Copyright © 1977 American Society for Microbiology Printed in U.S.A.
Replication Process of the Parvovirus
H-1.
VI.
Characterization of a Replication Terminus of H-1
Replicative-Form DNA
SOLON L. RHODE III
PutnamMemorial Hospital Institute for Medical Research, Bennington, Vermont 05201
Receivedfor publication 11June1976
Thelinearduplexreplicative form (RF) DNA of the parvovirus H-1 has been
characterized withrespect tocleavageby thebacterial restriction endonuclease
of Escherichia coli, EcoRI. RF DNA has a single cleavage site 0.22 genome
length fromthe left end ofthemolecule. Themolecular weight ofH-1RF DNA determined by gel electrophoresis is 3.26 x 106. H-1RF DNA has beenfoundto dimerize by hydrogen-bonded linkage at the molecular left end, and in some molecules the viral strand is covalently linked to the complementary strand.
Some 10%ofmonomeric RFDNA also hasacovalent linkage betweenthe viral
and complementary strandsatthe left end. TheEcoRI-B fragment, containing the left end of the RF molecule, appears to be a replication terminus by its labelingcharacteristics forboth RFand progeny DNA synthesis. These findings
suggest that the left end of H-1 RF DNA has some type of "turn-around"
structureandthat this endisnotan origin forDNAsynthesis.
The parvovirus H-1 contains a genome of
Nevertheless,
itis clear that thegenetic
capac-single-stranded
DNAwithamolecularweight
ity
of H-1 is verylimited,
andthus itsreplica-of
approximately
1.7 x 106(15). We haveprevi-
tion must be executedlargely by
cellularen-ously characterized
a double-stranded DNA zymes. Anumber of conditionallethal mutantsreplicative form (RF)
synthesized
during
the ofH-1have been isolated andnoneofthese has replication ofH-i-infected cells (17, 18). H-1RF shownaviralproteinrequirement
for RFDNA DNAhas been foundtobe linearbyisopycnic
replication (19, and unpublished data). It is centrifugation in ethidium bromide-CsCl gra- possible, but not established, that RFreplica-dients, velocity
sedimentation(17, 18),gel
elec- tionistotally
under the control of cellular pro-trophoresis (in this paper), andby
electronmi- teins. Inany case,analysis ofRFreplication
iscroscopy (23). a powerful probe of cellular DNA replication
In this study, H-1 RF DNA and its dimer and could be usefulinthestudy of the mecha-have been characterized further. An
improved
nismsofreplication of linearmolecules.estimateofH-1 RF DNAmolecular
weight,
3.26 Ihave examined thephysicalmapof H-1RFx 106, has beenobtained by gelelectrophoresis. DNA with respect to the bacterial restriction
Basedon a DNAsize of about 4,920 basepairs endonuclease of Escherichia coli, endo
(bp),
the H-1 genome has a totalcapacity
to R-EcoRI (EcoRI). This enzyme has allowedacode forproteins with acumulative molecular moredetailed characterization of one end of the
weightof about 172,000. Todate, only thetwo RF DNA molecule, and evidence for several polypeptides of the H-1
capsid,
VP1 andVP2,
structural alternatives for this end arepre-have been demonstrated as viral-induced pro- sented. Experiments described here indicate
teins ininfectedcells, andtheir total molecular that thisend is a terminus for the
semiconser-weightof 164,000 is close tothecoding capacity vative replication of RF DNA and for progeny
of the genome (14). It hasnotbeendetermined DNAsynthesis. This terminus is in some cases
whether the H-1 genome is transcribed to the self-cohesive, as manifestedby the formation of
extentof the otherparvoviruses (50to 100%of dimer molecules linked in aninvertedmanner
the rat virus viral strand is transcribed
[20],
ratherthan in tandem. The viral (V) andcom-and 75% of the adenovirus-associated virus plementary (C) strands in some monomer or
[AAV]
genome istranscribed[3-5]),
orwhether dimer RF molecules are covalently linked atVP1 andVP2 havea commonstructural gene. this end. These findings suggest a structure for
694
on November 10, 2019 by guest
http://jvi.asm.org/
the RF DNA replication terminus similar to (Haemophilus aegyptius, kindly provided by David one recently proposed for the AAV DNA ter- T. Denhardt)werecarriedoutfor 16 hat37°Cin50
mini (8, 24). ml of 10 mM Tris (pH 7.5)-10 mM NaCI-10 mM
MgCl2-lmMdithiothreitol and with5,lI of enzyme.
MATERIALS AND METHODS Reactions were stopped by addition of EDTA to a Virus and cell strains. Parasynchronous cultures finalconcentration of 50 mM.
ofsecondary hamster embryoor NBcellswerepre- Analytical gel electrophoresis. Electrophoresis pared and infectedaspreviously described (14, 16). wasconducted in cylindrical gels (0.6 by 15 cm) with TheH-1 strains used here were wild type (wt) and various concentrations of agaroseor 3% acrylamide-ts 1 (19). AnE.coli (Wi100)lysogen ofXcI857 susS 0.5% agarose in E buffer (40 mM Tris-20 mM Na wasobtained from Herbert Boyer, San Francisco. acetate-1 mM EDTA) (9) at 23°C under constant Infection, labeling, and viral DNA extraction. voltage as specified in the text. DNA samples were Parasynchronous hamster embryo cultures were in- mixedwith 0.5 volume of a solution of E buffer, 50% fected at12h poststimulation, and NB cell cultures glycerol, and 0.01% bromophenol blue for layering wereinfected at the time of reversal of the metho- on thegel. Gels were sliced into 1.0-mm or 1.17-mm trexate block with medium containing thymidine slices,and two slices pervial were incubated2to16
(TdR; 10-5M), adenosine (13 ,ug/ml), and minimal h in 0.5 ml of NCS solubilizer (Amersham/Searle) essential medium nonessential amino acids as a and assayed for radioactivity by liquid scintillation
source of glycine (19). Virus adsorptions were per- spectrometry in 10 ml of a toluene-based scintilla-formed for 30 min at33°C. After virus adsorption, tionfluid. Ninety percent or more of the activity is the cultures were incubated at the temperatures eluted from the gel slices by this method.
indicated. Viral DNA was labeled by changing the Preparative gel electrophoresis. Viral DNAwas mediumafter two washes with Hanks balanced salts fractionated in cylindrical gels (1.2 by 6 cm) of 1% (5 ml per petri dish) to aprewarmed medium con- agaroseinEbuffer using a constant voltage of 150 taining the appropriatelabeled precursor. In most V. DNA infractions of 2 ml was collected every 10 cases, labeled TdR was usedinthe presence of 5'- minby continuous electroelution into a chamber of fluorodeoxyuridine (FUdR) at 0.5
Ag/ml
after en- ourdesign.Aliquotsweretaken fordetermination of dogenous pools of TdR had been depleted by a prein- radioactivity, and appropriate fractions were col-cubation in medium with FUdR alone. Labeling lectedand precipitated with0.1 volumeof3 M Nawith32po4wasdoneinmedium buffered by 25mM acetate(pH 5.0)and2.5volumes ofethanol for16or
HEPES (N-2-hydroxyethylpiperazine-N'-2-ethane- more h at -20°C.
sulfonicacid)and with the appropriate Cl salts sub- Slab-gel electrophoresis. Slab-gelelectrophoresis stituted for PO4inthe medium. wascarriedoutingels (0.3 by12by16cm) of 1.8% Viral DNA was extracted by amodification of the agarose in an EC470 apparatus (E-C Apparatus
Hirt procedure (12). The monolayer cultures were Corp., Philadelphia, Pa.). The gel and electrode washed once with 5 ml ofTris-buffered saline per buffers were buffer E, and electrophoresiswasfor18 100-mmdish and lysed with 0.5 ml of0.6%sodium hat 16°C witha constant voltage of50 V. Thegel dodecyl sulfatein 50 mMTris (pH 7.5)-10 mM EDTA wasdriedinvacuoand exposed toKodakno-screen
perdish. After15to 20minat roomtemperature,0.5 X-rayfilmNS2T for4daysbeforedevelopment.
mlof asolution containing 5 mg of Pronase per ml Velocity centrifugation. Neutral sucrose
gra-was added to each dish, and the incubation was dientswere 5 to20% sucrose in 50 mMTris(pH 8.0)-continuedfor30minat37°C. A solution of5MNaCl 1MNaCl-1 mM EDTA-0.2% Sarkosyl centrifugedin
was then added dropwise to each dish to a final anSW25rotorfor18 to 22hat24,000 rpm.Alkaline concentration of 1 M and gently mixed with the sucrose gradients were 5 to 20% sucrose in 0.2 N lysate. The lysates were collected into centrifuge NaOH-0.8 M NaCl-1 mM EDTA centrifuged inan tubes and incubatedovernight at0°C.After centrif- SW50 rotorfor 16hat 30,000 rpm. Fractions were
ugation at 25,000 xgfor 50 min, the supernatant, collectedthrough the bottoms of the tubes. Aliquots containing viral DNA, was precipitated with 2.5 weretaken fordetermination ofradioactivity either volumes of ethanol for 6 to 16 h at -20°C. The by solubilization directlyin 10 volumes of NCSor precipitateswereredissolvedin 50 mMTris (pH 7.5)- after precipitation on cellulose acetate filters with
1 mM EDTA anddigested with pancreatic RNase 10%trichloroaceticacid and100,ug of bovine serum (heatedto80°C for20min) at 50 ug/ml. Theywere albumin, followed by liquid scintillation spectrome-theneither extracted with phenolordirectly frac- try. Alkalinegradientswerefractionatedon 25-mm
tionatedby centrifugation in 5to 20% neutral su- Whatman no. 3 filter paper disks, which were
crose gradients as detailed previously (17). Gener- washed twice in cold 5% trichloroacetic acid and ally,abouttwo-thirdsofthe totalisotopic incorpora- once in acetone, dried, andcounted with 0.2 ml of tion was recovered asviralDNA in the Hirtsuper- NCS solubilizerandtoluene scintillationfluid. natant. [3H]TdR-labeled X DNA was prepared as Isopycnic centrifugation. DNA was banded to described (11). equilibrium inCs2SO4 as previouslydescribed (17, Digestion with restriction endonuclease. Viral 19),andaliquotsweremeasured forradioactivityas DNAdissolved in 50to 100 ,ul of100 mM Tris (pH for alkalinesucrosegradients.
7.5)-50mMNaCl-10 mMMgCl2wasincubated with Benzoylated DEAE-cellulose (BDC) chromatog-10 to chromatog-100Uof EcoRI for2or 3h at37°C. Endonucle- raphy. BDC was purchased from Schwarz/Mann
ase R*EcoRI was purchased from Miles Laborato- (Orangeburg, N.Y.).Columns(1by5cm)were con-ries, Inc. Digestions with endonuclease R-HaeIII structedand washedwith 100to200volumesof0.3
on November 10, 2019 by guest
http://jvi.asm.org/
M NaCI-50 mM Tris (pH 7.5)-l mM EDTA (0.3 M above. Final recoveries were 80 to 100% of theDNA
TBS). DNA in 2 to 5 mlof0.3 MTBSwasappliedto applied to the columns. the columns with aflowrateof1 ml/min, and the
column waswashed with15-, 30-,and 30-ml volumes RESULTS of0.3 MTBS. Double-stranded DNA waselutedby
two 10-ml applications of 1 MTBS, and DNA with Endo R-EcoRI digestion of H-1 RF DNA.
single-stranded regions was similarly eluted with H-1 RF DNA labeled with [3H]TdR was
pre-two 10-ml applications of 2% caffeine in 1 M TBS pared from
tsl H-1-infected
parasynchronouswarmed to40°C.Insome cases,the caffeine elution NB cells incubated at the restrictive
tempera-was carriedout with 20 ml of a linear gradient of 0 to ture of39.5°C
(17).
A portion of the major peak2%caffeinein 1MTBS. Thewashesand aliquots of
thel M TBS and 2% caffeine fractions were assayed at
i6S
in the preparative neutral sucrosegra-for radioactivity bytrichloroacetic acidprecipitation dient was digested with EcoRI for 2 h at 37°C
as above. DNA in the eluates was precipitated for and a control incubated in the absence of the
further studies by 2.5 volumes of ethanol overnight enzyme, and thedigestsweresubjected to
elec-at -200C. trophoresis in replicate agarose gels; the results
Alkaline denaturation-BDCchromatography was are illustrated in Fig. 1.EcoRI-cleavedH-1RF
done by incubating the DNA in a final concentration DNA into a large
fragment,
RI-A, and one orof 0.2 N NaOHina volumeof 200
Al
for 20 min at more smaller fragments. This digestion did notambient temperature; the solution was cooled to
0C, neutralized, and diluted 25-fold with cold0.2 M go to completion, but identical results were
ob-TBS containing 0.2 ml of 0.2 N HCl. To minimize tamned with complete digests using RF DNA
intermolecular annealing, the DNA solution was prepared with either tsi or wt H-i and with
immediately applied to the column of BDC within1 electrophoresis in3% acrylamide-0.5% agarose
to 2 min.The elutions were carried out as described gels. The EcoRI-Afragment containedan
aver-* I
I
I
I1
I
|
50-
RF
40-EcoRIA
30 -0
E
20 -i
Ig~~~~~~~~~~cR
It~~~~~~~~~~~~~
B2
Fraction
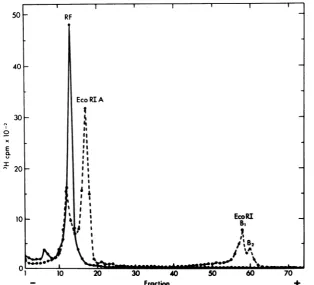
FIG. 1. Analyticalgel electrophoresis ofH-1RF DNA andEcoRI-digestedH-1 RF DNA. H-1 RF DNA
labeledwithP3H]TdRwasisolatedfromhamsterembryocellsinfectedwithwtH-1at37°C.AportionofRF DNAthatsedimentedat16.5S(cf. Fig.3,fractionA)wasdigestedin 50piofdigestion buffer for20 minat 37°Cwith 5 ,ulofEcoRI,andacontrol wasincubatedsimilarlywithoutenzyme. The reaction wasstopped
with 20 ,ulof2.5% sodiumdodecyl sulfate-50%glycerol-0.01% bromophenol blue-10 mMEDTA,and the
samplesweresubjectedtoelectrophoresis incylindrical gels of1.4% agarose.Inthiscasetheelectrophoresis
bufferwas90mMTris-borate(pH8.3)-2.5mMdisodiumEDTA,andelectrophoresiswasfor20hat35Vat
roomtemperature.Intheillustration, theresultsoftheseparategelsarecombinedtosimplify comparison.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.106.421.298.583.2]age of 78.4% of the [3H]TdRor32P when
32PO4
wasused as alabel, and the smaller fragments A.Haem
contained the remaining 21.6%. The dispersed 4 electrophoretic pattern of the smaller
frag-ments suggested several possible interpreta- 3
tions: (i) threeEcoRI cleavage sites, producing
one large fragment migrating as RI-A, one as A B C
RI-B2, andtwo asRI-B1 aslabeled in Fig. 1; (ii) 2 D E
twoEcoRI
cleavage
sites,
onewithinEcoRI-B%,
Fyielding
asmallfragment
lostfrom thegel
and 1 dmeLAAALGL1
converting
RI-B,
to RI-B2; and (iii) one EcoRI Acleavage
site,
with structuralheterogeneity
of\ xthe EcoRI-B fragment giving heterogeneous E
migration inthe gelelectrophoresis. u B.
EcO
RIThe firstalternativewas ruled out by deter- .
A'
mining the size of the EcoRI-B fragments. EcoRIA
Treatment of H-1RF DNA withthe restriction
endonuclease endo RNHaeIII of H. aegyptius .* ' A.^
produces
12unique fragments,designatedA to 10 103 -Eco RIBL, ranging in size from about 1,550 to 30 bp as
B.
.edeterminedbytheir contents of 32p as shown in . S s H
Fig. 2A. In areplicate gel (Fig. 2B), theEcoRI-H.* F
Bfragmentswere notresolved and migrated as
onebroadpeak at a sizeposition ofabout 1,050 Ha H
bp,
or 21% of the genome, inagreement
with 5_0
a 1the proportionofcountsrecoveredin all of the B Relative
Mobility
fragments (Fig. 2G). Thus, the B fragments Brepresent about 22% of the genome by size and composition and must be aheterogeneous
col-lectionof the samebasicfragment.
Theobserved8%differenceinrelative mobil- o 3 ity of
EcoHI-B,
and EcoRI-B2 with respect to _ FractionEcoRI-A in 3%acrylamidegelssuggests that a FIG. 2. Analytical gel electrophoresis of
32P-la-difference in size of about 50 to 70 bp would be beledH-1RF DNA digested by HaeIII or EcoRI.H-1
expected. Althoughtheproportionsof
EcoRI-B1
RF DNA labeled with 32po4 was prepared in the toEcoHl-B2were constant at 2:1independent of usualmannerthrough thesucrosegradient step. RFwhether the digestion was complete or notasin DNA infinal volumes of 50p1 wasdigested for 16 h
Fig. 1, afurtherexperiment to detect a possible at37°C with 5
pi
ofHaeIII (A)orfor 2 h at 37°Cwithsecond cleavage site near the first was con- 5 p of EcoRI (B).TheDNA wasanalyzed by
electro-ducted. Arelativelylargeamount of[3H]TdR- phoresis in
cylindrical gels
of
3%acrylamide-0.5%
ducled.
reA. larigesamoun
to[omH]tdR
agaroseataconstantvoltage of30Vfor16 h atroomlabeled RF DNIA was digested to completion temperature. The counts recovered in each fraction
with EcoRI.Thisdigest contained40,000 cpm of were converted to percent total counts, and the
EcoRI-B and was analyzed by electrophoresis number of base pairs for each fraction was calculated
for 3 h at50Vina3%acrylamide gel (datanot using a total of4,920 (cf. Fig. 4). The electrophoretic
shown). Using the mobility of the EcoRI-B frag- mobility ofeach fragment expressed as a relative
ment and a linear extrapolation- of mobility mobility in comparison to EcoRI-A fragment was
versus logarithm of molecular size as in Fig. plottedagainstthefragment size on alog scale in(C).
2C,
the3-hgel
wouldhavecontainedfragments The mobility ofEcoRI-B indicates a fragment size of2Ci
tharelative
3-hmgelwouldt
of9withrespecto about1,050bp
or21.3%of
thegenome.InFig.
2B,with a relative
mobility
of 9 withrespect
EcoRI-B contained 20% of the total 32P
recovered.
EcoHI-B and an estimated sizeof 2 to 4 bp. A The
HaeIII
fragments I-Lmigrated
off this gel.putative fragment of 50 to 70 bp would have
containedapproximately1%of the totalcounts extracts of H-1-infected cultures that the RF
per minute or about 2,000 cpm. No evidence for DNA peak has a fast-sedimenting shoulder
suchafragmentwasobtained. Evidenceinsup- (e.g.,Fig. 3). Similarly, gel electrophoresishas
port of the thirdalternative,namely, that there revealed a species of DNA migrating more
is oneEcoRI cleavagesiteandthat the EcoRI-B slowlythanRFDNA,e.g., fraction6in Fig. 1.
fragment is heterogenous, ispresentedbelow. To determine the molecular weight of this
H-1 dimer RF DNA. It has been observed DNA, the
[14C]TdR-labeled
viral DNAillus-routinely in neutral sucrose gradients ofHirt trated in Fig. 3 was pooledasillustrated, and
on November 10, 2019 by guest
http://jvi.asm.org/
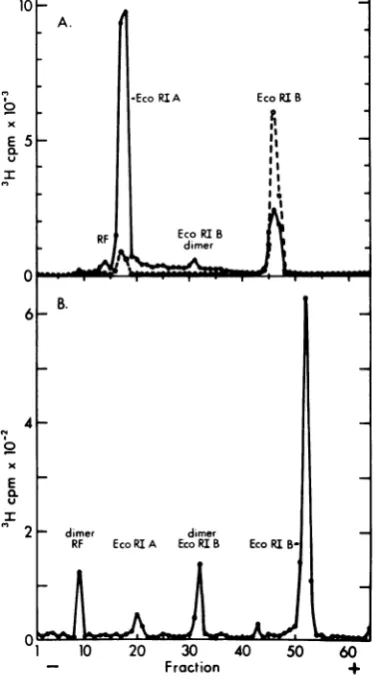
[image:4.501.252.444.51.412.2]B. .A . beleddimerwas
prepared
asabove andpurified
30
I-+--A-I
by preparative gel electrophoresis. A portion ofthis preparation was digested by EcoRI and
analyzed by electrophoresis in a 1.4% agarose
25 - gel(Fig. 5).Threefragmentswereobtained;the
largest had the electrophoretic mobility of
EcoRI-A, the second migrated with a mobility
20 - equivalent to 2,100 bp or twice the size of
o EcoRI-B, andasmaller portionwasat the
posi-x tionof
EcoRI-B.
This is theexpected
result foraE lS dimer RF DNA linked through the EcoRI-B
E 15 fragments, rather than in tandem. A similar
u l result was obtained after a digestion of a
mix-10 .lo} ture of equal parts of monomer and dimer RF
. DNA. The small amount of 3H recovered at the
position of EcoRI-B probably arose from
con-RI
I1 5 10 15 20 25 o \
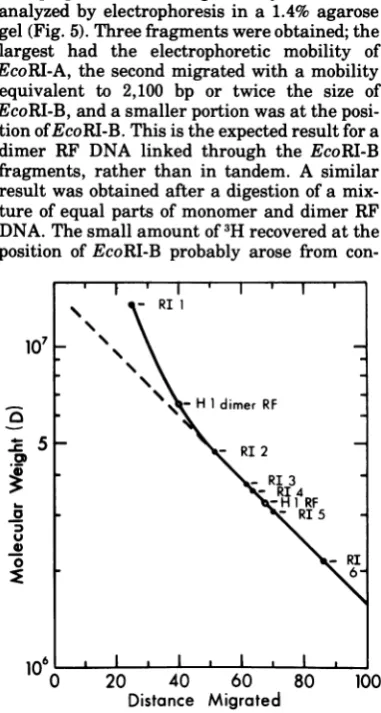
Fraction 9g
FIG. 3. Preparative sucrose gradient centrifuga- L - H 1 dimer RF tion of H-1 RF DNA. H-1 RF DNA was labeled with
['4C]TdR by incubating parasynchronous hamster .C 5 - RI 2
cultures infected with tsl H-1 at 39.5°C 12 to 16 h p.i. *j R
in medium containing FUdR (0.5 pg/ml) and RI 3
[f4C]TdR (1 pCilml, 57.1 mCi/mmol). ViralDNA , H1 RF
wasextractedbythe Hirt method and sedimentedin - RI5
agradient of 5 to 20% sucrose in 50 mM Tris (pH \
8.0)-i MNaCl-1 mMEDTA-0.2% Sarkosyl in the 0- RI
SW25rotorfor 18 h at 24,000 rpm. Fractions of1 ml 6
werecollected through the bottom of the tube,10-pl
aliquots were assayed for radioactivity, and the frac-tions were pooled as in the figure. The DNA was precipitated with 2.5 volumes of ethanol at -20°C for
16or more h.The precipitates were collected by cen- 106 ' I ' I ' I I
trifugation and washed once with 70% ethanol-50 0 20 40 60 80 100
mMTris (pH 7.5)-1.5 MNaCl-1 mMEDTA, and the Distance Migrated
DNA was stored in 10 mM Tris (pH 7.5)-10 mM FIG. 4. Determination of the molecularweights of NaCl-0.1 mM EDTA at -20°C.
H-1
monomer and dimer RF DNA. The molecular weights of H-1 monomer and dimer RF DNA were pool B, previously determined to contain some measured by coelectrophoresis of[f4C]TdR-labeledof the RF DNA of pool A, was subjected to H-1DNApreparedas in Fig.3 with the
[3H]TdR-coelectrophoresis with an EcoRI digest of labeled EcoRIfragments of X phage DNA.
[3H]TdR-[3H]TdR-labeled
bacteriophage X DNA in0.7% labeled XDNA was prepared as described(11) andagarose gels. The results are presentedasthe digestedtocompletion with100 U of EcoRIin a
vol-umeof100pi.Thereactionwas stopped by the
addi-log
molecularweight
versusdistance
ofmigra-
tion ofEDTA to 50 mM, and 20pdoftheXdigest andtion in Fig. 4, and the molecular weights de- a
5-pi
aliquot of the["4C]TdR H-1
DNA wereana-rivedfor thetwopeaks ofH-1DNAwere 3.26x lyzed by electrophoresis in duplicate in 1%agarose
106forH-1RF DNAand6.5 x 106for the faster- cylindrical gels as in Materials and Methods. The
sedimenting species. These values represent figure illustratesthe distance migratedand thelog
the average of two separate determinations, molecular
weight of
EcoRIXfragments
(a)andthewhich differed by less than 1%. It should be electrophoretic mobility ofH-1 RF DNA monomer
noted thatthe curve isnonlinearinthe regions and dimer
(0).
The molecular weight of the smallerwhere thefaster-sedimenting H-1DNA is rep-
H-1
DNA was calculated using a linearregressionwhresent.
Thes
D * constructed by the methodof least squares for logrespeted.ofa
This
erDN
Has
themoleculAr
weig molecularweight
as afunction
ofdistance
migrated
expected of a dimerofH-i RF DNA. for the XEcoRI fragments
2-6
(11).
The molecularToclarifyfurther the identity and structure weight of the larger
H-1
DNA was estimatedgraphi-of this putative dimer RF DNA, [3H]TdR-la- cally, since it lay on the nonlinear part of thecurve.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.60.286.46.293.2] [image:5.501.265.456.96.453.2]EcoRIA' ' ' '
'IF
cohesive termini of themonomer RF DNAdur-15 - E _ ing extraction and subsequent handling. To
de-termine whether all RF DNAcould anneal, a
portion of the 11-to 12-h RF DNApreparation
abovewas incubated in 50%formamidefor4 h at 400C before electrophoresis. No increase in
10_ dimerRF DNAoccurred.
or
.A more directtest for the possibility ofran-x - dom annealing of cohesive RF monomers was
E made
using
thedensity
labelbromodeoxyuri-u dine (BUdR). In this experiment, wt RF DNA
^5 - _ was
prelabeled
with[14C]TdR
and incubated30dimer
minwithFUdR,
followedby
30 minoflabeling
EcoRIB with [3H]BUdR in thepresence of FUdR. This
EcoRI B
0 10 20 30 40 50 60 20 A. + _10
Fraction +
FIG. 5. Analytical gel electrophoresis of an EcoRI 15- t
digest ofH-1 dimer RF DNA. Electrophoresis was in o L \
a1.4% agarose gel as in Fig. 1 except buffer E was x x
used for 16 h at 25 V at room temperature. DNA Elo_, 5 E
peaks were identified from the position of monomer u O- u
fragments on a replicate gel and their mobilities rela- I \ i tivetothe marker EcoRI-A.o
taminationof the dimer
preparation
withmon- 0 OomerRF DNA.
To determine whether the dimer DNA con- B. tained covalently linked dimer length strands,
[3H]TdR-labeled dimer or monomerand 32P-la-
10
beledmonomerRF DNA werecomparedinal- °
kaline sucrose gradients (Fig. 6). Virion DNA
tE
was sedimented in a replicate gradient (not X ls shown),thepositionof which is indicated by the Iarrow
(V),
and thearrow(D)
indicates theposi-
_tion of dimer length DNA expected using the /
equationsofStudier(25).Itcanbeseenthat the ° / t
majorityof bothmonomerand dimer DNAsare
genome length in the alkaline gradient and d
thus not covalently linked. Both preparations
show
fast-sedimenting
shoulders not seenwith01
5 10 15 20 2530
virion
DNA,which
we will consider in detailFraction
i-9below.
Thelw
relative proportions of dimer andmon- FIG. 6.
Centrifugationofdimerand
monomerH-1 RF DNAinalkalinesucrosegradients.Labeled H-1omer RF DNA synthesized at different times RF DNA was prepared as in
Materials
and Methods.postinfection (p.i.) were examined in wt H-1- The B fraction of the 3H-labeled
H-1
DNA shownininfected hamster embryo cells labeled with Fig. 3 was fractionated into monomer and dimer [3H]TdR11to 12 hp.i.,11to 12 hp.i. plus a 5-h fractions by preparative gel electrophoresis in 1%
incubation in the presence of unlabeled TdR, agarose. 32P-labeled monomer
H-i
RF DNA wasand 16 and 17 h p.i. The viral DNA was ana- addedto the 3H monomer (A) ordimer(B)fractions,
lyzed by electrophoresis in 0.7% agarose gels. and after the mixtures were adjusted to 0.2NNaOH,
Theproportionof viral DNAthatwasdimer RF they were sedimented in a 5 to 20% alkalinesucrose
DNAvaried little: 17%o for the 11- to 12-h
pJ
i gradient(cf.
A inFig.
3)for
6hat42,000rpmat4°C
in the SW50 rotor. A replicate tube contained 3H-labeling period, 26% for 11 to 12 h plus 5-h labeled virion H-I DNA, and its sedimentation
posi-chase, and 22% for the 16- to 17-h label. Since tion is indicated by the arrow V. The arrow Ddenotes
the majority of the dimers are not covalently the expected sedimentation position of dimer DNA
linked, they may be formed by annealing of relative to monomerby theequation of Studier (25).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.246.442.226.520.2]period ofdensity labelingwaschosentoproduce AIim
*.l
alabeling of onlyasmall portion of the total RF H Lightpool. Therefore, if dimers formed by random Hybrid Light
association ofmonomers, the majority of the W
V
dimerslabeledwith3Hwould containonlyone LO)
[3H]BUdR-substituted monomer RF DNA and ° x (
beless than hybrid density. On the otherhand, x 7
ifdimers replicate semiconservatively during E 5 \ E
thedensity labelingoriftheyaregenerated by u
afailure of
daughter
RFmonomerstosegregate
° 1after replication, 3H-labeled dimers would have - 135
hybrid density.
Dimer RF DNAwasprepared
, lbysucrosegradient centrifugation and bypre- *
parativegel electrophoresis, and the dimer and
monomer (sucrose
gradient only)
ugtin-n-
RF DNAswere analyzed
by
isopycnic
centrifugation
in - .50Cs2SO4
(Fig. 7A and B). About 60% of the3H- \Bronomer
mlabeledmonomer isof hybriddensity, whereas O
the density of the 3H dimer is intermediate 15 51*
between light and hybrid, and virtuallyno 3H > 4 \ 4
dimer isofhybrid density. This is compatible 0 - o t
witharandom annealing ofmonomerRF DNA x x14 E
asthemechanism of dimer formation and rules E 5 10
outafailure ofdaughter RFmonomersegrega- u °
tionas a meansof dimergeneration. However, -X
this result would also be obtained if dimers 5
replicated more slowly than monomers, such o
thata 30-mindensity labelisless thanorequal
to the replication time for dimer RF under I
these conditions.
_1
To examine this possibility, the experiment C 10 20 30 40
was repeated in asimilar manner, except the Fraction
period of density label was extended to60min FIG. 7. Isopycniccentrifugationofdensity-labeled
(not
shown). After the
1 hof
density labeling,
H-i
RF monomer and dimer DNA.H-1
DNA was mostof the 3H-labeleddimer DNA was of hy- labeled in parasynchronous NB cultures infectedbrid density; however, appreciable amounts of withtsl at 39.50C first with['4C]TdRfrom 12 to 15.5 thetotalmonomer RF DNA werecalculatedto hp.i. (FUdR,0.5pg/ml; [14C]TdR, 1 XCi/ml,57.1
have become heavy (13%) and hybrid (38%) mCi/mmol), followed by a 0.5-h treatment with
densities. Thus, the distribution of density la- FUdR alone, and then labeled with
[3H]BUdR
frombel inthedimerRF DNA wascompatiblewith 16 to 16.5 h p.i. (FUdR, 0.5 pglml;[3H]BUdR,2
a slower dimer replication process, random
XCi/ml,
10-6
M). Viral DNA was extracted by theannsalow
dimrF
DNA* Hirt method andfractionated
intomonomeranddi-annea
of RFDnA
merbycentrifugation
in
a neutral sucrosegradient
complex situation. and bypreparative gel electrophoresis in 1 % agarose.
Covalentlinkage of theV toC strandinthe Monomer and dimer DNA were banded to equilib-EcoRI-B fragment ofmonomer RF DNA. It rium inCs2SO4:(A) dimer RFDNA, (B) monomer
waspreviously observed that the RF monomer RF DNA.Centrifugationwasfor48 hat35,000 rpm
and dimer DNApeaks had afaster-sediment- at
4°C
in thetype 40rotor.ing shoulder in alkaline gradients, suggestive
of the presence of single-stranded DNA of guanine plus cytosine content that might fail to
greater than genome length. Tattersall et al. denature with heat under the conditions used
have presented evidence for the existence of forMVM RF DNA (23). Nevertheless, if such a
some RFDNAoftheparvovirus MVM (minute covalent cross-linkage is present in a portion of
virus of mice) that renatures unimolecularly theH-1RF monomer and dimer DNAs, then it
after heat denaturation, andsuggested that a could be isolated by using its rapid renatur-covalently closed turnaround may exist in RF ationrate. This was tested by alkaline
denatur-DNA (27). We have obtained evidence for a ationof wt monomerH-1 RFDNA followed by
region at one end of H-1 RF DNA with a high rapidneutralization and immediate application
melting temperature implicating a very high to a BDC column. Double-stranded DNA was
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.269.458.49.399.2]H-1
TABLE 1. Akaline denaturation-BDCchromatography of H-1 RF DNAa
Cycle1 Cycle2
Procedure
cpm % Total cpm %Total
Washes 1,295 3,108 1.7
1 MTBS-total 261,560 18.5 128,560 73.1
Caffeine-total 1,151,340 81.4 44,300 25.2
a [3H]TdR-labeled wt H-1 RF DNA was denatured by 0.2 N NaOH, neutralized at
0°C,
andchromato-graphed on a column of BDC as in Materials and Methods. The DNA eluted with 1 M TBS was collected by ethanol precipitation, and theprocesswasrepeatedincycle2. Values shown are total counts per minute recovered and theirrespective percentages.
selectively elutedwith 1 M TBS and DNA con- oldandnewmolecules. This DNA was fraction-taining single-stranded regions recovered by ated by alkaline denaturation-BDC
chromatog-elution with 2% caffeine. The results are sum- raphy, followed byanalkalinesucrose gradient
marized in Table 1. When the double-stranded as above. The results in Fig. 8B show that the
DNA was putthrough asecondcycleofalkaline 3H/1'4C ratios were lower for the dimer length
denaturation-BDC chromatography, the per- DNA than for the unfractionated monomer RF
centagerecoveredasdoublestrandedincreased DNA, 0.29 versus 1.1. Thus, newlyreplicated
from 18.5 to 73%. A portion of this DNA was monomerRF molecules (within 30min)contain
then analyzed by alkalinesucrosegradientcen- less of the covalent linkage than the general trifugation (Fig. 8A). A majority of the DNA pool,asrepresented by the[14C]TdR-labeledRF
(68%) sedimented at the expected position of DNA.
dimer length singlestrands, in constrast tothe Covalentlinkage of Vto C strand in dimer [14C]TdR-labeled tsl RF DNA monomer RFDNA. Experimentssimilar to those used to marker. The final recovery indicated that at show linkage ofV toCstrandsinmonomerRF least 9%of the RFmonomercontaineda cova- DNAwerecarriedoutwith[3H]TdR-labeledwt
lentlinkagebetween the Vand C strands. dimerRF DNApurifiedby gelelectrophoresis.
The localization of the V-C linkage was de- Afteronecycle ofalkalinedenaturation, 24%of termined by digesting RF DNA with EcoRI the 3H was recovered as double stranded in before alkalinedenaturation andBDCchroma- BDC chromatography, and 87% of this DNA tography. The double-stranded DNA so ob- sedimentedasdimerlengthin an alkaline su-tained was analyzed by electrophoresis in a crose gradient (not shown). This represented
1.4% agarose gel and compared with the total about 21% of the original dimer preparation.
digest (without selection for the rapidly rena- The gel electrophoresis of this dimer prepara-turing DNA) inareplicate gel. The resultsare tion after EcoRI digestion is shownin Fig. 5.
illustratedtogether forconvenience inFig. 9A. This DNA contained about 5%
EcoRI-B
mon-TheEcoRI-Bfragmentwas clearlyselected for omer would be dimer length
single-stranded
by the procedure isolating rapidly annealing 20% contamination with monomer RF DNA. DNA, indicating that the covalentlinkageis in Sincewehaveseenthat about10%of the mon-the EcoRI-B fragment. A control experiment omer would be dimer-length
single-stranded
showedthat BDCchromatography ofanEcoRI DNA, this would constitute only2%of the total digest of RF DNA without alkalinedenatura- dimer preparation, so that it can be deduced tion resulted in the retention ofabout 30% of that some of the dimers contained covalently theDNAafter 1MTBSelution,suggestingthe linkedVand C strands.
presenceofsomesingle-strandedregions,butit To test whether the covalent linkage inthe
was notselective for eitherfragment. This sub- dimers is at the internal left end of the
mon-ject will beconsideredin moredetailin asubse- omer, and probably at the site oflinkage
be-quent sectionof this
study.
tweenthemonomers, dimerDNA wasdigested
Ifthe covalent linkage of V to C strand in with EcoRI and then subjected to the alkaline
monomer RF DNA was involved in the initia- denaturation-BDC chromatography procedure.
tionofreplication of RF DNA or progeny DNA The rapidly renaturing DNA was
analyzed
bysynthesis, then
rapidly
labeled molecules gel electrophoresis (Fig. 9B). Itwasfoundthat would be enriched in this property compared theBfragmentwasisolatedina40-foldmolarwiththetotal collection of RF DNA molecules. excesscomparedwith the Afragment and thus
The [14C]TdR-prelabeled RF monomer DNA containedthe V-Cstrandlinkage.It isunlikely
used for the
[3H]BUdR
densitylabel in Fig. 7B that a covalent linkage occurred during thewas a ready source of DNA for comparison of Hirt extraction and subsequent
handling,
soon November 10, 2019 by guest
http://jvi.asm.org/
A. crudelocalization of the
origin
ofreplication.
Inthese experiments, tsl H-1 was used at the
restrictive temperature to inhibit progeny DNA
20 - > \ ,0, _
synthesis.
This method has beensuccessfully
x 8 | , z used by Danna and Nathans for simian virus 40
E (7) and
by
Godson forOX174
(10). The firstproblem to be overcome with the linearH-1 RF
Ut
I I0
I
'10~ ~ q10
~~~~10I
~~~~~~~~~~~A.
X 0
* -Eco RIA Eco RI B
B. dimer monomer Eo
II I
60 30 Eo
I
~~~~~~~~~~~~~~~~~~~~~~Eco
RI B9 ~~
~~~~~~~~~~~~~RF
~ darner140 20 A
9, ~~~~~~~~~~0
x x
E E B.
FIG. Alaln -ursrdetcnrfgto
a-0~~~ ~ ~~~~~~~.
020 01
4
0~~~~~~~~~
C1
~~~10
20 30 EFraction 9-ga
FIG. 8. Alkaline sucrose gradient centrifugation 2 -dimer
dirner
of rapidly renaturing RF DNA. (A) H-1 RF DNA 2 RF Eco RIA Eco RIB EcoRIB monomer was selected for its ability to reanneal
rap-idly by two cycles of alkaline denaturation-BDC chromatography (Table 1). The DNA in the 1 M TBS
eluate
of
thesecondchromatography
wasadjusted
to aJLAb2Ahl
A0.2 NNaOH andsedimented in agradient of5 to
20% sucrose in 0.2 N NaOH-0.8 M NaCl-1 mM 1 10 20 30 40 50 60
EDTA for 16 h at 32,000 rpm at 4°C. A marker of - Fraction +
[14C]TdRmonomer RF DNA obtained frompoolA of FIG. 9.
Analytical
gelelectrophoresisofEcoRIdi-Fig. 3 was included. (B) H-1 RF DNA monomer gsso RFDAbyeoeandaferoakaie
dEntRa-prelabeled with ['4C]TdR (12 to 15.5 h p.i.) and gestsofRFDNAbeforeand
afiter
alkalinedenatura-pulse-labeledwith[H]BUdR (16to16.5 hp.i.) as tion-BDC chromatography. (A) RF DNA was
di-puselbeldwi
h.
t7,HBUd
(16 to16. hyci)
as gestedto completion withEcoRI and thenfraction-describe in Fig.7, sctdmtog
onehycleo
ated byalkaline denaturation-BDCchromatography.raplk
annalineDN
denaturation-BDC chr1MaTo
Tewa AsampleoftherapidlyannealingDNA inthe 1 Mrapidlyannealing DNA inthe1 M TBS eluate was TBS
eluate
and an aliquot removed before alkalinecentrifugedinanalkalinesucrosegradientasin(A). denaturation-BDC chromatography were analyzed
separately in replicate gels of 1.4% agarose.
Electro-thisresult strongly suggests that the dimer RF phoretic conditions were 17 h at 25 V at room
temper-DNAexists in vivo. ature. Results ofthe replicate gels wereplotted
to-Evidence thatthe Bfragment is a terminus gether for convenience: before alkaline
denaturation-in RF DNA
replication.
By
analyzing
the dis- BDC(), andafter alkalinedenaturation-BDC
(0).
trinution
repltion.dBe
zIAang
te- (B) DimerRF DNA was digested with EcoRIandtribution of [3H]TR in the EcoRI-A and B fractionatedbyalkalinedenaturation-BDC
chroma-fragments of H-i RF DNA that have
just
com- tography. The1 M TBS eluate was analyzed in a gelpleted replication during a short incubation in of1.4% agarose as in (A),except electrophoresis was
thepresence of[3H]TdR,it is possible to make a for 20 h at 25V.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.501.65.268.56.410.2] [image:9.501.268.456.158.501.2]DNA is to devise a method of isolating the .
moleculesthat have just completed replication A.
during the short pulse of [3H]TdR. In these IMTBS 0-27 Caffeine molecules, the specific activity of TdR at each
position in the molecule will increase as the
distance of that position from the origin in- o
creases. In comparison, RImolecules will have x
adeclining specific activity in TdRas the TdR E 5
-positionfrom theorigin increases.For this pur- l
pose, I have usedchromatography on columns I
ofBDC, which separates double-stranded DNA
fromthe more tightly bound DNA that contains °
single-strandedregions (2, 27). Thismethodor °
a similar one using benzoylated napthoylated i DEAE-cellulose has been used for the
separa-tion of viral RI molecules from mature DNA for 0
lambda (13),adenovirus(26),
OX174
(21), MVM B.(27) and, most recently, herpes simplex virus I-I - 15
(22).
Rapidly labeled H-1 RF DNA was found to bind preferentially to BDC and to require caf- °
feine for elution, as has been shown for other x 5 - 10 x
E E
examples of replicative intermediates cited Q lI
above. Rapidly labeled viralDNA wasprepared I
bylabelingtsl-infectedNBcultures,afterincu- "V
bationat39.5°C, for5minat22°C, orfor5min
50
or 15minat39.5°C, with [3H]TdR before
extrac-tion of viral DNA. A
uniformly
labeled RF DNAwassimilarly prepared by
incubating
the 0cultures from 12 to 16 h p.i. in FUdR and 1 2 3 1 10 20 30 40 [14C]TdR (cf. fractionA in Fig. 3).Afterapre- Fraction
liminary purification in neutral sucrose gra- FIG. 10. BDC
chromatography of rapidly
labeled dients, the [3H]DNA that sedimented as mon-H-1
RF DNA.ParasynchronousculturesofNBcellsomer RF DNA (but which may also include were infected with tsl H-i at 39.5C and treatedwith
early RI molecules), wascombined with apor- FUdR for 30 min, 14.5 to 15 hp.i.At 15 h p.i. the tionof['4C]TdR-labeledRF DNA andwasthen cultures were labeled for5or 15 min with [3H]TdR chromatographed on BDC columns (Fig. 10). (20 p.Ci/ml, 50Ci/mmol) in thepresence ofFUdR.
Thepercentage of the labeled DNA that eluted Viral DNA
sedimenting
as monomerRF DNA waswithcaffeinewas 95%forthe22°C,5-min label prepared in the usual manner and was chromato-(not
shown),
80% for the 5-min 3H label at graphed with a['4C]TdR
marker (cf. Fig. 3) on39.50C,
61%for the15-min 3Hlabel,and38%for columns ofBDC-cellulose using elutionsof2O
ml of1thethe4-h-h[14C]TdR-labeled[4C]TR-laeledDNA.ThusDNA. Thus,
therap-caffeine
theMrap-
M TBS andin 1 Ma 20-mi linearTBS. One-millilitergradient
fractionsof
0 to 2%were
idly labeled RF DNA was preferentially re- collected and
50-pi
aliquots were assayedforradio-tained after the 1 M TBSelution, even though activity; (A)5-minlabel, (B)15-minlabel. The1 M
the more rapidly sedimenting RI molecules TBS and 2% caffeine eluates were pooled as illus-were already partially excluded by the zonal trated and the DNA wasprecipitated with 2.5
vol-centrifugation
inneutralsucrose. umesof ethanol at-20°C
overnight.The rather high fraction of the
[14C]TdR-la-beledDNAthat remained boundafter salt elu- wouldbe retained
preferentially
after the 1 M tion deserves further study. Electron micro- TBS elution. This wastestedby
digesting
3H-scopevisualizationof thisfraction showed that labeled RF DNA
(4-h labeling period)
withonlyabout 25% of the molecules were RI mole- EcoRI. The
digest
was extracted withphenol,
cules (23), so that one explanation would be
precipitated
withethanol,
redissolved in0.3Mthat these RF DNA molecules have large
TBS,
andchromatographed
on BDC.Thirty-enough
single-stranded
DNA regions to bind seven percent of the total 3H eluted withcaf-BDC in 1 M TBS, as recently reported for feine. Portions of the 1MTBSand 2% caffeine
herpes simplex virus (22). Ifthis were true, eluate were analyzed by gel electrophoresis,
then theseregions may beuniquelylocated, so and there wasnopreferential binding
ofEcoRI-that afterEcoRI cleavage,oneof thefragments A
compared
with -Bby
the BDC(Table
2).on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.501.252.446.50.379.2]TABLE 2. Compositionof BDCchromatographic visualization by electron microscopy.
There-fractions ofEcoRI-digestedH-i RF DNAa fore, the saltfractioncontains the
nonreplicat-%Composition ing molecules, even though some are retained
Elution with: by thecolumn for anunknownreason.
RF EcoRI-A EcoRI-B Returningnow to the ts 1 RF DNAlabeled for
1 MTBS 11 67 22 15minat39.5°C and fractionated by BDC
chro-2%Caffeine 23 60 17 matography (cf. Fig. 10), the1 MTBS fraction
aH-1 RF DNAlabeled with 3H wasdigested to and 2%caffeine fraction wereseparately
precip-near completion with EcoRI. The digest was ex-
itated
with ethanol anddigested
WithEcoRI,
tracted with phenol andprecipitated with ethanol. and the digests were analyzed by gel
electro-Theprecipitate wasredissolved in 0.3 M TBS and phoresis (Fig. 12). Additional
[14C]TdR-labeled
fractionatedon acolumn of BDC. The 1 M TBS and marker RFDNA,not fractionatedbyBDC,was 2%caffeineeluates wereprecipitatedwithethanol, added before the digest to improve the 3H/14Cand the DNA was analyzed by electrophoresis in ratios (theabsolutevaluesforthe 3H/14C ratios
1.4% agarose gels. The values shown represent the of the twocolumn fractionswere notadjustedto
percent composition of RF, EcoRI-A, and EcoRI- be equal). The 3H/14C ratios for the 1 M TBS
Bin each eluate. eluate EcoRI-A and -B fragments prepared
from this mixture were 3.32 and 4.42, respec-The 2% caffeine eluate was enriched for the
undigested RF DNA.
If the postulated single-stranded regions A' I were distributed within both EcoRI-A and
EcoRI-B, then RF DNA in the caffeine eluate shouldshowadecreaseinsedimentation
veloc-ity inalkalinesucrosegradientsincomparison
10
to the 1 M TBS eluate. In this experiment, uniformly labeled monomerRFDNAwas frac-tionatedonaBDCcolumn, and theDNA inthe 1MTBSeluate andthe 2%caffeinewasprecipi- 5
tatedby ethanoland thensubjectedto
sedimen-'l
tation inalkaline sucrose gradients. Amarker
of
[14C]TdR-labeled
RF DNA (Fig. 3) was in- xcluded, and its modal sedimentation is indi- E °
catedbythearrow (Fig. 11). Althoughthe caf- X 20 -B.
feine eluatewasenrichedslightly for DNA sed- I
imenting slower thanthegenomelength DNA, 1
both preparations were largely intact dimer
15
length ormonomerlengthsingle strands. This indicatesthat
internal
single-strandedgaps arenotcommon in the RF DNA of the2% caffeine
eluate.
10
_Another possibility is that the DNA is re-tained nonspecifically andnotbecause internal single-strandedregions exist. Ifthisisthecase,
rechromatography
of the 1 M TBS eluate 5shouldagain produce a sizable fractionretained
after 1 M TBS elution. This experiment was
doneusing aportionofthe first 1 M TBS eluate
of
Fig.
11, whichrepresented
one-tenth of the01
1020
30
total DNAused for the first BDCcolumn. The
result obtained was that 24% ofthe 1 M TBS Fraction g
fraction remained tightly bound and eluted in FIG. 11. Alkalinesucrose
gradient
centrifugationthe 2% caffeine eluate. This is a smaller per- of BDCchromatographic fractions of H-1 RF DNA.
centage than duringthe first chromatography
wt
RF DNA labeled4 h with[3H]TdR
wasfraction-(37%),
but it does indicate arelatively
high
ated by BDC chromatography using batch elutions of (37%, buevel.of
nonspeclinc
iteinic
does
retentIon
ten
aafter
rltel
1rM
gT
irTB
1collected by ethanol precipitation and sedimentedMTBSand 2%caffeine. The DNAfractions wereaselution. In conclusion, RI molecules are re- alkalinesucrosegradientsas inFig. 8. The marker
tained on BDC columns after salt elution on DNA of[14C]TdR-labeledRFsedimentedatthe
ar-the basis ofthe kinetics oflabelingand direct row; (A) 1 M TBS eluate, (B) 2%caffeineeluate.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.501.263.455.266.582.2]IA.
ERIA
labeled DNA, migrating faster than EcoRI-A,5-A.EcoRIA arose as aresultof the BDC chromatographyor
4-20 R h 5 EcoRI
digestion,
thebehavior ofrapidly
labeled
4-20 RF DNA and RI DNA in
gel
electrophoresis
3 -15 cJ > \was examined.
3H-labeled
tsl RF DNA wasx;
3-31~prepared by
a5-min incubation with[3H]TdR
I _E at
39.5°C
and fractionatedinaneutralsucrosez2 a Fg ubsfo h
".ffi , _0 gradient
(Fig.
13). Tubes from the gradient. EcoRIB were pooled to make threefractions, A, B, and
1
C,
as illustrated. After precipitation with---
ethanol,
each fraction wassubjected
toagaroseo_ gel electrophoresis in the presence of a
10 30
[14C]TdR-labeled
RF preparation containingmonomer and dimer RF DNA (Fig. 14). Frac-tionAshows[3H]DNAexhibitingahigh 3H/14C
-15 20 ratio migrating in advance of monomer RF
IO
s I DNA(Fig.
14A).
Therefore,
therapidly
labeledx -o < v 9 10 DNA
migrated
ahead ofRF DNA before BDCa- chromatography and EcoRI digestion, with a
I o 1 _
°0
mobility equivalent to a duplex DNA 10 to 15%-5a 1' smaller than RF DNA.
After
EcoRI
digestion-o
Q ! zthis DNA migrated ahead of EcoRI-A, and itsmobility
increased about the amountexpected
10 20 30 40 50 if EcoRI has cleaved theEcoRI-B region from
Fraction +
FIG. 12. Analytical gel electrophoresis of EcoRI
digests of RF and RI DNA labeled15 min.H-1 tsl 15 I
RFDNAlabeled15minwith[3H]TdRwas fraction-ated by BDC chromatography as in Fig. 10. The DNA in the1 MTBS and2%caffeine eluates were
separatelydigested with EcoRI after the addition of C B A
analiquot ofuniformlylabeled[14C]TdR-labeledtsl I
RF DNA and analyzed by electrophoresis in 1.4%
agarosegelsasinFig.9.Electrophoresiswasfor17 h 10_
at25Vat roomtemperature;(A)1MTBS eluate, (B) ||
2%caffeine eluate. 0
x
tively. Thus, if the 3H/ 4C ratio ofEcoRI-A is E
normalized to 1, the ratioforEcoRI-B was 1.33.
This increase is
expected
if EcoRI-B does not 5contain the origin ofreplication. The caffeine
eluate shows 3H/14C ratiosof7.94 and8.27 for
EcoRI-A and -B, respectively. The normalized
value for the
EcoRI-B/EcoRI-A
was 1.04. In thecaffeine eluate the EcoRI-A
fragment
did not show theexpected
higher3H/14C
ratiocompared
0with EcoRI-B, but it can be seen in Fig. 12B 1 10 20 thatasmall
portion
oftherapidly
labeled[3H]- Fraction g DNA with high 3H/14C ratios (as high as 40) FIG. 13. Preparative sucrose gradient centrifuga-migrated aheadof EcoRI-A. This DNA was not tion of rapidly labeledH-1 RF DNA.Parasynchro-included in the average ratio of 7.94 cited for nous NB cultures infected with tsl H-1 at 39.5°Cwere
EcoRI-A above. treated 14 to15 hp.i. with FUdR (0.5 pglml) and
It would beexpected that RI moleculeswould labeled at39.5°Cwith [3H]TdR (50 pXi/ml,20
Cil
migrate moreslowly than linear RF DNA due mmol, + FUdR 0.5 pg/ml) for 5 min. Viral DNA
to their larger size, nonlinear configuration, was extracted as in Materials and
Methods
andsedi-andby analogywith the resultsreportedfor RI mented ina
gradient
of5to20%
sucroseasinFig.3.moleculeofsimianvirus4028. Ontheot Fractions of1 ml were collected, and 20 pi was
moleculesof simian virus40 (28). On the other assayed for
radioactivity
directly,accounting for the hand, single-strandedregions andgreater flexi- acid-soluble[3H]TdR seen in fractions 23 to 28. Se-bility might accelerate their electrophoretic lectedfractions were pooled as illustrated, and themobility. To determine whether this rapidly DNA was collectedby ethanolprecipitation.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.501.51.242.55.318.2] [image:12.501.255.450.326.542.2]A.
d;mRF RF
0
x~~~~~~~
O
65
15B.
~~~~~~~~~dimer
RF RF'o
C4 I
M I,
9t
2t5
C.
1 10 S S Z 9 K
- ~~~~~Fraction +
FIG. 14. Analyticalgel electrophoresis of rapidly labeledH-iRF DNA.H-iRFDNA waslabeled for 5mi
at39.5°Casin Fig. 13. The DNA fractions pooled as A, B, and C from the preparative sucrose gradient ofFig.
13 were analyzed in 0.7% agarose gels.A marker DNA of['4C]TdR-labeled mnonomerand dimer RF DNA (pool B of Fig. 3) was added before electrophoresis. Pools A, B, and C are illustrated in (A), (B), and (C), respectively. It should be noted that thealiquot used for A was 10% of the total, and B and C were 25% of the total. Gel electrophoresisof the total DNA of apreparativesucrose gradient equal to a composite offractions A + B + C of a tsiRF DNA labeledsimilarly for 5minat22°Cwith[3H]TdR is shown in(D).
these molecules. This suggests that this DNA the following papers of this series.
contains a high specific activity of 3H in the Gel electrophoresis of fractions B and C of
EcoRI-A portion of the molecule, which pre- Fig. 13 is shown in Fig. 14B and C. The
pat-sumably contains the origin ofreplication. Ex- terns revealed predominantly dimer RF DNA
periments to demonstrate this more conclu- and some [3H]DNA exhibiting high
3H/"4C
ra-sively using other restriction endonucleases are tios migrating more slowly than dimer RF.
inprogress, anda similarconclusion has been A DNA
preparation
from tsl H-i-infectedreached by electron microscopy, asdescribed in
min
cells that had been labeledfor
5 at22°C
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.501.120.397.64.501.2]REPLICATION TERMINUS OF 707
wasfractionatedon aBDC columnasdescribed vatedslightly by the 30 to 40%contaminationof above, and the caffeine-eluted fraction was the2%caffeine eluate with the 1MTBS DNA banded by isopycnic centrifugation in a CsCl aspreviously discussed. The expectedhigh 3H/ gradient, precipitated with ethanol, and then 321pratio for EcoRI-A may be absentbecausethe subjected toelectrophoresis in a 0.7% agarose RImolecules are selectively lost bythis
proce-gel (Fig. 14D). Thiselectrophoreticpattern rep- dure. The sucrose gradient fractionused here
resents an approximate compositeof Fig. 14A, excluded DNA migrating faster than monomer
B, and C. Thus, even with relativelyshort la- RF DNA. Also, examinationof the 2%caffeine
beling times the majority of the label has the eluate byelectron microscopyrevealed that RI mobility of eitherthe RFdimeror monomer. molecules were not the majorformpresent (23). Origin forprogeny DNAsynthesis is inthe Anotherfactorcomplicating theanalysis of the EcoRI-A fragment. In the preceding section, 2% caffeine eluate isthat [3H]DNA represent-evidencewaspresented which implied that the ing one-third of the total of the fraction and origin for semiconservativereplication ofts 1H- with thehighest3H/32Pratio(38.25,normalized
1RF DNA is in theEcoRI-A fragment. Since it to EcoRI-A) did not migrate as EcoRI-A or
was found earlier (17) that about 80% of the
EcoRI-B,
butwasinthefirst fraction of thegel.
labeled BUdR incorporated bywtRF DNAlate
Why
this DNA didnotmigrate
into thegel
is ininfectionat370Cwas intheVstrand, it can unknown. In summary, the distribution of la-be estimated that approximately 60% of the bel innewly completed
RFDNA,
as repre-totalincorporationis asymmetricV-strandsyn- sentedby the3Hlabelinthe 1MTBSfraction,
thesis. Thisprobably represents progenyDNA is notuniform and hasahigher
specific activity
synthesis by displacement of the preexistingV in the EcoRI-B fragment thanin theEcoRI-A strand from RF DNAby the nascentVstrand. fragment.
Thus,
thisfragment
appears toin-Using theexperimentaldesign of the preceding cludethe terminusforbothRFreplication and
section, the distribution of
[3H]TdR
in newly progeny DNA synthesis. Thecomplementary
completed wt RF DNA wasused todetermine analysis ofRI molecules asrepresented by the the polarity of synthesis and thusacrudeesti- 2% caffeine eluate is complicated by several
mate ofthe location of the origin for progeny factors andis notconclusive for the localization
DNAsynthesis. ofthe origin.
Parasynchronous NB cultures infected with
EcoRI-B2
fragment contains theturna-wtH-1 at
370C
werelabeled for 5 min at37°C round. The left end of H-1 RF DNAhas beenwith [3H]TdRin thepresence ofFUdR after a shown to be heterogenous in its structure, as
15-min preincubation with FUdR. The
mon-omerRFregion of the neutralsucrosegradient
of the Hirt-extracted viral DNA wasfraction- TABLE 3. Distribution of nascent DNAin wt RF ated by BDC
chromatography
and 10% of the DNAaapplied DNA was eluted with 1 M TBS. This
DNAand the2%caffeine eluateswerecollected 1 M TBS
2%
Caffeine asbefore anddigested
with EcoRIinthepres- Prepn3H/32P
Normal-
3H/32P
Normal-enceofuniformly
labeled 32Ptsl RF DNA, and ized izedthe digestwas
analyzed
by gel electrophoresis Gel top 7.65 38.25as in Fig. 11 (not
shown).
The 3H/32P ratios RF 0.457 3.07wereaslistedinTable3.In the1MTBSeluate, EcoRI-A 0.149 1.0 0.200 1.0 the EcoRI-B
3H/32P
ratio was 7.4-fold higher EcoRI-B 1.107 7.43 0.512 2.56 than the ratio for EcoRI-A. There were only wtRF DNAwaslabeled with[3H]TdR for5mm
small differences (no more than 12%) in at16 h p.i. at
370C
(50 ,uCi/ml; specificactivity, 20[3H]TdR/32
P ratios for EcoRI or HaellI frag- Ci/mmol). The DNAsedimenting
in a neutralsu-ments when the [3H]TdR-labeled RF was la-
crose
gradientasmonomer RF DNA (e.g., fractionbeled for a4-h
period.
Therefore,
thedistribu- A, Fig. 3) was fractionated by BDC chromatog-tionof TdRinRFDNA isnotsufficiently
asym- raphy.The 1 M TBSand 2% caffeine fractions weremetric to affect these results. This suggests
precipitated
with70% ethanolat-220C,
collectedby
that EcoRI-B is a terminusforwtDNAsynthe- centrifugation, and dried in vacuo. The DNA was
sis. redissolved in 50
,ul
ofEcoRIdigestion buffercom-Thedigestof the2%caffeine eluate was com- bined with a portion of 32P-labeled RF DNA and
plete,
and the3H/32Pratiowasagainhigher
fordigested
for2 h at37°C with 100 U ofEcoRI. Thedigest
wasfractionated by gel electrophoresisasinEcoRI-B (0.512) than for EcoRI-A (0.200), but
Fig. 12.
The 3H and32P
for each DNAspecies werethe difference was not as greatas for the salt- summed, and the 3H/32P ratios were calculated.
Val-elutable RF DNA, 2.5 versus 7.4
(normalized
uesshownare 3H/32P ratios and theratiosnormal-values). The ratio for EcoRI-B would be ele- ized to theratio for Eco RI-A.
on November 10, 2019 by guest
http://jvi.asm.org/
manifested by twoforms of the EcoRI-B frag- tweenthetwoelectrophoretic species for
EcoRI-ment, i.e.,
B1
and B2. We have also presented B wereduetodifferencesin sizeandnotstrand-other evidence for structural diversity inthis edness,sinceneitherB1norB2 wasremoved by
region: a covalent linkage of V strand to C BDC chromatography. TheEcoRI-B fragment
strandin 10%of the RF forms and self-cohesive- with the covalent Vto Clinkage had the
elec-nessof the leftendasmanifested by dimeriza- trophoretic mobility of the smaller
EcoRI-B2
tion. When the distribution of [3H]TdR in fragment. The twotypesofEcoRI-B
fragments
newlyreplicated RF DNA wasanalyzed,itwas were produced bya single cleavagebyEcoRI,
observed inbothexperiments that the ratio of andasimilar result has beenobtained with the
3Hto theuniformlabel of14Cor32p washigher H. aegyptius restriction endonuclease endo
forthe slower-migrating portion of the EcoRI-B R HaeII. This enzyme also cleaves the mole-fragment peak corresponding to B1- than for cule once near the EcoRI site (0.20 genome
EcoRi-B2 (cf. Fig. 12).The slightly faster migra- length from theleft end), producingtwo
HaeII-tionof the"snap-back" EcoRI-Bversusthetotal B fragments (unpublished data). Since both
EcoRI-B in Fig. 9 suggeststhatit migrates as
EcoRI-B1
and -B2 were found withtsi
RF asEcoRI-B2. A more definitive analysis of these well as wtRF, both fragmentswere
preserved
findingswascarriedoutbyadirectcomparison
during
thecloning ofts1. It isprobable that the ofanEcoRI digest of 32P-labeled H-1 RF DNA EcoRIfragments differ by about 50 to 70 bpin before and after alkaline denaturation-BDC length, asthedifferenceintheir electrophoretic chromatography by slab-gel electrophoresis mobilities suggests, or they may have some(Fig. 15). It isclear thattherapidly renaturing more complicated structural heterogeneity,
EcoRI-Bfragment migrates asEcoRI-B2.Inad- such as proposed for AAV DNA (8). Probably
dition, digestion of dimer RF DNA
produced
the simplestexplanationisthat there isa turn-the expected dimer EcoRI-B fragment, which aroundorhairpin atthe left endthatisabout shows a singleelectrophoretic
mobility.
Frac- 50 bp in length in the B2 fragment, asillus-tionation of theEcoRI-B digest on aBDC col- trated in Fig. 16 and, in the
B1
fragment, it isumnwasalso donetodetermine whether either extended by a full-length complementary fragment hada
significant single-stranded
re- strand, but this remains to be confirmed.gion,andelectrophoresis of the1MTBS eluate Analysis ofH-1 intracellular viral DNA by
shows that both
EcoRI-B,
and -B2arepresent. agarose-gelelectrophoresishas shown that H-1Therefore, it canbeconcluded thatnewly repli- RF DNA exists in two forms, a monomerand a
cated RFDNAmolecules contain the
configura-
dimer. The identity of the larger species as a tion characterized by anEcoRI-B,
fragment dimer RF wasmade by establishing its molecu-and that these molecules are probably con- larweight astwicethat ofmonomer RF DNAverted to the foldback configuration with or and by the specificity of cleavage of the
mole-without the covalent linkage of the V to C cule by EcoRIendonuclease. H-1 monomer and strand afterward. Thedifferencesin
electropho-
dimer RF DNA molecular weights weremea-reticmobilites of
EcoRI-B,
and-B2suggestthat sured by agarose-gel electrophoresis using thethe foldback region is about 50 to 70 bp in EcoRIfragments2through6ofbacteriophage X length, assuming thattheyareboth linear du- as markers of known molecular weights (11).
plexes. Monomer RFDNAhadan estimatedmolecular
DISCUSSION
weight
of3.26 x 106,and the dimerwasfoundtoDISCUSSION
be a "tail-to-tail" linkage of monomer RF DNA Thelinear duplexRF DNA oftheparvovirus attheir leftends rather than atandem union. H-1replicatessemiconservatively andprovides Thiswasrevealedby digestion of the dimer RFthe template for the asymmetricprogeny DNA DNA with
EcoRI,
and the EcoRI fragmentssynthesis (17, 18). Thelatterprocess was found obtained included twoEcoRI-Amoleculesanda
torequire at least one of thecapsid proteins by new fragment equal to twice the molecular
analysis of temperature-sensitive capsid pro- weight of EcoRI-B. Sedimentation of dimer RF
tein mutants of H-1 (19). In this study, the DNA in alkaline sucrosegradients showed that
cleavage of H-1 RF DNA by the bacterial re- most of the DNA sedimented as monomer
strictionendonucleaseEcoRI is shown to occur length single strands. Thus, the dimers were
at a unique cleavage site 0.22 genome length inferred to be linkedby hydrogenbonds in the
from theempirically designated left end of the self-cohesiveterminiatthe left end of monomer
molecule. The smaller EcoRI fragment B was RF DNA. Incubation of RF DNA in50%
form-heterogeneousin electrophoreticmobility, and amide for 4 h at 37°C resulted in a slight
de-anumber ofalternative structures were shown crease in thepercentage of the preparation that
to exist in this fragment. The differences be- wasdimerlength, revealing that the majority
on November 10, 2019 by guest
http://jvi.asm.org/
1
2
3
4
_w
*-4---DI
RF
_RF
a,-
R
I
A~~I
L.mp
---Di
RI B
,--
.--RI
B1
,
_
-
RIB2
FIG. 15. Slab-gelelectrophoresis ofEcoRI
digests
of
RF DNA.RFDNA labeled with 32Pwasdigested
for
3hat37°Cwith 100 UofEcoRI. Thedigestwasextracted with
phenol,
precipitated
with ethanolat-20'C,
andredissolvedin 20M Tris(pH
7.5)-i
mM EDTA.Aliquots
wereseparately
fractionated
by
alkalinedenatura-tion-BDCorneutral BDCchromatography.Aportion
of
32P-labeleddimer RF DNA(cf.
pool
B,
Fig.
3)wasalsodigestedwith EcoRI.Electrophoresiswasfor18hat
16°C
ataconstantvoltage of
50V.Samples
were:(1)EcoRIdigestofRFDNA; (2)1MTBS eluateofalkalinedenaturation-BDC
chromatography
of
(1);(3)1 M TBS eluate ofneutral BDCchromatography of (1); (4) EcoRIdigest of
dimer RFDNA. Viral DNA andfragments are labeledasfollows: dimer RFDNA,
DIRF;
RFDNA, RF; EcoRI-A, RIA;dimerEcoRI-B,
DIRIB; and
EcoRI-B,
and-B2,RIB,andRIB2,respectively.ofthe monomer RF DNA molecules do notdi- 15to25% of thetotalat severaltimes
p.i.
andmerize invitro. alsowith
pulse-chase labeling. Experiments
us-Dimer RF DNA was found to be
relatively
ingthedensity
label BUdRweredonetodeter-constantin
proportion
tomonomerRF DNAat minewhether dimer moleculesmayformby:
(i)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:16.501.42.441.44.508.2]b' a'
Eco RI B2 b 4- iC
b a a b b' a'
Eco RIB, _______________
a' b' b a
dimer Eco RI B a b b a'C a
b'
b asI .. . I . . . ..
100 0 100 990
Base Pairs Eco RI A
cleavage
siteFIG. 16. Modelfor the structure proposed for the left end ofH-i RF DNA. Letters representterminal sequences of 50 to 70 nucleotides, and sequence complementarity is indicated by a primed letter. The structuresshownareillustrated withagap(arrow) betweenadjacent terminiofthe V and Cstrands,whichin
some cases iscovalently closedasindicatedby a dot.
random annealing ofmonomers with cohesive AAV DNA(23). It wassuggestedthat covalent
left termini, perhaps in vitro; (ii) replication of linkage between plus and minus strands of
previously formed dimer DNA; or (iii) failure of AAV may be the result of the 3' end of one
replicature intermediates of monomer RF DNA strandacting as the primer for the initiation of
to segregate. The results showedthat dimer RF the C strand synthesis, as had also been
pro-DNA incorporated the density label more posed previously for MVM DNA (26). If this
slowly than monomer RF DNA (i.e., became were the case for H-1, then newly labeled RF
hybrid density)andruledoutnonsegregation of DNA would be expected to contain relatively
monomerRI moleculesastheimmediatemech- moreof the covalentlinkage thanthe total RF
anism of formation ofdimer DNA. The com- DNA pool. In the present study the contrary
plexity of densitylabelingwithlabeling periods resultwasobtained; RF DNAlabeledfor 3.5 h
greater than 30 min precluded a definitive dis- followedby a 1-h chase had a higher proportion
tinction between (i) possible formation of la- ofthe covalently linked V to C strands than
beled dimers by random association of mon- DNAlabeled for the 0.5 hprecedingthe extrac-omerRFDNA,or(ii) by replication of parental tion.
dimer DNA. The location of the covalent linkage was
Tattersall et al.reported whatappearedtobe shown to be in the EcoRI-B fragment, and it
intramolecular reannealing ofMVMRF DNA seems mostlikely that it is locatedat the left afterheatdenaturationat100°Cinasolution of endin an invertedself-complementary "turna-moderate ionic strength (27). In a subsequent round" suggested by the dimerizationobserved
report inthisseries, we will describe evidence occurring atthissite. Dimer RF DNA was
ana-foraregionattheright end of RF DNA witha lyzed in asimilar manner, and covalent link-veryhigh meltingtemperature, which, if pres- ageofVtoC strandswasfoundfor at least 19% ent in MVM DNA, could have accounted for ofthepopulation. This linkagewas again dem-incomplete denaturation at
1000C.
Infact, the onstrated to occur in the EcoRI-B fragmentand alkaline sucrosegradient analysis
of MVM RF probablyatthemolecular left end of RFDNA. DNA did not show the expected proportion of The presence of dimer length V-C single single-stranded DNA greater than monomer strands in some of the dimer RF DNAmole-length (26). Nevertheless, a morerigorous test cules strongly implies that atleast this type of
forcovalentlinkageof VtoCstrandsinH-1 RF dimerexistedinvivo, whatever themechanism
DNAwas carried out usingalkaline denatura- of their formation.
tionfollowed byrapidneutralization andBDC Finally, thedistributionof isotope in the two
chromatography to separate double-stranded EcoRI fragments, after short labeling times,
DNA that had reannealed rapidly from DNA wasexaminedtogaininsight as to the location
with single-stranded regions. After two cycles ofthe origin for replication of RF DNA and ofthis procedure, a fraction was isolated that progeny DNA synthesis. When isotope is first
represented19% of thetotal RF DNA, and this introduced into the precursorpools, it is
incor-fraction sedimented as a dimer length single- porated into RI molecules. After a labeling pe-stranded DNA in alkaline sucrose gradients. riod that does not greatly exceed the time
re-During the preparation of this manuscript a quired for a round of replication, RI molecules
similar structure wasreportedfor intracellular that have completed replicationduring the