Diminished Production of Monocyte Proinflammatory Cytokines during Human Immunodeficiency Virus Viremia Is Mediated by Type I Interferons
12
0
0
Full text
Figure
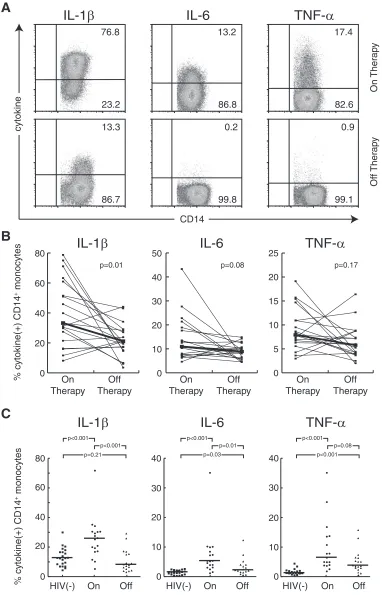
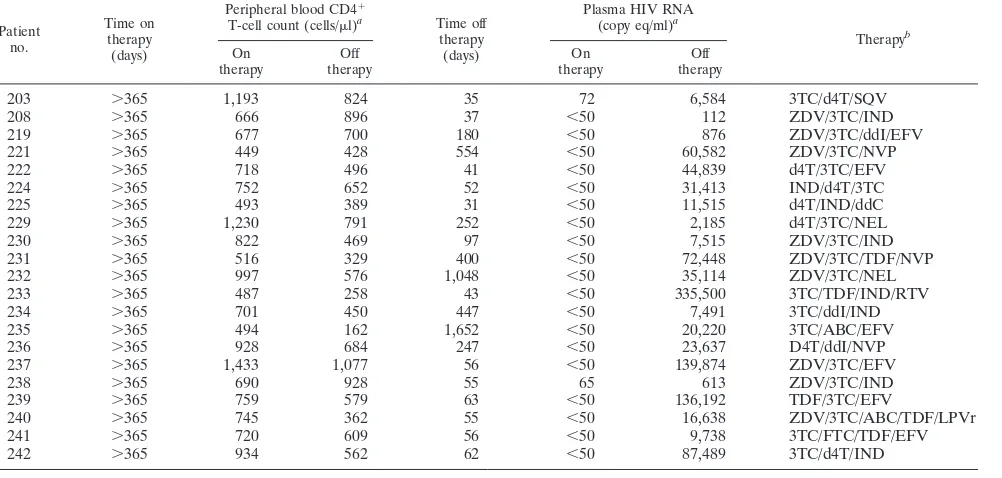

![FIG. 4. Diminished monocyte proinflammatory cytokine production [% cytokine(monocytes] from HIV-1-infected patients offwere not enhanced by the addition of autologous PBMC of patients on therapy when the populations were separated by a transwell insert](https://thumb-us.123doks.com/thumbv2/123dok_us/167532.44676/6.585.52.537.69.471/diminished-proinammatory-production-monocytes-autologous-populations-separated-transwell.webp)
+3
Related documents