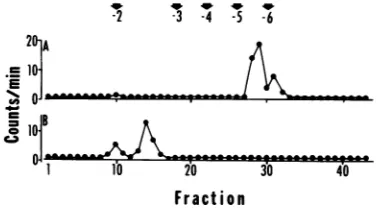
described above. The el double
peak
ofradioa4
tetra- and
pentanuclec
(Table
1 andFig.
3A)
less than 5% ofthe to
position
ofmononucle3A).
Theradioactivity
witha
charge
of-5to anddigested
with Plraphy,
thispeak elutez
charge
(Fig.
3B).
Th with a"capped"
m7G(5')ppp(5')X'pYml
although
in this inst could also bepresent.
base-methylated
nucleentering
cytoplasm
Iquestions
as to whetl fered from all other Xmade atthat time. A(
the total
cytoplasmic
bound and eluted from
glass-fiber
columns.T;
nylated
RNAspecies
aplasm
share with virciency
ininternal basDISC] The salientfeatures
thispaperare asfollov
themRNA'sof
eukary
some viruses
infecting
RNA
accumulating
in bosomesof infectedcelterminus. The modifi
in a
mRNA,
inviral:
infection,
and inviralported
intothecytople
-2 -3
20]
.5
10]
Ei
A
ID
F
FIG. 3. Elutionprofile
gest on DEAE-Sephadex
labeledfrom11 to14 hp4
digest of HSV-specific RA ofmaterial eluting with
desaltinganddigestionu
linephosphatase.
,sted with T2 RNase as cause current methodology does not permit the
lutionprofile showedone recovery of adequate amounts of viral RNA, we
ctivity atthe position of have not sequenced the modified
5'-ologonu-)tides (-5 to -6 charge) cleotide. The chromatographic behavior of the
A minor peak, usually oligonucleotide and its digestion products
indi-)tal, was detected at the cates that the form m7G(5')ppp(5')XmpYmpNp,
otides (-2 charge) (Fig. commonly seen in many other mRNA species,
eluting from the column predominates on viral RNAs accumulating in
-6 waspooled, desalted, the
cytoplasm.
We do not know whether allnuclease. On chromatog- modified termini are identical; these studies
dwith a -2and -2to -3 awaitpurificationofindividual mRNA's.
is pattern is consistent (ii) In conformity with mRNA of other
spe-5' end structure cies, the T2 RNase digest ofHSV-1 (F) RNA
pNp
as described above, yieldedbase-methylated mononucleotides.TwoLance a cap 1 structure observations are of special interest. The first
The absence of internal relates to the fact that viral RNA sequences ,otidesinHSV-1(F)RNA
normally
made andtranslocated intocytoplasm late in infection raised immediately after infection contain internal ier HSV-1 (F) RNA dif-base-methylated
nucleotides, whereasthe RNAadenylated
RNA species made and translocated into the cytoplasm at ccordingly, we examined the time of peak synthesis of y polypeptides andpolyribosomal
RNA doesnotcontainappreciable
amountsof these ipoly(U) immobilizedon nucleotides. We cannot atthis time determine able1shows that all ade- whether the absence of internal base-methyl-Lccumulatinginthecyto- atednucleotidesin RNAmade11 to 14h afterral RNA species a defi- infection is the consequence of the absence of
;e-methylated
sequences.signals
forbasemethylation
inRNA,
inhibitionof the enzyme
by
avirus-specific product,
orUSSION rapid turnover coupled with a lack of new
syn-;of the data presented in thesis of the enzyme because of viral inhibition
fs.
(i)
In conformity with of hostprotein synthesis
(27, 28). The fact thatroticcells (1, 4, 20) andof all adenylated RNA, which includes both host
them (17, 24, 30), HSV and virus-specific RNA, lacked the
base-meth-the cytoplasmon polyri- ylated nucleotides suggests the absence of
ac-lls contains amodified5' tive enzyme, but this remains to be
deter-ad terminus waspresent mined. It is perhaps useful to note that among
RNAlabeled
throughout
the various possibilities left open by thisobser-RNA labeled andtrans- vation are extreme ones. The first is that the
asm lateininfection. Be- disappearance ofa host enzyme due to rapid
decay
and overallinhibition ofhostprotein syn-thesismayinfactserve aregulatory
function in-4 -5 -6 thatitimparts onthe RNAastructural
altera-tionwhichcouldserve todifferentiate this RNA
from otherspecies. At the otherextremeis the
possibility
that internalbase-methylated
nu-cleotides have no
regulatory
function.Experi-mentsare inprogressto differentiate between
these
possibilities.
The secondobservation ofinterestrelates to
20
30 the fact that cytoplasmic viral RNA labeledraction from 0 to 14 hpostinfection contained internal ofHSV-1-specificRNAdi-
methylated
nucleotides,
whereas the RNAla-oflHSVm1nspeific
RNAdas
beledfrom 11 to 14h didnot. Wedonot knowcolumns. The RNA was
ostinfection.
(A) T2 RNase theprecise time
after infection when internaliA.
(B)Rechromatography base methylation ceases; the data indicate, a -5 to -6 charge afterhowever,
that the RNAcontaining
the internalvith Pl nuclease and alka-
methyl-3H-label
is conserved; the data alsoreinforce the conclusion reached from
experi---I'1---
--tm VOL. 20, 1976
a
I lo
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.46.236.474.579.2]mentspublished earlier thataRNAsequences
are present in the cytoplasm of infected cells
many hours after a polypeptides cease to be
made (16, 21).
ACKNOWLEDGMENTS
Wewishtothank Aaron Shatkin and his associatesat
the Roche Institute for invaluable advice in the early
stagesof thesestudies.
These studies were aided by grant VC 103L from the American Cancer Society and Public Health Service Grants CA 08494and CA 19264 from the National Cancer Institute. M. B. isaPublic Health Servicepostdoctoral trainee (Al
00238) of the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Adams, J., and S. Cory. 1975. Modified nucleosides and bizarre 5'-terminiin mousemyeloma mRNA. Nature
(London) 255:28-33.
2. Both, G. W., A. K. Banerjee, and A. J. Shatkin. 1975. Methylated dependent translation of viralmessenger
RNAs in vitro. Proc. Natl. Acad. Sci.U.S.A. 72:1189-1193.
3. Brawerman, G.,J. Mendecki, and S. Y. Lee. 1972. A procedure for the isolation of mammalianmessenger
ribonucleic acid. Biochemistry 11:637-641.
4. Desrosiers, R., K. Frederici, and F. Rottman. 1974. Identification ofmethylated nucleosides in
messen-gerRNA from NovikoffHepatoma cells. Proc. Natl. Acad. Sci. U.S.A. 71:3971-3975.
5. Dulbecco, R., and M. Vogt. 1954. Plaque formation and isolation ofpurelineswithpoliomyelitisviruses. J. Exp.Med. 99:167-182.
6. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968. Characterization of herpes simplex virus strains dif-fering in their effectson the social behavior of
in-fected cells. J.Gen. Virol. 2:357-364.
7. Fernandez-Munoz, R., and J. E.Darnell. 1976. Struc-tural difference between the 5' termini of viral and cellular mRNA in poliovirus-infected cells: possible
basis for the inhibition of host proteinsynthesis. J. Virol. 18:719-726.
8. Frenkel, N., S. Silverstein, E. Cassai, and B. Roizman. 1973. RNA synthesis in cells infected with herpes simplex virus. VII. Control of transcription and of transcript abundancies of unique and common
se-quencesofherpes simplex virus 1 and 2. J. Virol. 11:886-892.
9. Fugimoto, M., A. Kuninaka, and H. Yoshino. 1974.
Substrate specificity of nuclease P1. Agric. Biol. Chem.38:1555-1561.
10. Furiuchi, Y., M. Morgan, S. Muthukrishnan, and A.J.
Shatkin. 1975. ReovirusmessengerRNA containsa
methylated, blocked5'-terminalstructure: m7G(5')-ppp(5')GmpCp. Proc. Natl. Acad. Sci. U.S.A. 72: 362-366.
11. Gilboa, E., C. Prives, andH. Aviv. 1975. Purification of
SV40messengerRNAbyhybridizationtoSV40 DNA covalently boundtosepharose. Biochemistry 14:4215-4220.
12. Honess, R. W., and B. Roizman. 1973.Proteins
speci-fiedby herpes simplex virus. XI. Identificationand
relative molar rates of synthesis of structural and
nonstructural herpesvirus polypeptides in the
in-fectedcell. J.Virol. 12:1347-1365.
13. Honess, R. W., and B. Roizman. 1974. Regulationof herpesvirus macromolecular synthesis. I. Cascade
regulation of the synthesisof three groups of viral proteins. J.Virol. 14:8-19.
14. Honess, R. W.,and B. Roizman. 1975. Regulation of herpesvirus macromolecular synthesis: sequential
transition of polypeptide synthesis requires
func-tional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A. 72:1276-1280.
15. Kieff, E., B. Hoyer, S. Bachenheimer, andB. Roiz-man. 1972.Geneticrelatednessof type 1 andtype 2 herpessimplex virus. J. Virol. 9:738-745.
16. Kozak, M.,andB. Roizman. 1974. Regulationof her-pesvirus macromolecular synthesis: nuclear
reten-tion of non-translated viral RNA sequences. Proc. Natl.Acad. Sci.U.S.A. 71:4322-4326.
17. Lavi, S., and A. J. Shatkin. 1975. Methylated simian virus 40 specific RNA from nucleiandcytoplasm of infected BSC-1 cells. Proc. Natl. Acad. Sci. U.S.A. 72:2012-2016.
18. Maden, B., M.Salem,andD.Summers. 1972. Matura-tion pathway for ribosomalRNA in the HeLa cell nucleolus. Nature(London)NewBiol. 237:5-9. 19. Muthukrishnan,S., G.Both,Y.Furuichi, andA.
Shat-kin. 1975. 5-terminal 7-methyl-guanosine in eukar-yotic mRNA isrequiredfor translation. Nature (Lon-don)255:33-37.
20. Perry, R.P.,andD.Kelley.1974.Existence of methyl-atedmessengerRNA in mouse L cells. Cell 1:37-42. 21. Roizman, B., M. Kozak, R.W. Honess, and G.
Hay-ward. 1974.Regulationofherpesvirus
macromolecu-lar synthesis: evidence for multilevel regulation of herpes simplex 1 RNA andprotein synthesis. Cold Spring HarborSymp. Quant.Biol. 39:687-702. 22. Roizman, B., and P. G. Spear. 1968. Preparation of
herpessimplexvirusofhightiter.J. Virol. 2:83-84. 23. Roy, P., and D. H. L. Bishop. 1973. Initiation and
direction of RNA transcriptionby vesicular stomati-tis virus virion transcriptase. J.Virol. 11:487-501. 24. Shatkin,A. J. 1974. Methylatedmessenger RNA
syn-thesis in vitrobypurifiedreovirus.Proc.Natl. Acad.
Sci. U.S.A. 71:3204-3207.
25. Silverstein,S.,S. L. Bachenheimer, N.Frenkel,and B. Roizman. 1973. The relationship between post
tran-scriptional adenylation of herpesvirus RNA and
mRNA abundance. Proc. Natl. Acad. Sci. U.S.A.
70:2101-2104.
26. Silverstein, S.,R.Millette,P.Jones, and B. Roizman. 1976. RNA synthesis in cells infected with herpes simplexvirus. XII.Sequence complexity and proper-ties of RNA differing in extent of adenylation. J.
Virol. 18:977-991.
27. Spear, P.G.,andB.Roizman. 1972. Proteinsspecified by herpessimplex virus. V. Purification and
struc-turalproteins of theherpesvirion.J.Virol. 9:143-159. 28. Sydiskis,R.J.,andB. Roizman. 1967.The
disaggrega-tionofhostpolyribosomesinproductive and abortive
infection withherpessimplex virus. Virology 32:678-686.
29. Tener,G. M. 1967. Ion-exchangechromatography in the presence of urea, p. 398-404. In L. GrossmanandK. Moldave(ed.),Methods in enzymology, vol. XII. Aca-demic Press Inc., New York.
30. Wei, C.-M., andB. Moss. 1974. Methylationofnewly
synthesizedviral messenger RNA by an enzyme in vacciniavirus. Proc.Natl.Acad. Sci. U.S.A. 71:3014-3018.