JOURNAL OFVIROLOGY, Oct. 2005, p. 12218–12230 Vol. 79, No. 19 0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.19.12218–12230.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Four-Dimensional Visualization of the Simultaneous Activity of
Alternative Adeno-Associated Virus Replication Origins†
Daniel L. Glauser,
1Okay Saydam,
1N. Alexander Balsiger,
1‡ Irma Heid,
1R. Michael Linden,
2Mathias Ackermann,
1and Cornel Fraefel
1*
Institute of Virology, University of Zurich, 8057 Zurich, Switzerland,1and Department of Gene and Cell Medicine,
Mount Sinai School of Medicine, New York, New York 100292
Received 21 April 2005/Accepted 5 July 2005
The adeno-associated virus (AAV) inverted terminal repeats (ITRs) contain the AAV Rep protein-binding site (RBS) and the terminal resolution site (TRS), which together act as a minimal origin of DNA replication. The AAV p5 promoter also contains an RBS, which is involved in Rep-mediated regulation of promoter activity, as well as a functional TRS, and origin activity of these signals has in fact been demonstrated previously in the presence of adenovirus helper functions. Here, we show that in the presence of herpes simplex virus type 1 (HSV-1) and AAV Rep protein, p5 promoter-bearing plasmids are efficiently amplified to form large head-to-tail concatemers, which are readily packaged in HSV-1 virions if an HSV-1 DNA-packaging/cleavage signal is
provided incis. We also demonstrate simultaneous and independent replication from the two alternative AAV
replication origins, p5 and ITR, on the single-cell level using multicolor-fluorescence live imaging, a finding which raises the possibility that both origins may contribute to the AAV life cycle. Furthermore, we assess the differential affinities of Rep for the two different replication origins, p5 and ITR, both in vitro and in live cells and identify this as a potential mechanism to control the replicative and promoter activities of p5.
Adeno-associated virus (AAV) is a nonpathogenic human parvovirus with a single-stranded DNA genome of 4,680 nu-cleotides (nt). The AAV genome consists of two open reading frames (ORFs) including therepand thecapgenes, flanked by two 145-nucleotide inverted terminal repeats (ITRs). Therep
genes are transcribed from two different promoters, the p5 promoter and the p19 promoter. The former controls tran-scription of the large rep genes, rep68 and rep78, while the latter controls expression of the small rep genes, rep40 and
rep52. Thecapgene is transcribed from the p40 promoter and encodes VP1, VP2, and VP3, which form the icosahedral cap-sid (reviewed in reference 33). The ITRs can form T-shaped secondary structures, and they comprise the Rep-binding site (RBS) and the terminal resolution site (TRS), which together act as a minimal origin of DNA replication (ITR ori) (55, 56). AAV has a unique biphasic life cycle that is characterized by a productive infection in the presence of a helper virus, such as adenovirus (Ad) or herpes simplex virus (HSV), and a latent infection that occurs in the absence of helper virus (reviewed in reference 33). The AAV type 2 (AAV2) genome, for instance, can site-specifically integrate into human chromosome 19 at position 19q13.4, a locus that is termedAAVS1(8, 22, 23, 44). This process requires the RBS incis, as well as either of the large Rep proteins, Rep68 or -78 (1, 50, 61). The p5 promoter also contains an RBS (28), which is involved in the Rep-mediated regulation of the p5 activity (24, 38). However, the
discovery of a functional TRS within the p5 promoter (54) raised the possibility that it may also act as an alternative origin of viral DNA replication (Fig. 1). Indeed, several groups have reported the amplification of integratedrep-capsequences in the absence of ITRs and in the presence of adenovirus (3, 4, 10, 27, 51). Furthermore, the RBS and TRS within the p5 promoter have recently been demonstrated to act as an origin responsible for this ITR-independent replication (31, 35). In addition, and possibly related to its function as the origin of DNA replication, the p5 promoter sequence can also mediate, albeit inefficiently, the packaging of single-stranded, ITR-defi-cientrep-capsequences into AAV particles (34) and enhance Rep-mediated, site-specific integration of ITR-flanked trans-gene DNA (39). In fact, the p5 sequence incisis sufficient to allow site-specific integration of transgene DNA intoAAVS1in the absence of ITRs (40).
The activity of the putative replication origin within the p5 promoter (p5 ori) in the presence of ITR origins and its po-tential role in the AAV life cycle have not been determined. Other viruses, including herpes simplex virus type 1 (HSV-1) (30, 49), Epstein-Barr virus (reviewed in reference 53), and baculoviruses (21, 37), also contain alternative origins of DNA replication. In some viruses, the alternative replication origins have distinct activities in different phases of the viral life cycle (reviewed in reference 53) or else are differentially regulated (12), while in other cases, the biological functions of the alter-native origins are unknown.
In this study, we investigated the replicative capacity of the p5 ori in the presence of HSV-1 as the helper virus. In partic-ular, we used a live-cell visualization assay to address the ques-tion of whether both the ITR and the p5 origins can be active simultaneously and therefore potentially contribute to the AAV life cycle. The p5 RBS consists of one central GAGC tetranucleotide repeat and four flanking imperfect repeats,
* Corresponding author. Mailing address: Institute of Virology, Uni-versity of Zurich, Winterthurerstrasse 266a, CH-8057 Zurich, Switzer-land. Phone: 41 44 6358713. Fax: 41 44 6358911. E-mail: cornelf@vetvir .unizh.ch.
† Supplemental material for this article may be found at http://jvi .asm.org.
‡ Present address: Molecular Biomedicine, Swiss Federal Institute of Technology, 8952 Zurich-Schlieren, Switzerland.
12218
on November 8, 2019 by guest
http://jvi.asm.org/
whereas the ITR RBS comprises three central GAGC repeats and two flanking imperfect repeats (Fig. 1). As the GAGC sequence is necessary for binding of Rep (6, 7, 29, 58), it seems conceivable that the efficiency of Rep binding to the p5 RBS and ITR RBS may be different and that this differential affinity may control the formation of individual p5 and ITR replication compartments. To test this hypothesis, we monitored the dis-tribution of AAV Rep protein in the course of DNA replica-tion from the ITR or the p5 origin in live cells and compared its affinity for the two alternative replication origins in an in vitro DNA binding assay. An interesting side observation was that in the presence of HSV-1 and AAV Rep, p5 origin-bear-ing plasmids were efficiently amplified to form head-to-tail concatemers, which were readily packaged in HSV-1 virions if an HSV-1 DNA-packaging/cleavage signal was provided incis. This opens the possibility to design novel HSV/AAV hybrid vectors that combine the efficient replication/integration func-tions of the AAV p5 element with the large transgene capacity of HSV-1.
MATERIALS AND METHODS
Cells and viruses.VERO 2-2 cells (46) and HeLa cells were maintained in Dulbecco’s modified Eagle medium supplemented with 10% fetal bovine serum,
100 units/ml penicillin G, 100g/ml streptomycin, and 0.25g/ml amphotericin
B. For culture of VERO 2-2 cells, 500g/ml G418 was included in addition.
Wild-type HSV-1 strain F was grown, and titers were determined in VERO 2-2 cells.
Plasmids.Plasmids pRep, pRep-red (see Fig. S1 in the supplemental mate-rial), and pAAVlacO were described previously (9, 13). Plasmid pSV2-EYFP/ lacI expressing a fusion gene for enhanced yellow fluorescent protein (EYFP)
linked to thelacrepressor (LacI) was kindly provided by D. L. Spector (Cold
Spring Harbor Laboratory, Cold Spring Harbor, N.Y.) (52). Plasmids pSA1.TetO.EYFPnlsTetR and pSA1.TetO.ECFPnlsTetR (48) were kindly pro-vided by R. D. Everett (MRC Virology Unit, Glasgow, United Kingdom).
Bac-terial artificial chromosome fHSV⌬pac⌬27⌬Kn and plasmid pEBHICP27
to-gether represent a replication-competent, packaging-defective HSV-1 genome (42) and were used to provide HSV-1 helper functions for packaging of pP5RcagGFP into HSV-1 particles.
Construction of the other plasmids used in this study was done as follows. (i)
For pCMVrep-red (expressing the first 522 codons of rep68/rep78fused to
DsRed2from the human cytomegalovirus immediate-early 1 enhancer/promoter [CMV promoter] [see Fig. S1 in the supplemental material]), pDsRed2-1 (Clon-tech) was cleaved with AgeI and XmaI and self-ligated, resulting in
pDsRed2-1/N3, which contains a shifted open reading frame forDsRed. A 0.5-kb fragment
containing the CMV promoter was inserted between the SacI and HindIII sites of pDsRed2-1/N3, resulting in pCMVDsRed2-1/N3. A fragment containing the
first 522 codons of AAVrep68/rep78flanked by HindIII sites originating from
plasmid pBsR68/78 (C. Fraefel, unpublished material) was inserted into the unique HindIII site of pCMVDsRed2-1/N3, resulting in pCMVrep-red. (ii) For
pCMVrep-HcRed (expressing the first 522 codons ofrep68/rep78fused toHcRed
from the CMV promoter [see Fig. S1 in the supplemental material]), plasmid pHcRed1-1 (Clontech) was first cut with AgeI and XmaI and religated, resulting
in pHcRed1-1/N3, which contains a shifted ORF forHcRed. Subsequently, the
2.2-kb BglII-KpnI fragment from pCMVrep-red (containing the CMV promoter
and the first 522 codons ofrep68/rep78) was inserted between the BglII and KpnI
sites on pHcRed1-1/N3, resulting in pCMVrep-HcRed. (iii) For pCMVrep68/
78-kan (expressingrep68/rep78from the CMV promoter [see Fig. S1 in the
supplemental material]), first pCMVrep-red was cut with BglII and SalI, and the 1.7-kb fragment (containing the CMV promoter and the first 369 codons of
rep68/rep78) was inserted between the BglII and SalI sites of pRep (13), resulting in the ampicillin-resistant plasmid pCMVrep68/78. Second, pCMVrep68/78 was cut with BglII and NotI, and the 3.1-kb fragment (containing the CMV promoter, therep68/rep78genes, and a simian virus 40 polyadenylation signal) was inserted between the BglII and NotI sites on pHcRed1-1 (Clontech), resulting in the
kanamycin-resistant plasmid pCMVrep68/78-kan. Expression of rep68/rep78
from a kanamycin-resistant plasmid was required, because in the replication assay (see Fig. 3B), replication products of pBs-p5-tetO were detected with a probe for the ampicillin resistance gene. (iv) For pBs-p5-tetO, a 330-bp SalI fragment (containing seven tetracycline repressor [TetR] binding sites) from pSA1.TetO.EYFPnlsTetR (48) was self-ligated, and a five-copy repeat was
in-serted into the unique XhoI site of the pBluescript II KS(⫹) cloning vector
(Stratagene), resulting in plasmid pBstetO. A fragment containing the AAV p5 promoter (AAV2 nt 152 to 299) was amplified by PCR from plasmid pAV2 (25)
using primers p5ABforward (5⬘AAAATTACTAGTGGAGTCGTGACGTGAA
TTA3⬘) and p5Dgreverse (5⬘AAAATTGCGGCCGCCCGCTTCAAAATGGA
GA3⬘), which were designed to introduce a SpeI and a NotI site (underlined),
respectively. The resulting PCR product was cut with SpeI and NotI and ligated between the SpeI and NotI sites in pBstetO, resulting in plasmid pBs-p5-tetO. The correct orientation and sequence of the insert were confirmed by sequenc-ing. (v) For pEYFPTetR and pECFPTetR (expressing fusion genes for EYFP-TetR and enhanced cyan fluorescent protein [ECFP]-EYFP-TetR, including nuclear localization signals from the CMV promoter), pSA1.TetO.EYFPnlsTetR and pSA1.TetO.ECFPnlsTetR (48) were cut with NheI and HpaI, and the resulting 1.5-kb fragment (containing the fusion genes for EYFP-TetR and ECFP-TetR, including nuclear localization signals) were inserted between the NheI and HpaI sites of pEGFP-N3 (Clontech), resulting in the plasmids pEYFPTetR and
pECFPTetR. (vi) For pRep-CFP-pac (expressing the first 528 codons ofrep78
fused to the gene for ECFP from the p5 promoter and containing an HSV-1
packaging/cleavage signal [pac]), plasmid pRep-CFP (Fraefel, unpublished),
which expresses the first 528 codons ofrep78fused to the gene for ECFP from
the p5 promoter, was cut at the unique XhoI site, and a 1.4-kb XhoI fragment
[image:2.585.48.283.65.313.2]from pBXPpacPX (43) (containing HSV-1pac) was inserted, resulting in plasmid
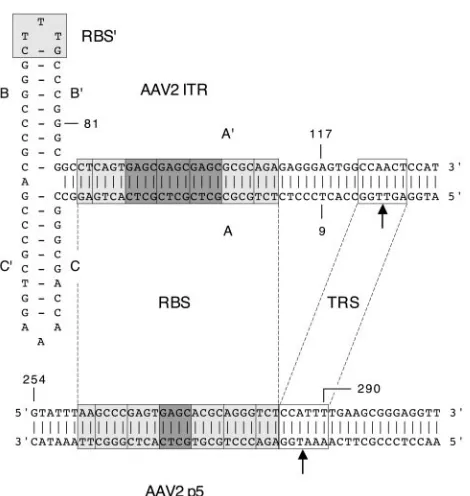
FIG. 1. Sequence alignment of the RBS and TRS of the AAV2 ITR and the AAV2 p5 promoter. The 25-bp RBS of the A-stem (28) and its homologue in the p5 promoter (28) are indicated by boxes in light gray. The A-stem RBS contains three perfect GAGC tetranucle-otide repeats (dark-gray boxes) flanked by two imperfect repeats, while the p5 RBS contains only one perfect GAGC repeat and four imper-fect repeats. The CTTTG element within the secondary-structure el-ement of the ITR, which was previously described as enhancing Rep binding affinity (41), is indicated by a light-gray box and is termed RBS⬘. The TRS of the A-stem (2, 17) and its homologue in the p5 promoter (54) are indicated by open boxes. Nicking at the A-stem TRS occurs between the two thymidine bases (top arrow), while at the p5 promoter TRS it occurs between a thymidine and an adenosine base (bottom arrow). Note that the p5 TRS is spaced 10 bp closer to the RBS than the A-stem TRS. The borders of the DNA substrates used in the EMS assays (Fig. 8 and 9) are indicated by the numbered AAV2 nucleotides: ITR substrate, nt 9 to 117 (single stranded and self-annealed); A-stem substrate, nt 81 to 117 (double stranded); p5 sub-strate, nt 254 to 290 (double stranded).
on November 8, 2019 by guest
http://jvi.asm.org/
pRep-CFP-pac. (vii) For pP5RcagGFP (based on pRep-CFP-pac and in addition containing an enhanced green fluorescent protein [EGFP] expression cassette), a 2.8-kb SalI fragment from plasmid pcagGFP (containing the gene for EGFP
under thecagpromoter and a bovine growth hormone polyadenylation signal,
bGHpA [Fraefel, unpublished]) was inserted into the SalI site upstream of the p5 promoter of pRep-CFP-pac, resulting in pP5RcagGFP.
Replication assays.Replication assays in VERO 2-2 cells were performed essentially as described previously (9, 13). For the results presented in Fig. 2A,
cells were cotransfected with 0.4g pRep-red or pCMVrep-red and 0.2g pRep
and superinfected with HSV-1 at a multiplicity of infection (MOI) of two 50%
tissue culture infective doses (TCID50) per cell or mock infected. For the results
presented in Fig. 2B, cells were transfected with 0.3g pRep and superinfected
with HSV-1 at an MOI of 1. For the results presented in Fig. 3B, cells were
cotransfected with 0.2g pBs-p5-tetO or pBs-tetO and 0.2g
pCMVrep68/78-kan or pCMVrep-HcRed and superinfected with HSV-1 at an MOI of 1 or mock
infected. A control transfection without anyrep-expressing plasmid was also
[image:3.585.97.493.71.499.2]performed. Hirt DNA (16) prepared 48 h postinfection (p.i.) was digested with DpnI and/or BglII as indicated in the figures. Agarose gel electrophoresis, trans-fer to nylon membranes, hybridization with digoxigenin (DIG)-labeled probes, and immunological detection have been described previously (9, 13). PCR am-plification and DIG labeling of probes was performed using the PCR DIG Probe Synthesis Kit (Roche) according to the manufacturer’s manual. For the results in
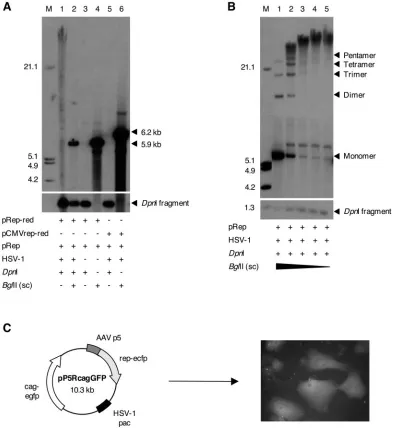
FIG. 2. Replication of AAV p5-containing plasmids. (A) Replication of plasmid pRep-red. VERO 2-2 cells were cotransfected with pRep (lanes 1 to 6) and either pRep-red (lanes 1 to 4) or pCMVrep-red (lanes 5 and 6). Subsequently, the cells were superinfected with HSV-1 (lanes 1, 2, 5, and 6). Hirt DNA prepared 48 h later was digested with DpnI and/or BglII and analyzed by Southern blotting with a DIG-labeledDsRed
probe. The sizes of the linearized plasmids pRep-red (5.9 kb) and pCMVrep-red (6.2 kb), as well as a prominent fragment of the DpnI digest, are indicated (arrows). (B) Conformation of p5 replication products. VERO 2-2 cells were transfected with plasmid pRep and superinfected with HSV-1. Hirt DNA was prepared as in panel A, digested with DpnI and decreasing amounts of BglII, and analyzed by Southern blotting with a DIG-labeledrepprobe. The bands corresponding to the linear monomer and multimers of pRep and a prominent fragment of the DpnI digest are indicated (arrows). The fragment sizes of the DIG-labeled molecular weight standard (M) are indicated on the left.⫹, present;⫺, absent. (C) Packaging of p5 replication products into HSV-1 particles. Schematic representation of pP5cagGFP (left) and representative fluorescence micrograph of transduced cells (right). The AAV p5 promoter (AAV p5), the HSV-1 DNA-packaging/cleavage signal (HSV-1 pac), therep-ecfp
fusion gene (rep-ecfp), and the EGFP expression cassette (cag-egfp) are shown.
12220 GLAUSER ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
Fig. 2A, replication products from plasmid pRep-red were detected with a
683-bp probe to the entireDsRed2coding sequence. For the results presented in
Fig. 2B, replication products of plasmid pRep were detected with a 1.6-kb probe
to therepgene (AAV2 nt 324 to 1904). For the results in Fig. 3B, replication
products of pBs-p5-tetO were detected with a 780-bp probe to the ampicillin resistance gene (ORF nt 4 to 783).
Packaging into HSV-1 virions. VERO 2-2 cells were cotransfected with pP5RcagGFP, packaging-defective HSV-1 helper DNA (42), and pRep, and putative vector particles were prepared and titrated as described previously (13, 42).
Live visualization of AAV DNA replication. (i) Transfection-infection proce-dure.The detailed transfection-infection procedure was described previously (9). Briefly, the day before transfection, 50,000 HeLa cells were plated on Lab-Tek four-well chamber slides or chambered coverglasses (Nalge Nunc International). The cells were transfected using Lipofectamine Plus Reagent as described by the manufacturer (Invitrogen). The amounts of individual plasmids used for
transfection were as follows: pBs-p5-tetO, pAAVlacO, and pBs-tetO, 25 ng; pEYFPTetR, pECFPTetR, pSV2-EYFP/lacI, pCMVrep68/78-kan, and pCM-Vrep-HcRed, 2.5 ng. Helper functions for AAV replication were provided by superinfection with HSV-1 at an MOI of 1 to 4 50% tissue culture infective doses at 3 to 4 h after transfection. Cultures of live or fixed cells were examined by standard or confocal fluorescence microscopy between 16 and 48 h after trans-fection.
(ii) Microscopy.Cells were observed by standard and confocal microscopy essentially as described previously (9). Images from confocal microscopy were deconvolved with a blind deconvolution algorithm using the Huygens Essential program suite (SVI, Hilversum, The Netherlands) and processed with Imaris 4.1.1 (Bitplane AG, Zurich, Switzerland) and Adobe Photoshop Elements 2.0 (Adobe) software.
[image:4.585.61.524.68.447.2]Electrophoretic mobility shift (EMS) assays.As DNA substrates for Rep binding, synthetic oligonucleotides corresponding to the ITR in the hairpin conformation (ITR substrate), the RBS of the ITR (A-stem substrate), or the
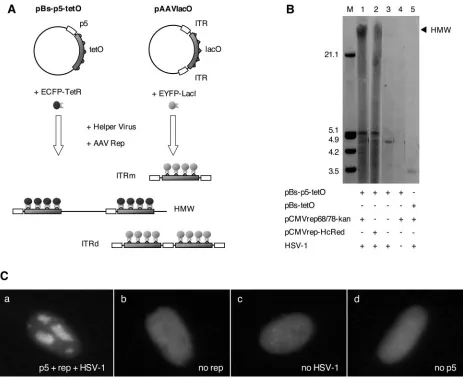
FIG. 3. Live visualization of AAV DNA replication from alternative replication origins. (A) Schematic representation of live-visualization assays. Replication from the AAV p5 ori is visualized with plasmid pBs-p5-tetO, which contains the AAV p5 promoter (p5) and five reiterations of the seven-copy tetO sequence comprising a total of 35 TetR binding sites. HSV-1 and Rep-dependent accumulation of concatameric replication products (HMW) is visualized by binding of an ECFP-TetR fusion protein. Visualization of replication from the ITR origins (pAAVlacO), which employs lacO interactions with an EYFP-LacI fusion, has been described previously (9). Double-stranded monomeric (ITRm) and dimeric (ITRd) replication intermediates are indicated. (B) Replication assay with plasmid pBs-p5-tetO. VERO 2-2 cells were cotransfected with pBs-p5-tetO and pCMVrep68/78-kan (lanes 1 and 4) or pCMVrep-HcRed (lane 2); pBs-p5-tetO (lane 3); or pBs-tetO and pCMVrep68/78-kan (lane 5) and superinfected with HSV-1 (lanes 1, 2, 3, and 5). Hirt DNA was prepared at 48 h p.i., digested with DpnI, and analyzed by Southern blotting with a probe for the ampicillin resistance (Ampr) gene present on pBs-p5-tetO. Concatameric, high-molecular-weight replication products are indicated (HMW).⫹, present;⫺, absent. (C) Formation of nuclear AAV p5 replication compartments in the presence of the p5 ori, AAVrep, and HSV-1. Cells were cotransfected with pBs-p5-tetO, pEYFPTetR, and pCMVrep68/78-kan (a and c), pBs-p5-tetO and pEYFPTetR (b), or pBs-tetO, pEYFPTetR, and pCMVrep68/78-kan (d) and superinfected with HSV-1 (a, b, and d) or mock infected (c). The micrographs were taken 36 h p.i. under a standard fluorescence microscope with a filter specific for EYFP.
on November 8, 2019 by guest
http://jvi.asm.org/
RBS of the p5 promoter (p5 substrate) were used. The oligonucleotides were synthesized at Microsynth GmBH (Balgach, Switzerland) and contained the following AAV2 sequence elements: ITR substrate, nt 9 to 117 (single stranded); A-stem substrate, nt 81 to 117 (double stranded); p5 substrate, nt 254 to 290
(double stranded) (Fig. 1). 5⬘32P labeling of the ITR substrate and of the bottom
strands of the A-stem and p5 substrates was performed by Hartmann Analytic GmbH (Braunschweig, Germany). Self-annealing of the ITR substrate or an-nealing of the two complementary strands of the A-stem and the p5 substrates was performed as described previously (29). His-tagged Rep68 protein (Rep68H) was expressed and purified as described elsewhere (45, 59, 60). Bind-ing reactions were performed as described previously (18). The amounts of Rep protein and DNA substrates are indicated for each experiment. After addition of
2l loading buffer (consisting of binding buffer including 50% glycerol and
0.025% bromophenol blue), the binding reactions were resolved on 4%
poly-acrylamide gels (29:1 poly-acrylamide/bisacryamide weight ratio) in 0.25⫻
Tris-bo-rate-EDTA at 200 V for 2.25 h (see Fig. 9) or 3 h (see Fig. 8). The gels were dried and analyzed with the PhosphorImager system (Molecular Dynamics) and Im-ageQuant software (Molecular Dynamics).
RESULTS
The AAV p5 promoter contains an HSV-1- and
Rep-depen-dent origin of DNA replication.We used plasmid pRep-red
(9), which contains a rep-DsRed2 fusion gene under the p5 promoter, in order to assess the replicative capability of the p5 ori in HSV-1-infected cells. pRep-red served as both the rep-licon plasmid and the expression plasmid for the Rep-DsRed fusion protein, which was previously shown to support replica-tion of recombinant AAV (rAAV) genomes, although at a lower efficiency than the wild-type Rep proteins (9). In order to provide maximal Rep activity for efficient replication of pRep-red, a plasmid expressing all four wild-typerepgenes (pRep) was included in the assay. Maps of allrep-expressing plasmids used in this study are provided in Fig. S1 in the supplemental material. Transfection of pRep-red and pRep and superinfec-tion with HSV-1 led to the accumulasuperinfec-tion of DpnI-resistant, high-molecular-weight (HMW) replication products of the pRep-red plasmid DNA, as demonstrated by Southern blot analysis using a probe specific for DsRed (Fig. 2A, lane 1). Cleavage with restriction endonuclease BglII, which cuts only once within pRep-red, reduced the HMW replication products to a single discrete band with a size corresponding to the linearized pRep-red (Fig. 2A, lane 2). In the absence of HSV-1 or the p5 promoter (plasmid pCMVrep-red), no replication products were observed (Fig. 2A, lanes 3 and 5). In order to assess the requirement for the Rep protein, we performed replication assays with plasmid pBs-p5-tetO, which contains the p5 promoter but no rep sequences. Replication of this plasmid was observed only in the presence of HSV-1 and when
repwas provided intrans(Fig. 3B).
In order to analyze the conformation of the HMW p5 rep-lication products, we performed partial restriction digests of replication products from either plasmid pRep-red (data not shown) or plasmid pRep, which contains the wild-typerepgene under control of the native p5 promoter (Fig. 2B) and served as both the replicon and therep-expressing plasmid. Replica-tion products from plasmid pRep were detected with a probe for therepcoding sequence. Partial digestion with BglII led to the disappearance of the HMW smear and appearance of a ladder of discrete bands, with sizes corresponding to linear monomers and multimers of plasmid pRep. These data suggest a head-to-tail-linked concatameric conformation of the repli-cation products, consistent with a rolling-circle mechanism of
DNA replication. As the conformations and sizes of the p5 replicons strongly resembled those of replicating HSV-1 ge-nomes and amplicons, we hypothesized that they could be packaged into HSV-1 particles if HSV-1 DNA-packaging/ cleavage signals (pac) were present incis. We therefore con-structed plasmid pP5RcagGFP, which contains the p5 pro-moter, an EGFP reporter gene, and an HSV-1pacsignal (Fig. 2C). Replication products of pP5RcagGFP were readily pack-aged into HSV-1 virions in the presence of HSV-1 helper functions, as transduction of VERO 2-2 cells resulted in EGFP expression (Fig. 2C).
Live-cell visualization of DNA replication from the AAV p5 ori.We recently established a live-cell visualization assay for rAAV DNA replication employinglacoperator (lacO)-lac re-pressor (LacI) interactions combined with autofluorescent pro-teins (9). The rAAV genome in plasmid pAAVlacO consisted of 40 lacO repeats that are flanked by ITRs acting as origins of DNA replication (ITR replicon). In order to similarly visualize the replication of plasmids containing the p5 ori (p5 replicon), we developed an assay that is based on the interaction of tetracycline operator sequence (tetO) with the tetracycline re-pressor DNA binding domain (TetR) fused to autofluorescent proteins (Fig. 3A). TetO-TetR interactions have previously been employed for the visualization of parental HSV-1 ge-nomes and HSV-1 replication compartments in live cells (48). We constructed the pBs-p5-tetO replicon, which contains the p5 promoter and five repeats of the seven-copy tetO array (a total of 35 TetR binding sites) (Fig. 3A). Replication of pBs-p5-tetO in the presence of HSV-1 and AAV Rep should lead to the accumulation of sufficient tetO binding sites to support visualization of the replicated DNA by an EYFP-TetR fusion protein.
We first verified the efficient replication of plasmid pBs-p5-tetO using replication assays in VERO 2-2 cells, which yielded abundant DpnI-resistant replication products in the presence of HSV-1 and either the wild-type Rep proteins or a Rep-HcRed fusion protein (Fig. 3B). When cells were cotransfected with plasmids pBs-p5-tetO and pEYFPTetR, as well as the
rep-expressing plasmid pCMVrep68/78-kan, and then superin-fected with HSV-1, we observed the formation of nuclear rep-lication compartments as early as 16 to 24 h p.i. (Fig. 4). The time of appearance of the replication compartments was de-pendent on the MOI of the helper virus (data not shown). Depending on the transfection efficiency, 1 to 5% of the cells displayed 16⫾6 (mean⫾standard deviation;n⫽40) cation compartments. In the course of ongoing plasmid repli-cation, the replication compartments grew larger until they filled most of the nucleus (Fig. 4). At early stages, the yellow fluorescence was diffuse in the nucleus, while at later stages, it was entirely recruited into the replication compartments, indi-cating that EYFP-TetR synthesis was not limiting for the vi-sualization of the replicating DNA. Nuclear fluorescence stayed diffuse in the absence of eitherrep, HSV-1, or p5 se-quences as tested with the control plasmid pBs-tetO, which contains the tetO repeats but no p5 promoter sequences (Fig. 3C).
Covisualization of AAV p5 replication and AAV Rep protein.
We have previously shown in live cells that AAV Rep is effi-ciently recruited into rAAV replication compartments (9). Be-cause Rep is also required for replication from the p5 ori (Fig.
12222 GLAUSER ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
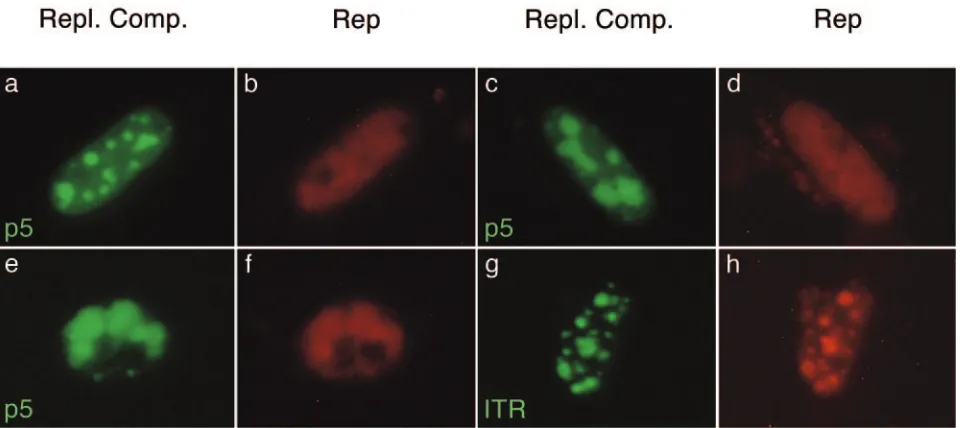
3B and C), we expected a similar recruitment also into p5 replication compartments. In order to test this hypothesis, we constructed pCMVrep-HcRed, which constitutively expresses the first 522 codons ofrep68/rep78fused to HcRed under the control of the CMV promoter. Replication assays demon-strated that the Rep-HcRed fusion protein was functional, as it was able to mediate replication of plasmid pBs-p5-tetO, al-though the yield of replication product was somewhat lower than with wild-type Rep68/78 protein (Fig. 3B, compare lanes 1 and 2). Next, pCMVrep-HcRed was tested for its ability to support replication of both ITR and p5 replicons in live-cell visualization assays. The formation of nuclear replication
[image:6.585.53.535.69.252.2]com-partments was readily observed with both replicons, but most interestingly, the distribution of the Rep-HcRed protein was strikingly different between ITR and p5 replication compart-ments. While Rep-HcRed was almost completely recruited into even very small rAAVlacO (ITR) replication compart-ments (Fig. 5, g and h), the red fluorescent Rep protein re-mained diffuse within the nucleus in both early and interme-diate stages of pBs-p5-tetO replication (Fig. 5, a to d). Only at a very late stage, when replication compartments filled most of the nucleus, did Rep-HcRed fusion protein colocalize with p5 replication sites (Fig. 5, e and f). To rule out the possibility of altered behavior of Rep due to the fusion to HcRed, the same
FIG. 4. Formation of AAV p5 replication compartments in live HeLa cells. Cells were cotransfected with pBs-p5-tetO, pEYFPTetR, and pCMVrep-HcRed and superinfected with HSV-1. Time lapse images starting at 27.5 h p.i. were taken with a confocal microscope with settings specific for EYFP (p5 replication compartments) and HcRed (AAV Rep; inset).
FIG. 5. Visualization of AAV p5 and ITR replication compartments (green) and AAV Rep (red) in live HeLa cells. Cells were cotransfected with pBs-p5-tetO, pEYFPTetR, and pCMVrep-HcRed (a to f) or pAAVlacO, pSV2-EYFP/lacI, and pCMVrep-HcRed (g and h). After transfection, the cells were superinfected with HSV-1. The images were taken between 24 and 36 h p.i. with a standard fluorescence microscope and filters specific for EYFP (AAV replication compartments) or Texas Red (AAV Rep). Three different nuclei containing p5 replication compartments and one nucleus containing ITR replication compartments are shown. Repl. Comp., replication compartments.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.54.534.463.677.2]12224
on November 8, 2019 by guest
http://jvi.asm.org/
experiments were repeated with wild-type Rep68/78 and stain-ing with a monoclonal, Rep-specific antibody. As expected, Rep-HcRed fusion protein behaved identically to wild-type Rep68/78 protein (data not shown).
Covisualization of AAV replication from the p5 and ITR
origins. As a model to assess whether the p5 and the ITR
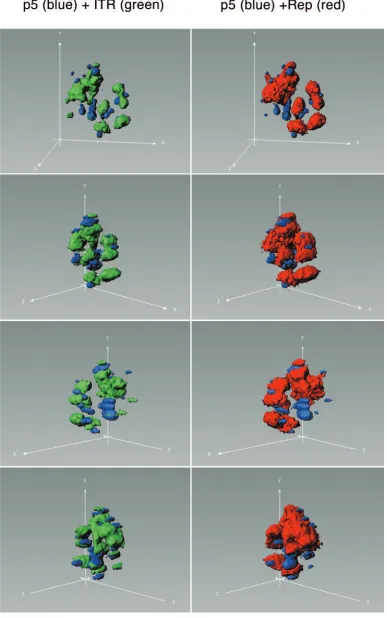
origins can, in principle, be simultaneously active during the AAV life cycle, we combined the visualization system for DNA replication from the p5 ori with our previously established visualization system for replication from the ITR origins (9). The combination of the two visualization systems allows the simultaneous monitoring of DNA replication from two differ-ent viral origins in a single live cell (Fig. 3A). In order to separately visualize pAAVlacO and pBs-p5-tetO replication, we used an ECFP-TetR fusion protein to label p5 replication products and EYFP-LacI to label ITR replication products. Upon cotransfection of pAAVlacO, pBs-p5-tetO, pSV2-EYFP/lacI, pECFPTetR, and a rep-expressing plasmid, fol-lowed by infection with HSV-1, the simultaneous replication of pAAVlacO and pBs-p5-tetO could be observed. Interestingly, the p5 and ITR replication compartments seemed to form independently of each other, but preferentially in neighboring nuclear compartments. In most cases, p5 and ITR replication compartments were juxtaposed to each other, but rarely was a p5 compartment found within an ITR compartment or vice versa. However, the different replication compartments did not fuse but rather stayed separate from each other (Fig. 6 and 7). Moreover, Rep-HcRed fusion protein strongly accumulated in ITR replication compartments, while the p5 replication com-partments appeared to be deprived of Rep. Possibly, both the requirement for HSV-1 helper functions and the local avail-ability of Rep protein accumulated at the ITR compartments direct the p5 replication compartments to adjacent locations. Together with the data presented in Fig. 5, these findings suggest a lower affinity of the AAV Rep protein for the p5 ori than for the ITR origins, the recruitment of fewer Rep mole-cules into Rep-p5 than into Rep-ITR complexes, or a combi-nation of both. This question was addressed in the following in vitro DNA binding experiments.
Differential affinities of Rep for p5 and ITR origins.In order
to confirm and further compare the interaction of Rep with the p5 and ITR origins, we proceeded to test the binding of Rep to these sequence elements by EMS assays. We assessed the bind-ing of prokaryotically expressed His-tagged Rep68 protein (Rep68H) to (i) the hairpinned ITR, (ii) the linear A-stem of the ITR (absence of the B and C palindromes excludes the formation of DNA secondary structures), and (iii) the p5 ori (Fig. 1). We first tested different concentrations of Rep protein in order to determine the required Rep concentration for each substrate. As demonstrated in Fig. 8A and B, the amounts of Rep required to achieve 90% binding of 0.04 pmol substrate DNA were 60 ng and 120 ng for the ITR and the A-stem
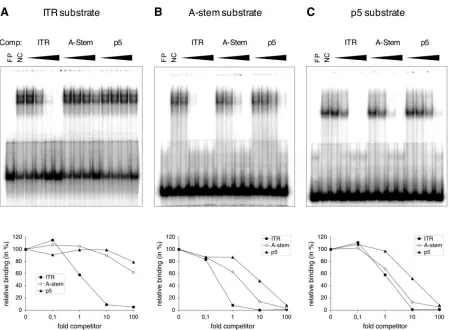
substrate, respectively, confirming the previously demon-strated higher affinity of Rep for the hairpinned ITR substrate over the A-stem substrate (29, 41). However, at the highest Rep amounts used (240 ng), only approximately 45% of the p5 substrate was bound. Moreover, the Rep-p5 complexes mi-grated significantly faster than Rep-ITR or Rep–A-stem com-plexes and appeared to consist of a single, discrete band as opposed to the ladder of bands observed with the ITR sub-strate and, under some conditions, with the A-stem subsub-strate. In order to estimate the relative affinities of Rep for the dif-ferent substrates, we performed reciprocal competition assays using unlabeled competitor DNA. The amount of bound probe in the absence of competitor DNA was set as 100% relative binding. In a first experiment (Fig. 9A), binding of 15 ng of Rep to 0.1 pmol of radiolabeled ITR substrate in the presence of increasing (0.1-, 1-, 10-, and 100-fold) amounts of unlabeled ITR, A-stem, and p5 competitor DNA was determined. While addition of a 10-fold excess of homologous competitor DNA reduced the binding by approximately 90%, the presence of a 100-fold excess of A-stem or p5 competitor led to reductions by only 40% and 20%, respectively. In analogous experiments, Rep binding to radiolabeled A-stem or p5 substrates in the presence of the different competitor DNA substrates was as-sessed (Fig. 9B and C). Both experiments confirmed the results from Fig. 9A, suggesting the following order of relative affin-ities for the three substrates: ITR⬎A-stem⬎p5. An estima-tion of the efficiency of competiestima-tion by each substrate sug-gested a 10-fold difference between ITR and A-stem, a 10-fold difference between A-stem and p5, and a 100-fold difference between ITR and p5 (Fig. 9A to C).
The data from the EMS assays led us to the following con-clusions. (i) The assembled Rep complex on the p5 ori is significantly smaller than on the hairpinned ITR or on the linear A-stem. Consequently, the p5 RBS sequesters fewer Rep molecules than the RBS on the A-stem of the ITR. (ii) Rep binds about 100-fold more efficiently to the hairpinned ITR than to the p5 RBS. (iii) Rep has an approximately 10-fold-lower affinity for the linear p5 than for the linear A-stem substrate. Therefore, the difference between p5 and the hair-pinned ITR is likely a consequence of the absence of secondary structure and the different sequence of the RBS. Taken to-gether, these data demonstrate that the differential recruit-ment of Rep protein into alternative AAV replication com-partments (Fig. 4 to 7) is due to both a differential affinity of Rep for the alternative origins and the formation of distinct Rep-DNA complexes.
DISCUSSION
In the present study, we assessed the replicative capability of the AAV p5 ori in the presence of HSV-1 helper functions and analyzed the conformation of the resulting replication
prod-FIG. 6. Covisualization of AAV DNA replication from alternative replication origins and AAV Rep protein. Three individual nuclei are shown. HeLa cells were cotransfected with pBs-p5-tetO, pAAVlacO, pECFPTetR, pSV2-EYFP/lacI, and pCMVrep-HcRed and superinfected with HSV-1. Images of live (a to f) or fixed (g to s) cells were taken between 36 and 48 h p.i. with a confocal microscope with settings specific for ECFP (p5 replication compartments), EYFP (ITR replication compartments), and HcRed (Rep protein). The images represent projections through three-dimensional reconstructions of nuclei.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 7. Three-dimensional view of the nucleus presented in Fig. 6 (a to f). p5 replication compartments are stained blue, ITR replication compartments green, and Rep protein red. Deconvolved three-dimensional reconstructions of the nucleus were processed in Imaris software using the surpass view mode.
12226 GLAUSER ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
ucts. We found that plasmids containing the p5 ori formed head-to-tail-linked, concatameric replication products with sizes of at least 150 kb in the presence of HSV-1 and AAV Rep (Fig. 2). Although the functionality of the RBS and TRS within the AAV p5 promoter as an origin of DNA replication has been demonstrated in the presence of both Ad (31, 35) and HSV-1 helper functions (this study), the question arose as to whether the p5 ori has a function in the replication of wild-type AAV. As a first step toward answering this question, we ex-amined whether both the p5 and the ITR origins can be active simultaneously in the same cell. For this, we established a live-cell assay for visualizing replication from the AAV p5 ori and combined it with the previously described visualization system for AAV ITR replication (9) (Fig. 3A). We show si-multaneous and efficient DNA replication from both replica-tion origins in the same cell, demonstrating that replicareplica-tion from the p5 ori in the presence of ITR replication intransis, in principle, possible. We therefore hypothesize that the AAV p5 ori, besides acting as an enhancer of ITR-mediated site-specific integration (39), may also play a role in rescue and replication of wild-type AAV. However, it remains to be de-termined whether both origins are simultaneously active if they are present incis, a situation that is very difficult to assess but that would more closely mimic wild-type AAV replication. The simultaneous visualization of AAV ITR- and p5-mediated rep-lication also led to some interesting findings about the nuclear sites of AAV DNA replication. First, p5 and ITR replication compartments seemed to form independently of each other but preferentially in neighboring nuclear compartments (Fig. 6
[image:10.585.66.516.75.338.2]and 7). The helper virus and cellular factors that define these selected sites of DNA replication still remain to be identified, but it is likely that the HSV-1 helicase-primase complex and the HSV-1 major DNA binding protein ICP8 (56, 57) play pivotal roles. Second, the distribution of the AAV Rep protein was strikingly different between sites of p5- and ITR-mediated replication. While Rep protein was efficiently recruited into the ITR replication foci, the p5 compartments appeared to be deprived of Rep (Fig. 6 and 7), indicating a differential affinity of AAV Rep for these two replication origins. In order to address whether Rep has a lower affinity for the p5 ori than for the ITR origins or whether fewer Rep molecules are recruited into Rep-p5 than into Rep-ITR complexes, we compared the binding of Rep to these alternative replication origins in EMS assays (Fig. 8 and 9). The results suggested the formation of significantly smaller complexes of Rep on the p5 ori than on the ITR ori (Fig. 8). To date, it has become apparent that Rep acts as a multimer. The recently solved crystal structures of the Rep motor domain have shown the presence of an “arginine finger” that highlights the requirement for an oligomeric in-terface in order for an active ATPase pocket to form (19, 20). Furthermore, it has been proposed that Rep can form different oligomers depending on substrate characteristics. Although our results do not allow conclusions about the exact number of Rep molecules assembled on a Rep-p5 complex, it is possible that Rep binds as a dimer to the p5 ori, in agreement with Zhou et al., who have proposed that Rep dimers constitute the active forms for both the ATPase and nicking activities (62). In contrast, it has been suggested that Rep binds to the ITRs as
FIG. 8. Rep binding to AAV replication origins: titration of Rep protein. (A) Twofold dilutions of Rep protein were bound to 0.04 pmol of radiolabeled DNA substrates consisting of the hairpinned ITR (ITR), the linear A-stem (A-stem), or the linear p5 substrate (p5). (B) Quantifi-cation of the fraction of bound DNA substrate. The PhosphorImager data were analyzed by ImageQuant software, and the bound fraction (yaxis) was plotted against the amount of Rep protein (xaxis).
on November 8, 2019 by guest
http://jvi.asm.org/
a hexamer (14, 15, 47). Although our EMS assays were not designed to analyze the conformation of Rep complexes formed on the different substrate DNAs, they confirmed the migration of Rep-ITR and, under some conditions, Rep–A-stem complexes in a ladder of several bands, consistent with the results of several previous studies (26, 29, 47). In contrast, Rep-p5 complexes migrated faster than Rep-ITR and Rep–A-stem complexes and appeared to migrate in a single discrete band. The finding that Rep–A-stem complexes migrated in several bands in Fig. 9B but in a single complex in Fig. 8A is probably due to the different substrate concentrations used (0.04 pmol in Fig. 8A and 0.1 pmol in Fig. 9B) or the different resolutions of the gel (see Materials and Methods). For in-stance, in a previous study that used identical binding condi-tions, Rep bound to hairpinned AAV termini was also de-tected in a single band rather than in a ladder of bands (18). Reciprocal competition experiments revealed an approxi-mately 100-fold-lower affinity of Rep for the p5 ori than for the hairpinned ITR ori (Fig. 9). These experiments also confirmed previous studies according to which the affinity of Rep for the linear A-stem sequence, without the B and C palindromes, is markedly lower than for the ITR in its hairpinned conforma-tion (29, 41). However, in our experimental setting, the
[image:11.585.67.517.69.399.2]differ-ence was only approximately 10-fold compared to the at least 125-fold difference found by McCarty et al. (29) or the 170-fold difference described by Ryan et al. (41). Since bases outside the A-stem RBS also have an effect on the efficiency of Rep binding (7, 41, 58), it is likely that the different A-stem sub-strates used account for the observed differences in Rep-bind-ing affinity. In contrast, Chiorini and coworkers found approx-imately equal affinities of Rep protein for a linear A-stem substrate and a hairpinned ITR substrate (7). Our results from the EMS assays led us to the conclusion that the lower affinity of Rep for the p5 RBS than for the ITR RBS is due to both the absence of the DNA secondary structure comprising the en-hancer of Rep binding, RBS⬘, and the sequence differences between the p5 RBS and the ITR RBS (Fig. 1). On the basis of the finding that the GAGC sequence is necessary for binding of Rep (6, 7, 29, 58), it seems conceivable that Rep binding to the p5 RBS containing only one perfect GAGC repeat is less efficient than its binding to the RBS within the ITRs containing three perfect GAGC repeats (Fig. 1). Moreover, the presence of only one perfect GAGC repeat may also explain why the p5 RBS assembles fewer Rep molecules than its counterpart in the A-stem. Nevertheless, DNA replication from the AAV p5 ori seemed to proceed with efficiency equal to that from the
FIG. 9. Rep binding to AAV replication origins: reciprocal competition analysis. Fifteen nanograms (A and B) or 60 ng (C) of Rep protein was bound to 0.1 pmol of radiolabeled ITR (A), A-stem (B), or p5 DNA (C) substrate in the presence of a 0.1-, 1-, 10-, or 100-fold amount of unlabeled ITR, A-stem, or p5 competitor DNA. The diagrams at the bottom represent the relative binding (yaxis) as a function of the amount (n-fold) of competitor DNA (fold competitor;xaxis). The amount of bound probe was quantified by ImageQuant analysis of the PhosphoImager data, and the value in the absence of competitor DNA was set as 100%. Comp., Competitor; FP, free probe; NC, no competitor.
12228 GLAUSER ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
ITR origins, possibly supported by the fact that the p5 replicon plasmid was provided as a circular DNA molecule, leading to the efficient mechanism of rolling-circle DNA replication. However, this situation is not entirely artificial, as circular duplex monomer AAV genomes have previously been de-scribed during AAV rescue and replication (31, 32). It also needs to be considered that in our experimental settings the expression levels ofrepfrom the constitutive CMV promoter were constantly high and there apparently was sufficient Rep protein present to support replication from both origins. With regard to the biological significance of the differential affinity of Rep to the p5 RBS and the ITR RBS, we hypothesize that at initial stages of AAV rescue or replication, the low levels of Rep protein are entirely recruited by high-affinity binding to the ITRs and initiate DNA replication, which in turn increases the gene dose ofrep-capand consequently raises the expres-sion levels of rep in concert with transactivation of the p5 promoter by Ad E1A (5) or HSV-1 ICP0 (11). The presence of abundant quantities of Rep protein then leads to low-affinity binding to the p5 promoter, which in turn mediates transcrip-tional repression ofrep68/rep78(38) and, simultaneously, may initiate DNA replication from the p5 ori, possibly accounting for the generation of defective interfering AAV particles.
Taken together, our data demonstrate the functionality of the RBS and TRS contained within the p5 promoter as a Rep-and HSV-1-dependent origin of DNA replication. The obser-vation that the resulting concatameric replication products were packageable into HSV-1 particles opens the way for the development of a new generation of HSV/AAV hybrid vectors (reviewed in reference 36) that use the replication/integration functions from AAV and the large transgene capacity of HSV-1. We also demonstrated that AAV DNA replication from alternative replication origins, in particular the ITR and the p5 origins, is in principle possible in the same cell and elucidated the differential affinity of AAV Rep protein for the RBS within the ITRs and the p5 promoter. The confirmation of the data obtained from the live-cell visualization assay by the biochemical assays underscores the accuracy and usefulness of the live-visualization model to study not only alternative but also competing viral replication origins. Future experiments will possibly resolve the so far elusive role of the p5 ori in rescue, replication, and site-specific integration of wild-type AAV.
ACKNOWLEDGMENTS
We thank Oleg Georgiev and Walter Schaffner (Institute of Molec-ular Biology, University of Zurich, Zurich, Switzerland) for their help with the EMS assays and Urs Ziegler (Institute of Anatomy, University of Zurich, Zurich, Switzerland) for his help with confocal time lapse microscopy.
This work was supported by the Swiss National Science Foundation grant 3100A0100195 (C.F.) and National Institutes of Health R01GM62234 (R.M.L.).
REFERENCES
1.Balague, C., M. Kalla, and W. W. Zhang. 1997. Adeno-associated virus Rep78 protein and terminal repeats enhance integration of DNA sequences
into the cellular genome. J. Virol.71:3299–3306.
2.Brister, J. R., and N. Muzyczka.1999. Rep-mediated nicking of the adeno-associated virus origin requires two biochemical activities, DNA helicase
activity and transesterification. J. Virol.73:9325–9336.
3.Caleo, M., M. C. Cenni, M. Costa, E. Menna, L. Zentilin, S. Giadrossi, M. Giacca, and L. Maffei.2002. Expression of BCL-2 via adeno-associated virus
vectors rescues thalamic neurons after visual cortex lesion in the adult rat.
Eur. J. Neurosci.15:1271–1277.
4.Chadeuf, G., D. Favre, J. Tessier, N. Provost, P. Nony, J. Kleinschmidt, P. Moullier, and A. Salvetti.2000. Efficient recombinant adeno-associated virus production by a stable rep-cap HeLa cell line correlates with adenovirus-induced amplification of the integrated rep-cap genome. J. Gene Med.
2:260–268.
5.Chang, L. S., Y. Shi, and T. Shenk.1989. Adeno-associated virus P5 pro-moter contains an adenovirus E1A-inducible element and a binding site for
the major late transcription factor. J. Virol.63:3479–3488.
6.Chiorini, J. A., M. D. Weitzman, R. A. Owens, E. Urcelay, B. Safer, and R. M. Kotin.1994. Biologically active Rep proteins of adeno-associated virus type
2 produced as fusion proteins inEscherichia coli. J. Virol.68:797–804.
7.Chiorini, J. A., S. M. Wiener, R. A. Owens, S. R. Kyostio, R. M. Kotin, and B. Safer.1994. Sequence requirements for stable binding and function of Rep68 on the adeno-associated virus type 2 inverted terminal repeats. J.
Vi-rol.68:7448–7457.
8.Dutheil, N., F. Shi, T. Dupressoir, and R. M. Linden.2000. Adeno-associ-ated virus site-specifically integrates into a muscle-specific DNA region.
Proc. Natl. Acad. Sci. USA97:4862–4866.
9.Fraefel, C., A. G. Bittermann, H. Bueler, I. Heid, T. Bachi, and M. Acker-mann.2004. Spatial and temporal organization of adeno-associated virus
DNA replication in live cells. J. Virol.78:389–398.
10.Gao, G. P., F. Lu, J. C. Sanmiguel, P. T. Tran, Z. Abbas, K. S. Lynd, J. Marsh, N. B. Spinner, and J. M. Wilson.2002. Rep/Cap gene amplification and high-yield production of AAV in an A549 cell line expressing Rep/Cap.
Mol. Ther.5:644–649.
11.Geoffroy, M. C., A. L. Epstein, E. Toublanc, P. Moullier, and A. Salvetti.
2004. Herpes simplex virus type 1 ICP0 protein mediates activation of
adeno-associated virus type 2repgene expression from a latent integrated form.
J. Virol.78:10977–10986.
12.Hardwicke, M. A., and P. A. Schaffer.1997. Differential effects of nerve
growth factor and dexamethasone on herpes simplex virus type 1oriL- and
oriS-dependent DNA replication in PC12 cells. J. Virol.71:3580–3587.
13.Heister, T., I. Heid, M. Ackermann, and C. Fraefel.2002. Herpes simplex virus type 1/adeno-associated virus hybrid vectors mediate site-specific inte-gration at the adeno-associated virus preinteinte-gration site, AAVS1, on human
chromosome 19. J. Virol.76:7163–7173.
14.Hickman, A. B., D. R. Ronning, R. M. Kotin, and F. Dyda.2002. Structural unity among viral origin binding proteins: crystal structure of the nuclease
domain of adeno-associated virus. Rep. Mol. Cell10:327–337.
15.Hickman, A. B., D. R. Ronning, Z. N. Perez, R. M. Kotin, and F. Dyda.2004. The nuclease domain of adeno-associated virus rep coordinates replication
initiation using two distinct DNA recognition interfaces. Mol. Cell13:403–
414.
16.Hirt, B.1969. Replicating molecules of polyoma virus DNA. J. Mol. Biol.
40:141–144.
17.Im, D. S., and N. Muzyczka.1990. The AAV origin binding protein Rep68 is an ATP-dependent site-specific endonuclease with DNA helicase activity.
Cell61:447–457.
18.Im, D. S., and N. Muzyczka.1989. Factors that bind to adeno-associated
virus terminal repeats. J. Virol.63:3095–3104.
19.James, J. A., A. K. Aggarwal, R. M. Linden, and C. R. Escalante.2004. Structure of adeno-associated virus type 2 Rep40-ADP complex: insight into nucleotide recognition and catalysis by superfamily 3 helicases. Proc. Natl.
Acad. Sci. USA101:12455–12460.
20.James, J. A., C. R. Escalante, M. Yoon-Robarts, T. A. Edwards, R. M. Linden, and A. K. Aggarwal.2003. Crystal structure of the SF3 helicase from
adeno-associated virus type 2. Structure11:1025–1035.
21.Kool, M., J. T. Voeten, R. W. Goldbach, J. Tramper, and J. M. Vlak.1993.
Identification of seven putative origins ofAutographa californicamultiple
nucleocapsid nuclear polyhedrosis virus DNA replication. J. Gen. Virol.
74:2661–2668.
22.Kotin, R. M., R. M. Linden, and K. I. Berns.1992. Characterization of a preferred site on human chromosome 19q for integration of
adeno-associ-ated virus DNA by non-homologous recombination. EMBO J.11:5071–5078.
23.Kotin, R. M., M. Siniscalco, R. J. Samulski, X. D. Zhu, L. Hunter, C. A. Laughlin, S. McLaughlin, N. Muzyczka, M. Rocchi, and K. I. Berns.1990. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci.
USA87:2211–2215.
24.Kyostio, S. R., R. S. Wonderling, and R. A. Owens.1995. Negative regulation of the adeno-associated virus (AAV) P5 promoter involves both the P5 rep binding site and the consensus ATP-binding motif of the AAV Rep68
pro-tein. J. Virol.69:6787–6796.
25.Laughlin, C. A., J. D. Tratschin, H. Coon, and B. J. Carter.1983. Cloning of
infectious adeno-associated virus genomes in bacterial plasmids. Gene23:
65–73.
26.Li, Z., J. R. Brister, D. S. Im, and N. Muzyczka.2003. Characterization of the adeno-associated virus Rep protein complex formed on the viral origin of
DNA replication. Virology313:364–376.
27.Liu, X., F. Voulgaropoulou, R. Chen, P. R. Johnson, and K. R. Clark.2000.
on November 8, 2019 by guest
http://jvi.asm.org/
Selective Rep-Cap gene amplification as a mechanism for high-titer
recom-binant AAV production from stable cell lines. Mol. Ther.2:394–403.
28.McCarty, D. M., D. J. Pereira, I. Zolotukhin, X. Zhou, J. H. Ryan, and N. Muzyczka.1994. Identification of linear DNA sequences that specifically
bind the adeno-associated virus Rep protein. J. Virol.68:4988–4997.
29.McCarty, D. M., J. H. Ryan, S. Zolotukhin, X. Zhou, and N. Muzyczka.1994. Interaction of the adeno-associated virus Rep protein with a sequence within
the A palindrome of the viral terminal repeat. J. Virol.68:4998–5006.
30.McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M. C. Frame, D. McNab, L. J. Perry, J. E. Scott, and P. Taylor.1988. The complete DNA sequence of the long unique region in the genome of herpes simplex virus
type 1. J. Gen. Virol.69:1531–1574.
31.Musatov, S., J. Roberts, D. Pfaff, and M. Kaplitt.2002. Acis-acting element that directs circular adeno-associated virus replication and packaging. J.
Vi-rol.76:12792–12802.
32.Musatov, S. A., T. A. Scully, L. Dudus, and K. J. Fisher.2000. Induction of circular episomes during rescue and replication of adeno-associated virus in
experimental models of virus latency. Virology275:411–432.
33.Muzyczka, N., and K. I. Berns.2001. Parvoviridae: the viruses and their
replication, p. 2327–2346.InD. M. Knipe and P. M. Howley (ed.), Fields
virology, 4th ed., vol. 2. Lippincott Williams & Wilkins, Philadelphia, Pa. 34.Nony, P., G. Chadeuf, J. Tessier, P. Moullier, and A. Salvetti.2003. Evidence
for packaging ofrep-capsequences into adeno-associated virus (AAV) type
2 capsids in the absence of inverted terminal repeats: a model for generation
ofrep-positive AAV particles. J. Virol.77:776–781.
35.Nony, P., J. Tessier, G. Chadeuf, P. Ward, A. Giraud, M. Dugast, R. M. Linden, P. Moullier, and A. Salvetti.2001. Novelcis-acting replication ele-ment in the adeno-associated virus type 2 genome is involved in amplification
of integratedrep-capsequences. J. Virol.75:9991–9994.
36.Oehmig, A., C. Fraefel, X. O. Breakefield, and M. Ackermann.2004. Herpes simplex virus type 1 amplicons and their hybrid virus partners, EBV, AAV,
and retrovirus. Curr. Gene Ther.4:385–408.
37.Pearson, M., R. Bjornson, G. Pearson, and G. Rohrmann.1992. The Autog-rapha californicabaculovirus genome: evidence for multiple replication
ori-gins. Science257:1382–1384.
38.Pereira, D. J., D. M. McCarty, and N. Muzyczka.1997. The adeno-associated virus (AAV) Rep protein acts as both a repressor and an activator to
regulate AAV transcription during a productive infection. J. Virol.71:1079–
1088.
39.Philpott, N. J., C. Giraud-Wali, C. Dupuis, J. Gomos, H. Hamilton, K. I. Berns, and E. Falck-Pedersen.2002. Efficient integration of recombinant
adeno-associated virus DNA vectors requires a p5-repsequence incis. J.
Vi-rol.76:5411–5421.
40.Philpott, N. J., J. Gomos, K. I. Berns, and E. Falck-Pedersen.2002. A p5 integration efficiency element mediates Rep-dependent integration into
AAVS1 at chromosome 19. Proc. Natl. Acad. Sci. USA99:12381–12385.
41.Ryan, J. H., S. Zolotukhin, and N. Muzyczka.1996. Sequence requirements for binding of Rep68 to the adeno-associated virus terminal repeats. J. Virol.
70:1542–1553.
42.Saeki, Y., C. Fraefel, T. Ichikawa, X. O. Breakefield, and E. A. Chiocca.2001. Improved helper virus-free packaging system for HSV amplicon vectors using an ICP27-deleted, oversized HSV-1 DNA in a bacterial artificial
chro-mosome. Mol. Ther.3:591–601.
43.Saeki, Y., T. Ichikawa, A. Saeki, E. A. Chiocca, K. Tobler, M. Ackermann, X. O. Breakefield, and C. Fraefel.1998. Herpes simplex virus type 1 DNA
amplified as bacterial artificial chromosome inEscherichia coli: rescue of
replication-competent virus progeny and packaging of amplicon vectors.
Hum. Gene Ther.9:2787–2794.
44.Samulski, R. J., X. Zhu, X. Xiao, J. D. Brook, D. E. Housman, N. Epstein,
and L. A. Hunter.1991. Targeted integration of adeno-associated virus
(AAV) into human chromosome 19. EMBO J.10:3941–3950. (Erratum,
11:1228, 1992.)
45.Smith, D. H., P. Ward, and R. M. Linden.1999. Comparative characteriza-tion of Rep proteins from the helper-dependent adeno-associated virus type
2 and the autonomous goose parvovirus. J. Virol.73:2930–2937.
46.Smith, I. L., M. A. Hardwicke, and R. M. Sandri-Goldin.1992. Evidence that the herpes simplex virus immediate early protein ICP27 acts
post-transcrip-tionally during infection to regulate gene expression. Virology186:74–86.
47.Smith, R. H., A. J. Spano, and R. M. Kotin.1997. The Rep78 gene product of adeno-associated virus (AAV) self-associates to form a hexameric
com-plex in the presence of AAV ori sequences. J. Virol.71:4461–4471.
48.Sourvinos, G., and R. D. Everett.2002. Visualization of parental HSV-1 genomes and replication compartments in association with ND10 in live
infected cells. EMBO J.21:4989–4997.
49.Stow, N. D.1982. Localization of an origin of DNA replication within the TRS/IRS repeated region of the herpes simplex virus type 1 genome. EMBO
J.1:863–867.
50.Surosky, R. T., M. Urabe, S. G. Godwin, S. A. McQuiston, G. J. Kurtzman, K. Ozawa, and G. Natsoulis.1997. Adeno-associated virus Rep proteins target DNA sequences to a unique locus in the human genome. J. Virol.
71:7951–7959.
51.Tessier, J., G. Chadeuf, P. Nony, H. Avet-Loiseau, P. Moullier, and A. Salvetti.2001. Characterization of adenovirus-induced inverted terminal
re-peat-independent amplification of integrated adeno-associated virusrep-cap
sequences. J. Virol.75:375–383.
52.Tsukamoto, T., N. Hashiguchi, S. M. Janicki, T. Tumbar, A. S. Belmont, and D. L. Spector.2000. Visualization of gene activity in living cells. Nat. Cell
Biol.2:871–878.
53.Tsurumi, T., M. Fujita, and A. Kudoh.2005. Latent and lytic Epstein-Barr
virus replication strategies. Rev. Med. Virol.15:3–15.
54.Wang, X. S., and A. Srivastava.1997. A novel terminal resolution-like site in
the adeno-associated virus type 2 genome. J. Virol.71:1140–1146.
55.Ward, P., and K. I. Berns.1995. Minimum origin requirements for linear
duplex AAV DNA replication in vitro. Virology209:692–695.
56.Ward, P., M. Falkenberg, P. Elias, M. Weitzman, and R. M. Linden.2001. Rep-dependent initiation of adeno-associated virus type 2 DNA replication by a herpes simplex virus type 1 replication complex in a reconstituted
system. J. Virol.75:10250–10258.
57.Weindler, F. W., and R. Heilbronn.1991. A subset of herpes simplex virus replication genes provides helper functions for productive adeno-associated
virus replication. J. Virol.65:2476–2483.
58.Weitzman, M. D., S. R. Kyostio, R. M. Kotin, and R. A. Owens.1994. Adeno-associated virus (AAV) Rep proteins mediate complex formation between AAV DNA and its integration site in human DNA. Proc. Natl.
Acad. Sci. USA91:5808–5812.
59.Yoon, M., D. H. Smith, P. Ward, F. J. Medrano, A. K. Aggarwal, and R. M. Linden.2001. Amino-terminal domain exchange redirects origspecific
in-teractions of adeno-associated virus rep78 in vitro. J. Virol.75:3230–3239.
60.Yoon-Robarts, M., and R. M. Linden.2003. Identification of active site residues of the adeno-associated virus type 2 Rep endonuclease. J. Biol.
Chem.278:4912–4918.
61.Young, S. M., Jr., and R. J. Samulski.2001. Adeno-associated virus (AAV) site-specific recombination does not require a Rep-dependent origin of
rep-lication within the AAV terminal repeat. Proc. Natl. Acad. Sci. USA98:
13525–13530.
62.Zhou, X., I. Zolotukhin, D. S. Im, and N. Muzyczka.1999. Biochemical characterization of adeno-associated virus rep68 DNA helicase and ATPase
activities. J. Virol.73:1580–1590.
12230 GLAUSER ET AL. J. VIROL.