0022-538X/88/041194-09$02.00/0
Copyright C1988, AmericanSociety forMicrobiology
Physical
Characterization of
the
Herpes
Simplex
Virus
Latency-Associated Transcript
in Neurons
EDWARD K.
WAGNER,'*
GAYATHRIDEVI-RAO,' LAWRENCE T. FELDMAN,2 ANTHONY T. DOBSON,2 YI-FANZHANG,1 W. MICHAEL FLANAGAN,1 AND JACK G. STEVENS2Department ofMolecularBiology andBiochemistry, University of California, Irvine, Irvine, California 92717,1and
Department
ofMicrobiology
andImmunology,
University of California,
LosAngeles,
LosAngeles,
California900272
Received 15 October1987/Accepted 15 December 1987
RNAtransfer(Northern) blotanalysiswasused to performthe physicalcharacterizationof thetranscript expressed in murinesensory nerve ganglia latently infected with herpes simplex virus type 1. Most of this latency-associated transcript (LAT)wasisolated in thepoly(A)-fractionfromganglia.Asmaller RNAspecies
wasalsodetectedatless than 10% the abundance ofthemajorone. LATwasnot detectedwithprobesfrom DNAoutside thelimits of the larger species. In situ hybridization data correlated well with Northern blot analysis; however, low levels ofhybridizationwereseenwithprobes immediately outsidetheregionofviral DNA giving positive Northernblotsignals.Sl nuclease andprimerextensionmappingwereused to locatethe 5' end of the LAT 510 bases to the left of a Kpnl site at 0.783 map units. The 3' end of the major latency-associated specieswasmapped tojust withina310-base-pairSmaI fragmentlocated 660 to 970 base pairstotherightof theSallsite at 0.790mapunits. These datawerecorrelated withananalysisof thesequence
of the DNAencodingthistranscriptand itspossible function in the latentphaseof infection.
Latency is a hallmark of herpesvirus infection. In this
state, viralgenomes areavailable for reactivationof produc-tive infection, but the viralproducts diagnosticofan active infection aredifficult ifnotimpossibleto detect (9, 27). For herpes simplex virus type 1 (HSV-1),themolecular investi-gation of viralgenes expressed during the latent state has been quite difficult since only a minority ofcells in neural tissue harborlatent virus.
Despite such technical difficulties, it has been clear for
sometime that duringthe latentphase, antigens character-istic ofproductive infection arenotreproducibly detectable (29). Further, the viral genome appears to be present in circular orconcatenated forms rather than the linear
mole-culesfoundin the intactvirion (21).These dataindicatethat the latentphaseisoneinwhich viralgeneexpressionisquite limited,asituationanalogoustothatseenin latentinfections with the related Epstein-Barr herpesvirus (11).
These indications have recently been confirmed by our findinganabundant and previously unreported transcript in latently infected mouse sensory nerve ganglia (29). This latency-associated transcript (LAT) was estimated to be about thesame sizeas,and partially complementaryto,the
a transcript encoding ICP0, oneof the transcriptional
acti-vators expressed atthe earliest phases ofproductive infec-tion by HSV-1.
Since our initial communication, two othergroups have detected and partially characterized HSV RNA in latently infectedcells,andthis RNAwasfoundtobe expressedfrom the same general region of the genome we are studying (2, 22, 24, 26). Both generally confirm ourearlier results and those reported here. It should also be noted that Puga and Notkins(20) usedaprobe made by oligo(dT)-primedcDNA synthesis oflatently infected neuronalRNA and reported a weak signal hybridizing to the same general genomic
posi-tionasobservedforLAT. Thismayreflectapoly(A)+RNA species ofvery low abundance that we detected with diffi-culty inthepresent study.
* Corresponding author.
Wenowpresenttheprecisephysical characterizationand genomic location of the HSV-1 LAT expressed in latently infectedmurinesensorynervegangliaandcorrelate this with thesequenceof the viral DNAencodingit. Interestingly,the mostabundant speciesof thistranscript (>95%)was recov-ered in the poly(A)- fraction; like many other HSV tran-scripts, it lackslarge introns. Ourfindings suggestthatthis geneencodes aprotein; however, the dataobtainedtodate also suggest that a protein is not expressed by the major LATduringthe latentphase ofinfection.
MATERIALS ANDMETHODS
Establishment oflatentinfection in mice andprocessingof spinal ganglia. Six-week-old outbred male Swiss-Webster mice (Simonson Laboratories, Gilroy, Calif.) were used in allexperiments. Theywereinoculated in eachrearfoot with 108 PFU of HSV-1 strain KOS(M). The derivation ofthis virus strain and methods used for inoculation have been described previously (29, 32). Three or more weeks later, after latentinfections had been established in lumbosacral spinal ganglia (28), gangliaweredissected from the animals, quickfrozen inliquid nitrogen, and eithercuton acryostat
or storedat -70°Cuntil used.
RNA isolation andfractionation. RNA was isolated from latently infectedganglia by the guanidinium isothiocyanate-hot phenol method as described previously (29). Poly(A)-containingRNAwasselectedby oligo(dT)-cellulose (Collab-orative Research, Inc., Waltham, Mass.) chromatography (4). RNAwassize fractionatedbyelectrophoresis with 1.4%
agarosegels containing 10mMmethylmercuryhydroxideas previously described (4, 29); RNAwas transferred by elec-trophoresis onto nylon membranes (Gene Screen; New England Nuclear Corp., Boston, Mass.). In some
experi-ments,RNAwasfractionatedon1.2%agarosegels contain-ing6%formaldehyde and blottedonto membranes(13).
Primer extension. Primer extension experiments were done with a synthetic 20-nucleotide oligomer (5'-dTGG TGTGCTGTAACACGAGC-3'). The location of the oligo-mer within the sequence of the DNA encoding the LAT
1194
on November 10, 2019 by guest
http://jvi.asm.org/
transcript is shown in the Results. Samples of 15 ng of
5'-32P-labeled
primer were annealed with RNA from 15 to 20latently infected mouse ganglia. Extension wasdone at50°C
with 15 to 20 U ofreversetranscriptase (Life Sciences, Inc., St. Petersburg, Fla.) in 15,ul of abuffer (pH 8.3) containing 50 mM KCI, 35 mM Tris, 5 mM MgCl2, and 400 p.M each deoxynucleoside triphosphate as described in references 10
and 15. Sequences obtained by the dideoxy chain
termina-tion method (see below) from the same primer annealed to
DNA were cofractionated with the primer-extended cDNA
as an absolute size marker.
RecombinantDNA andsequenceanalysis. All recombinant
DNA was from the KOS(M) strain of HSV-1; fragments
were cloned into either pUC or M13 vectors as described
previously (13, 16). Restriction endonucleases were pur-chased from either Bethesda Research Laboratories, Inc. (Gaithersburg, Md.) orBoehringer MannheimBiochemicals (Indianapolis, Ind.). For synthesis of 32P-labeled DNA probes, the HSV-specific DNA fragment was isolated by digestion of the total plasmid with appropriate restriction
enzymes, followed by electrophoresis and electroelution. Synthesisofuniformlylabeled probes was accomplished by nick translation with
[a-32P]dCTP
(3Ci/4Lmol;
Amersham Corp., Arlington Heights, Ill.). For 5' end labeling of DNA for S1 nuclease analysis, the KpnI site at 0.783 map units (m.u.) was cut with Asp 718 to provide a 5'overhang. The5' phosphateresidue was then removed with bacterial alkaline phosphatase (Bethesda Research Laboratories) and32p
la-beled with
[_y-32P]ATP
(7 Ci/,umol; ICN Pharmaceuticals,Inc., Irvine, Calif.) and bacteriophage T4 kinase (Bethesda
Research Laboratories) (4, 13). The appropriate fragment was then released bydigestion withPstI.
Sequence analysis was done with convenient restriction sites within the DNA region of interest by the general
methods ofMaxam and Gilbert (14). Sequences determined
in thisway wereconfirmed by the dideoxy chain termination
method of Sanger et al. (24) with single-stranded DNA
cloned inM13.
Hybridization conditions. RNA transfer blots were
hybrid-ized with 40 x 106 cpm(Cerenkov) ofradiolabeled DNA in
the presence of50% formamide, 0.4 M Na+, 0.1 M HEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid) (pH8.0),0.005 M EDTA, and Denhardt solution (3) at 49°C for 40
h. Details and procedures for rinsing have been described
previously (4, 29). For
S1
nuclease mapping withdouble-stranded DNA under R-looping conditions, hybridization
was in the presence of80% formamide in the same buffer
without Denhardt solution for 12 h at 68°C. Before the
hybridization, the 5'-end-labeled DNA was denatured with
alkali and neutralized before addition of RNA. Conditions for
S1
nuclease digestion and fractionation of material onacrylamide gels containing 8 M urea have been described
previously (1, 6).
In situ hybridization methods. Appropriate cloned DNA
fragmentswerenick translated with
[3H]deoxynucleotides
tospecific
activities of2 x107
to3 x107
cpm/,ug of DNA (29).Each probe was used on cryostat-cut sections of
lumbosa-cral spinal ganglia taken from uninfected, latentlyinfected,
or acutely infected mice. In the latter instance, mice had beeninfected4dayspreviously in rearfootpads, and at the
time ofsacrifice, virus wasreplicating to highlevels in the
ganglia (32). Methods for preparation of slidesand execution
ofRNA-specific
hybridization
reactionswereamodification of those described by Haase et al. (8). In the presentexperiments,
probes were prehybridizedand the stringencyof the hybridization method was increased asdescribed by
Stroop et al. (31). After a 2-week exposure, the slides were developed, stained with Giemsa, and observed.
RESULTS
Mapping LAT mRNA. Our previous report established the direction of transcription of the HSV LAT transcript as opposite that ofICPO (29). In this study, we used a number
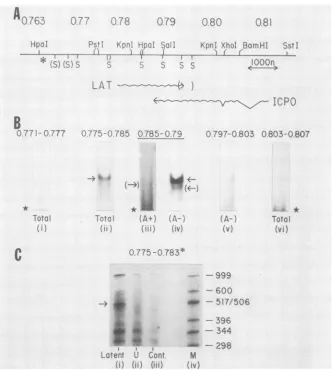
of subclones of DNA in the region of the HSV genome around the ICPO gene and RNA transfer (Northern) blots of RNA from latently infected ganglia to establish its extent. Our data are summarized in Fig. 1A, and representative blots are shown in Fig. 1B.
The transcript from latently infected cells migrated mar-ginally faster than did
ICPO
mRNA fractionated in parallelon methyl mercury gels. This difference in migration rates was accentuated when formaldehyde gels were used (data
notshown). Therefore, we concluded that the upper limit in
size was onthe order of 2.3 kilobases. Using total RNA, a
readily detectable transcript of this size was seen only with probes made with DNA spanning the 2,500 bases between
theSmaI sites at 0.777 and 0.794 m.u. (Fig.
1B,
lanes ii andiv).
Todetermine precisely the maximum extent of the 3' end
of LAT complementary to ICPO, we used small DNA
probes, and some ofthese data are shown in Fig. 2. Northern
blots hybridizedwith the660-base-pair SalI-SmaI fragment between 0.790 and 0.794 m.u. gave a strong positive signal.
Incontrast, the375-base-pairSmaI fragment immediately to
the right of this (0.794 to 0.796 m.u.) gave a weak positive signal for HSV-1 LAT. This signal is clearly seen in long exposures (Fig. 2). DNA probes made from fragments
im-mediately to the right of0.7% m.u. were negative (Fig. 1B,
lane v). Such data located the 3' end of the transcript to be
660 to 970 bases to the right of theSalIsiteat 0.790 m.u. The
low intensity of the signal obtainedwith this nick-translated SmaIfragment suggested that the transcript extends only a
short way (20 to 50bases) 3' of theSmaI site nearest theSalI
site at 0.790 m.u.
Asecond RNAband migrating with a size corresponding
to 1.8 to 2 kilobases was seen on most gels with probes
which hybridized to the major LAT species. Examples are
seen in Fig. 1B(lanes ii and iv) and in Fig. 2. This transcript
was found to be encoded by the same DNA strand as that encoding the major LAT by use of strand-specific probes
(data not shown). It was not detected with any probes
beyond the limits of the LAT, and we suggest that it represents a processed form of the major species (see
Discussion).
Although RNA of the size of the major andminor HSV-1
LATspecies was only detected withprobes mapping within
0.777 and 0.793 m.u., DNA probes encompassed by the SmaI fragment spanning 0.770 to 0.777 m.u. and probes containing DNA tothe right of the SmaI site at 0.796 m.u.
did hybridize to RNA migrating diffusely with asize of no
morethan 300 to 350 bases on some Northern blots (Fig.1B,
lanes i, v, and vi). This result implies that sometranscription
does occur beyond the nominal 5' and 3' ends ofthe LAT transcript in latently infected cells. Based on exposure
density of theautoradiographs, theamount of RNA
respon-sible for such diffuse signals was less than5% that of the 2.3-kilobase LAT transcript.
Insituhybridization oflatently infected ganglia. Indepen-dent data supporting the Northern blot mapping were ob-tained by in situ methods with the same probes as those used above. Examples of the in situ data with both latently
on November 10, 2019 by guest
http://jvi.asm.org/
077l
0.78
079
0
80_i.i .1
's 3)cC. v
5-
rrp*n.
Hpal 5 I1 I
KpnI Xhol BarnHI SstI II f. K'
:S S S 1OOn
"I,
-LAT
)1
ICePO
0 >75-0 785 0785-0.79
To-3
fA)
fA*
tr i,
(0A+)
(783
O) 775-0.78 3
:4
Latent Cont
(i) (ii) (iiI)
0797-0.803 0.803-0.807
(A-) (v)
-999
600
- 517/506
- 396
. - 344
298
M
(IV)
FIG. 1. Precise location of the LAT transcript onthe prototypical arrangementof the HSV-1 strain KOS(M) genome. (A) Summary restriction map of the region encoding the transcript. Unique restriction sites are indicated. SmaI sites (S) shown in parentheses are
approximatelocations, andnoSmaIsitestotherightof theKpnI siteat0.802m.u. areincluded. The asterisk(*)indicatesthe locationof
thebeginningof the longinternal repeat regionof thegenome. (B)Northern blotanalysisof RNA isolated from latentlyinfected mouse
sensorynerveganglia.Detailsof the methods usedaredescribedin the Materials and Methods and Results. RNA from25gangliawereused
for each Northern blot shown. Thearrowsindicate the sizeexpectedfor the LATtranscript;arrowsinparenthesesindicate minorspecies; thestarsindicate thepresenceof smalldiffusely migratingRNA discussed in the Results.TotalganglionRNAwasusedexceptfor lanes iii
andiv,in whichpoly(A)+andpoly(A)-fractions of thesameganglionRNApreparationwereused. Probes usedwere:0.771-0.777, SmaIto
SmaI; 0.775-0.785, PstI to HpaI; 0.785-0.79, HpaI to Sall; 0.797-0.803, SmaI to XhoI; 0.803-0.807, XhoI to BamHI. Exposure of
autoradiographswasfor 16hat-70°Cwith Cronexintensifyingscreens,exceptfor laneiii,whichwasexposedfor 6daysat thistemperature. (C) S1nucleasemappingof the5' end of LAT fromlatentlyinfectedganglionRNA(lane i).A DNAprobeextendingfrom thePstIsite to the
KpnI site(thelatter cleaved withAsp 718) selectively5'end labeledattheKpnIsitewashybridizedunderR-loopingconditionsasdescribed
in the text. S1 nuclease-resistant material wasfractionated on adenaturing 8% acrylamide-urea gel versus size standards derived from
EcoRI-Hinfl-digested pBR322DNA(lane M).The sizesareshowntotherightof laneM(nucleotides).Thespecificband of materialmigrating withasize ofapproximately490 bases is indicatedbythearrow.Other bandswerealsoseeninacontrol tracks of materialhybridizedwith uninfected-cell RNA(lane ii)and with commercial Escherichia colitRNA(lane iii).
infected anduninfected gangliaare shownin Fig.3. Strong, infected-cell-specific nuclear hybridization was observed with probes made from DNA fragments contained in the PstI-HpaI (0.775to0.785m.u.;Fig. 3A), HpaI-SaII (0.785to 0.790m.u.;datanotshown),and SalI-BamHI(0.790to0.807
m.u.; Fig. 3B) clones. No hybridization was seen in the uninfected controls (Fig. 3E and F). Positive signals were also obtained withprobesmadefromtheHpaI-PstI (0.764to 0.774 m.u.; Fig. 3C) and the BamHI-SstI (0.807 to 0.815
m.u.; Fig. 3D) DNA fragments. However, the signal from thesewaslessintense,and with the latterprobe, fewer than 50% of the nucleiwere labeled thanweredetected with the HpaI-SalI or the PstI-HpaI probes. With the HpaI-PstI
probes,therewasalsoahighbackgroundin uninfected cells (Fig. 3G).Thisbackground,and thepositive nuclearsignals
seen, mayrelatetothecross-hybridization reportedbetween this fragmentandribosomal orother cellulargenes(17, 19, 25). Any specific hybridizationdetected with thisprobeand thepositivenuclei observed with the BamHI-SstIprobemay
representthe small RNA detectable insomeNorthern blots. Alternatively, itmay reflecta low level oftranscription of othergenesthatarebelow the level of detectionbyNorthern blotanalysis (see Discussion).
Distribution of HSV-1 LAT betweenpoly(A)+and
poly(A)-fractions. Most of the LATtranscript was recovered inthe fractionlacking poly(A)uponoligo(dT)-cellulose
chromatog-A0.76
jHpaI
0.81
B
0. 7 71- 0.777
* Totsl
'i)
C
Total
(vi)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.143.475.75.445.2]0.790- 0.794-0.794 0.796
( z)(
16 Hr Exposure 120 Hr Exposure
FIG. 2. Northern blot analysis ofthe 3' end of the HSV-1 LAT. RNA samples from latently infectedmouse ganglia were fraction-ated asdescribed in the legend toFig. 1Cand electrophoretically transferredtofiltermembranes. The blots shown in the figure were hybridizedwith 28 x 106of32P-labelednick-translated probe made from either the Sall-SmaI fragment (0.790 to 0.794 m.u.) or the SmaI-SmaI fragments (0.794 to 0.796 m.u.) directly to the right of thisasshownin Fig. 1A. Exposuretimes shown were 16 and 120 h withintensifyingscreens as described in the legend to Fig. 1. Both the major and minor LAT transcripts are detectable with difficulty withthe probe covering 0.794 to 0.796 m.u.
raphy ofRNA preparations from whole ganglia (Fig. 1B).
When the poly(A)+ fraction of total ganglion RNA was
fractionated in parallel with the poly(A)- fraction and
hy-bridized with the same probe, a faint band of RNA with a
migration rate similar to that of the minor poly(A)- LAT
species could be detectedwith long exposures. An example of such anexperimentis shown in lanes iii and ivofFig. 1B. Here, the exposure time for the poly(A)+ fraction (lane iii)
A
B
was approximately 10 times longer than for the poly(A)-fraction (lane iv). We also found some diffusely migrating RNAofsmall size in the poly(A)+ fraction.
Preciselocationofcontiguous5'endof the LAT transcript. Welocated the contiguous 5' end of the LAT transcript using Si nuclease analysis of hybrids between latently infected neuronal RNA and DNA selectively 5' end labeled at the KpnI (Asp 718) site at 0.783 m.u. The results ofone such experimentareshown in Fig. 1C. The probewasisolatedas a 1,073-base-pair PstI-KpnI fragment (0.775 to 0.783 m.u.) cloned inpUC18, and hybridization of the DNAtolatently infectedganglion RNA wasdone as described in Materials and Methods. Si-resistant material was fractionated by
denaturing acrylamide gel electrophoresiswith EcoRI-Hinfl-digested pBR322 DNA as a size standard. An Sl-resistant fragment migrating with a size corresponding to 500 + 20 bases was seen in the sample hybridized with latently infected RNA (Fig. 1C, lane i). This band was completely absent inparallel tracks of DNAhybridized with uninfected HeLa cell RNA and with Escherichia coli tRNA, which served as controls (Fig. 1C, lanes ii and iii). We further confirmed this locationby using aprobe 5' end labeled at a
Hinfl sitethatislocated 150 bases3' ofthe capsite (data not shown). There were other bands of radioactive DNA resis-tant to Si digestion generated under the conditions of hybridization used (Fig. 1C). In each case, however, these bands were seen in both the control and ganglion RNA
tracks; therefore,they were not specific for latently infected
ganglion RNA. Although the reason for the occurrence of such nonspecific Si-resistant material has not been fully
investigated, itmay relate to thehigh G+C content of HSV DNA fromthisregion of thegenome. Whatever the source of thisbackground, its existence precluded the use of larger
[image:4.612.64.301.78.209.2]C
FIG. 3. Hybridizationin situ of 3H-labeledHSV-1DNAprobesto RNA inlatentlyinfected and uninfectedmurinespinalganglia.Probes
with the mappositions noted, preparedandhybridizedto tissue sections asdescribed in the Materials andMethods,wereused todetect
virus-specificRNA inganglionicneurons.(A) Probe, PstI-HpaI (0.775to 0.785m.u.),latentlyinfectedganglia.(B)Probe,SalI-BamHI(0.79 to0.807m.u.), latentlyinfectedganglia. (C) Probe,Hpal-PstI (0.764to 0.775m.u.),latentlyinfectedganglia. (D)Probe,BamHI-SstI(0.807 to0.815m.u.), latentlyinfectedganglia. (E) Probe, PstI-HpaI (0.775to 0.785m.u.),uninfectedganglia.(F)Probe,Sall-BamHI(0.79 to0.807 m.u.),uninfectedganglia. (G) Probe, Hpal-PstI (0.764to0.775m.u.), uninfectedganglia.
0.790- .
794-9i 0.7796
II'
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.68.558.433.671.2]probesforcarrying out a complete Si analysis ofthe HSV-1 LAT transcript.
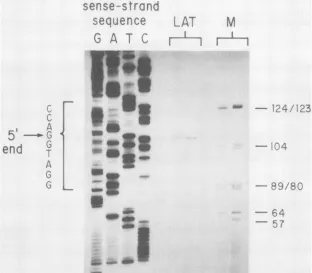
The location of the 5' endofthe HSV-1 LAT wasprecisely established by primer extension. Here, a 20-base primer complementary tothe direction oftranscriptionof the LAT
correspondingto aposition from 379to399 bases to theright (5') oftheKpnI(Asp718)site at 0.783m.u. wassynthesized by using the sequence data discussed in the next section.
This primer was annealed with total latently infected
gan-glion
RNA, andreversetranscriptase
wasusedtosynthesize
cDNAwith the 5' end of the LATasitstemplate.Atypical experiment is shown in Fig. 4; thesequenceseenis
comple-mentary to the mRNA coding (sense) strand. The only cDNA speecies ofany intensity consistently migrated at a
position which locatedthe cap siteatthe first G within the
trinucleotide AGG.Asshownin thesequencedatadiscussed below, this places theLAT cap site510bases to theleft of the KpnI site at 0.783 m.u. This result is in excellent
agreement with that determined by S1 analysis. Further,the
lack of any larger species confirmed our conclusion that
there is no readily detectable splice at the 5' end of the
HSV-1
from latently infected ganglia. PreliminaryRNA-primed dideoxysequence dataarefully consistent with the
location of thecap site shoWnin
Fig.
4(M. Flanaganand E.Wagner, unpublished data).
Analysis ofsequenceof HSV-1DNAencodingtheLAT. We nextdeterminedthenucleotidesequenceoftheDNA
encod-serse
-strand
sequence
G
A
T
C
6
I
A
5
-.G
end
rA
G
po wu
--L--~ ~
-9
40.
dw
4001l..W.dx
ing the HSV-1 LAT. A total of 2,400 bases from an SphI site located to theleft of the KpnI site at 0.783 m.u. to the SmiaI site at 0.794 m.u., 660bases to the rightof the Sall site at 0.790 m.u., are shown in Fig. 5. The 5' end of the LAT occurs at base 195. An RNA polymerase III B element
homology (7)occursbetweenbases 199and210,but there is no A element 5' of this region. The sequence 5' of the cap
site has some minimal features ofan RNA polymerase II promoter, with the cap about 25 bases 3' of the sequence
TTCAACAAA, which is a possible TATA box homolog.
Twoelements(CAGTAandCACT)atpositions109 and145, respectively, arepotentialCAAT boxhomologies.
There aretranslation initiationsignals in all threereading
frames throughout the sequenceofthe HSV-1LAT,butonly
twodisplaycodon usagepatterns which are at all consistent with their encoding a polypeptide, based on the analysis
methodof Fickett (5). Oneis the openreadingframe(ORF) initiated at base 625 with the nominal eucaryotic initiation
codon GATATGG, which remains open for 207 bases (67 amino acids). A second ORF encoding 305 amino acids
beginswith the ATG at base 1160,50 bases to the left of the Hpal site at 0.785 m.u. ThisORF extendstobase2074, 334
basestotherightof the Sall site at 0.790 m.u.;therefore,the 3' portionof this ORF iscomplementary tothe 3' 360 bases of the ORF for ICPO(18). The Northern blot data ofFig. 1 and 2indicate that the 3' end of the HSV-1LATextends as many as 50 bases to theright oftheSmaI siteat0.794 m.u.
A1
;.-
M
_
-.
1J4
/123
-"~Q9/30
- 64
--7
wo ,
FIG. 4. Primerextension localization of the 5' end of the HSV-1 LAT. As described in the text, a total of 20,ugof total RNA from latently infectedmouse sensory nerve ganglia was annealed with a primer of the sequenceTGGTGTGCTGTAACACGAGC which is complementary to alocation 395 to 415 bases to the left of theKpnI site at 0.783m.u. based on sequence data shown in Fig. 5. Reverse transcriptase was usedtosynthesizeacDNA fragmentwhichwas fractionated on a denaturing sequencing gel next to adideoxy sequence runfromthe same primer annealedtothesense(coding) strand of the DNA encoding the 5' end of the HSV-1 LAT. Two different preparations of RNA are shownin thetwolanes indicatedLAT. The four lanes of DNA sequence terminating at G, A, T, and C are shown.HaeIII-digestedpBR322 wasincludedas arough size marker in lanes M; note that in the gels shown here, these sizes only approximately correspond with the length of thesingle-strandedDNA fragments. Numbers to the right show sizes in nucleotides. The sequence of the sense (coding) DNA strand near the LATcapis also indicated.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.151.463.369.642.2]SphI
GCATGCTATG CGCTGCGTTT TTTTTTTTTT CTCCGTCGCC TTTCCTGTTC TCGCTTCTTC
CAAT?
CCCTCCCTTC CTCCCCCGTT ATCCCACTCG
Cap HSV-1 LAT
ACGCCGCGTT TCCAGGTAGG TTAGACACCT
AACGACAGGG GGCGCCCCAG TACAGCACAC CAGCCCGTTA CCCGTCCGAC CACCAACTGC GGCCGGTGTC GCTGTAACCC CCACCCCGCC TGTGTTTTTG GGAGTGCGGG GGAGGGGGGG TAAAGTTGTT CCTAAAGCGA CTCTGGCACG CGGGGGTGGG GCCGGCGCGG GTGCGCGTGC AACACCGCCC CCCCTCCTTC CGCCCATCCC CTCGTCTCCT GTTCCTGTTT CTGTCTCCCC GTGTGGCCGA CCCCCTTTTG HpaI GCCCTATAGT GTAGTTAACC GTCCGGGCGG GCAGGCGCTC I GCTCCATGCC AGTGGCAGGA HpaI(0.785 mu) TCTGTGGTTA ACACCAGAGC CCACGCACCC CCACTCCCAC ACTCCCACGC ACCCCCACTC CCCCACTCCC ACGCACCCCC GCACCCCCAC TCCCACGCAC AGGGAAAAGA TACAAAAGTA TTTTTTTATT AGGGCCAACA TCCACCTCCC CGACACGGAT CGACCCCCCC CTCCCCACAA SalI(0.790 mu) CGACGCCCCC TGCTCCCCGG GCCCACTAGG GTGCCCTGGT 1861 CCACTCCGAC 1921 GGGGAGCAGG 1981 TCCCGTCTGG 2041 GGGCAGGCAG 2101 GACCCCCGAG 2161 CGCGTGGCGC 2221 GCCGAGGGAG 2281 GGAAGCGGAG 2341 CGAGGAGGAA SmaI{ 2401 GGG GCGGGGGCCG _
______
_
AGGCTAAGGT TTTTCCCCCC CCCCTTATCT CCCACGCCAA TGCCTGGCTC40 50 60
TTTTTTTTTT TTTCTCGGTG TTCTGCCGGG
CAAT?
CCCCCCCCCC TTCACCCCCA GTACCCTCCT
TATA?
TCGAGGGCGC CCCGGTGTCG TTCAACAAAG
GCTTCTCCCC AATAGAGGGG GGGGGACCCA
Primer
CGGCCACGCC ACTCGCGGGT GGGCTCGTGT
CTCCCACCCT TAGTTAGACT CTGTTACTTA
AAGGGCCGGC TGGAAGACCG CCAGGGGGTC
TGACCCACGT ACTCCAAGAA GGCATGTGTC
TCTATGCTTG GGTCTTACTG CCTGGGGGGG
GGGTGTGGAA GGAAATGCAC GGCGCGTGTG TACCCCCCCC
I LAT ORF-1
GGATATGGAG GAGTGGCGGG TGCCGGGGGA CCGGGGTGAT KpnI(Asp7l8)(0.783 mu)
AAGGGTCGGG GGAGGGGGGG ATGGGGTACC GGCCCACCTG
CTTTGCACAC CAACCCCACG TCCCCCGGCG GTCTCTAAGA
SphI LAT ORF-1 ]
ATACCACCGA GCATGCCTGG GTGTGGGTTG GTAACCAACA
GTGATTCTCT GGCTGCACCG CATTCTTCTT TTCTAACTAT
CCCCACCCCT CCGCCCCACC CCCCAACACC CACGTCTGTG
GGCGCCCCGT CCCGCCCCGC TACCCCTCCC ATCCTTTGTT
CCCCCCCCCC CGCCCTTTGT GGCGGCCAGA GGCCAGGTCA
GCGGAAACTT AACACCCACA CCCAACCCAC TGTGGTTCTG
LAT ORF-2 SmaI
TGCTTTCGGG GATCGGTGGT CAGGCAGCCC GGGCCGCGGC
CTGCCCAACA Repeat Unit
t
TGGCACCCCC ACTCCCACGC ACCCCCACTC
GCACCCCCAC TCCCACGCAC CCCCACTCCC
CCACGCACCC CCACTCCCAC GCACCCCCAC
ACTCCCACGC ACCCCCACTC CCACGCACCC
CCCCACTCCC ACGCACCCCC GCGATACATC
AACCTTTATT TCCCAACAGA CAGCAAAAAT
CAAAAGACCC GCTGGTGTGT GGTGCCCGTG
TGGCTGGTGT AGTGGGCGCG GCCAGAGACC
[ ORF-ICPO
ACACGGGGGG CGTCCCTTAT TGTTTTCCCT
ACCACGGGTG CCGAGACCGC AGGCTGCGGA
CGAACAGCAT TCGGGTACTC GTGCGGCGGC TCCAGCCGGG TCCACCAGGA CCACGTACGC LAT ORF-2 TCCCCCGTGA TCGTCTTGTT ATGGGCAGGT AGCGCGTGAG GTCTTCCGGG CACACTTCCT GTTTCCTCTT GTCTCCCTCC GAGGACGCGG CCCCGGTGGC GAGGCGGAGG AGGAAGAGGC GTTCCCCACG GGGGGGCGTC GACCGCGGCC CCCGATGTTC ] CACGTAAGGC GCCGCCCGCG CGGCCCCCGC CAGGGCACCG GGAAGAGGTG GGAGGCCGCC GGGGTCATCC ACGTGGTTAC CGCAGCCGGG ACGCACCCCC TCCCACGCAC CCACTCCCAC CAACACAGAC CCCCTGAGTT TCTTTCACTT ACCCAGCGCC SmaI CGTCCCGGGT AGTCCAGGGC AGAGGCTGTT CCGCGGTCTC TCGCCATGTT CCCGTCTCCA TGTCCAGGAT GACAGGGCGA CCACGCTAGA GGGACGGCCC CGGAAGTCTC GGCCCAGAAG CAGCGCGGGG ACGGCCCCGC CCGAGGAGGC GCCCCCGCGG GAGTCGGGGC GAGGACGTCA GGGGGGTCCC
FIG. 5. Nucleotidesequenceof theDNAencodingthe HSV-1 LAT. Thesequenceof theKOS(M)strainof HSV-1between anSphlsite
708 basestothe left of theKpnIsite at 0.783 m.u.andanSmaI site 660 bases to therightof theSallsite at 0.790 m.u. is shown. Thecap site of the HSV-1 LAT is at base 195. A numberof other features discussed in the textare indicated. These include long stretchesof
pyrimidines(18to45,69to109,and 114 to137),apolymeraseIII B element(199to210),severalpossibleCAAT boxhomologies,and aweak
TATA boxhomology.Thesequencecomplementaryto theprimerusedforprimerextensioncorrespondsto bases 293 to312;the Hinflsite
used for Si analysisisatbase349. TheKpnIsite(0.783m.u.)is at base708, and there areHpaIsites at bases 1037 and1210, the latterbeing at0.785 m.u.TheSallsite at 0.790m.u.is at base 1740. The twopotentialORFs are between bases 625 and 831and 1160 and 2074.There
are15 repeats of thesequenceGCACCCCCACTCCCACbetween bases 1233and 1472. The translationalframe for ICPO ends at base1720 ontheopposite strand. Differencesbetween the sequences of the KOS(M)and17syn+ strains of HSV-1(Table1)areunderlined.
1199 1 61 121 181 241 301 361 421 481 541 601 661 721 781 841 901 961 1021 1081 .141 1201 1261 1321 1381 1441 1501 1561 1621 1681 1741 1801
10 20 30
on November 10, 2019 by guest
http://jvi.asm.org/
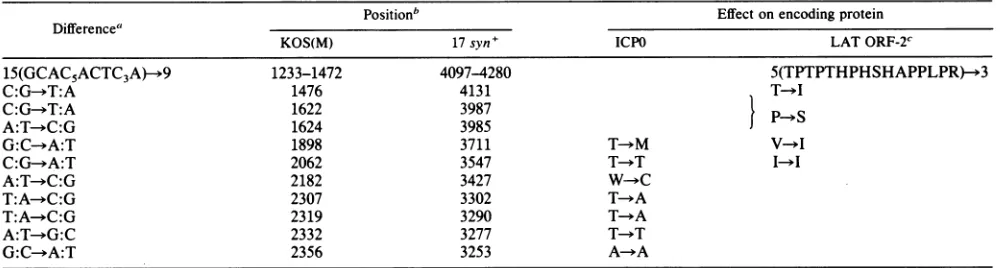
[image:6.612.137.478.48.666.2]TABLE 1. Strain differencesintheDNA sequenceencodingthe 3'portion of HSV-1 LAT
Positionb Effectonencoding protein
Difference"
KOS(M) 17syn+ ICPO LATORF-2C
15(GCAC5ACTC3A)-*9 1233-1472 4097-4280 5(TPTPTHPHSHAPPLPR)-*3
C:G--T:A 1476 4131 T YI
C:G->T:A 1622 3987 PS
A:T-*C:G 1624 3985
G:C-*A:T 1898 3711 T-*M V-*I
C:G--A:T 2062 3547 T-T I
A:T--C:G 2182 3427 W C
T:A-+C:G 2307 3302 T YA
T:A--C:G 2319 3290 T-*A
A:T-*G:C 2332 3277 T >T
G:C-*A:T 2356 3253 A-+A
a The base in the HSV-1 LATmRNA sense(coding)strand is shown for bothstrains;thebase inKOS(M)is shownfirst.
bThepositions oftheindicated nucleotides for theKOS(M)strainare asshown inFig. 5;thepositionin17syn+isasshowninreference18.
cIndicates changesinpredicted amino acid encodedby HSV-1KOS(M)to17 syn+. Sincethe sequenceof the ICPOgeneextending
through
the HpaIsiteat0.785m.u. has beenpublished forHSV-1 strain 17 syn+(18),
some comparisons are of interest and aresummarized in Table 1. The same ORF complementary to
that encoding the C-terminal
region
ofICPO is seen, and except where 9 sets of the repeat motif TGCGTGG GAGTGGGGGare seenin strain17 syn+, while 15occurinKOS(M), the homology between the strains is
high.
Tensingle-base differencesoccur: seven aretransitions and three are transversions. Five of the changes are unique to the
C-terminalportion oftheICP0-coding
region,
whilethreeareunique to the second ORFcontained in theDNA
encoding
the major LAT. Two base changes occur in the 360-base complementary overlap between the ICPO and the second LATORFs. Interestingly,theseresults inmoreconservative changesinthe aminoacidsequenceofthe second LAT ORF than in the ICP0 ORF. The samelevel ofhomology is seen between these twostrains ofHSV-1 to theleftoftheHpaI site at0.785m.u., basedon unpublished sequencedata(D.
McGeoch, personal communication). DISCUSSION
Inthis communication, we defined several properties of the LAT which we previously detected in murine sensory neuronslatently infected withtheHSV-1 strainKOS(M).
Of
particular interest in the presentstudy was adetermination ofLAT map coordinates on the viralgenome, the detailed characterization of the 5' end of the molecule, and the
analysis ofthe DNA sequence encodingthetranscript.
Ourprecise location ofthe 5' end ofthetranscript
com-bined with ourhigh-resolution Northern blot analysis indi-catethat the major HSV-1 LAT which is no more than 2.3
kilobasesfrom itsmigration on high-resolution gels is readily detected only with probes extending from bases 195 to 2400
(Fig.
5)
and is weakly detected with the DNA fragmentimmediately to the right of this (Fig. 2). The use of
Si
analysis to locate precisely the 3' end of the transcript as
describedinprevious publications (4, 6) was not particularly
fruitfulsince we did not obtain probes with very high specific activities. Preliminary analysis, however, does indicate that
probes 3' end labeled at the Sall site at 0.790 m.u. do not protectmorethan 700 bases of DNA (G. B. Devi-Rao and E.
Wagner,unpublished data). These considerations strengthen our
impression
based on Northern blot data such as shown inFig. 2that the 3' end of the major LAT maps quite close to theSmaI site at 0.794 m.u. The close correlation between
transcript size and sequence data suggests that any intron
withinthe majorHSV-1 LATmust bequite small.
The strong in situ hybridization in latently infected neu-ronsobtained withprobesthat detect the LATtranscripton Northernblotsconfirmsourearlier conclusion that the major LAT species is the most abundant transcript expressed in latent infection (29). A smaller and less abundant LAT
specieswasdetectedonlywithprobes ofthesamesenseand
positionof the viral genomeasthose usedtodetect themajor species. We suggestthat this smaller speciesisaprocessed
form of the major one. The fact that a very faint band of RNA ofthis approximate size isalso seen in the poly(A)+ fraction (Fig. 1B, laneiii) tendsto support this impression,
but the small amount of this smaller species precluded further characterization at this time. We have no evidence that any otherviraltranscript is expressedduringthe latent
phase ofinfection in murine sensory nerve ganglia.
Obvi-ously, however, low levels oftranscription cannot be
ex-cluded, particularly if these molecules are metabolically rapidly degraded aftersynthesis.
With respecttothecellular abundanceof LAT,ourRNA samples werederived from 20gangliathatcontainedatotal of 107 cells. Ofthese cells, roughly 3 x
105
-werelatentlyinfected neurons. Theintensity of
the
Northernblotsignalswascomparable to that seenfor
abu'ndant
HSV transcripts,such as VP5, isolated from 106 cultured cells [1 ,ug of infected-cell poly(A)-containing
RNA].
Therefore,the LATis present at an average level of abundance comparable to thatofabundant viraltranscripts isolated during productive infectionorto2 x 104to5x 104moleculesperinfected cell. This estimate was further confirmed by ourin situ hybrid-ization experiments. In these, equivalently labeled
gene-specific DNAprobes used todetectLATin latentinfection
and VP5 mRNAinproductive infection demonstrated equiv-alent signals (datanotshown).
The highlevels of the HSV LAT seen in latently infected murine sensory nerve ganglia are also seen in latently
infectedhuman (J. Stevens, L. Haar, D. Porter, M. L. Cook, and E. Wagner, submitted for publication) and rabbit (J.
Stevens, J. Hill, G. B. Devi-Rao, and E. Wagner, unpub-lisheddata)trigeminalganglia. In contrast to the abundance of the LAT transcript in these cells, recent work has established that the HSV-1 LAT is expressed at very low levels during productive infection of cultured rabbit skin cells (E. Wagner, G. B. Devi-Rao, Y.-F. Zhang, and J.
Stevens, manuscriptin preparation). Since only LAT of all
potential HSVtranscripts is expressed in the latent phase,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.59.561.89.223.2]promoter-control regions mediating its expression may have neuronal specificity. Inthis regard, further characterization
ofthe 5' end of the LAT will be required to identify fully the DNA sequences controlling its expression. However, it may be that the poor TATA and CAAT box homologies noted in the sequence shown in Fig. 5 have a role in the unusual expression of this transcript. Also, the several long stretches of pyrimidines seen (bases 16 to 43 and 59 to 135) may have some rolein tissue specificity.
Although we do not know the role of HSV LAT in latent infection, a mutant of HSV-1 (X10-13) with a deletion in the 5' end of the HSV-1 LAT and its control sequences has recently been characterized (R. Javier, V. Dissette, E. Wagner, and J. Stevens, manuscript in preparation). This virus establishes latent infections in mice yet expresses no detectable LAT or any other viral transcript, including the less abundant in situ signals mapping 3' to the LAT (Fig. 3D). This suggests that all RNA species seen in the latently infected neurons are related to the expression of the major LAT species and are not due to independent transcription units. It also demonstrates that expression of the LAT gene is not required to establish the latent state in mouse neurons. This does not, of course, exclude its acting to augment the normal restriction of the HSV lytic cycle in neurons.
As discussed in reference 29, one possible role of the major LAT species is to suppress the expression of the a
ICP0transcript to which it is partially complementary by an antisense mechanism. The ICP0protein is a transcriptional activator, and it was recently reported that HSV mutants lacking this gene replicate poorly at low multiplicities of infection (23, 30). Of equal interest, Schaffer and colleagues have found that twoICP0 deletion mutants cannot reactivate from the latent state in murine trigeminal ganglia even though viral DNA is present (D. M. Coen, D. M. Knipe, K. L. Taylor, and P. A. Schaffer, manuscript in prepara-tion).
The abundance and nuclear localization of the major HSV-1 LAT is consistent with itsfunctioning as anantisense regulator. An experimental system involving antisense reg-ulation which demonstrates mostoftheproperties described here has been reported (12). There thymidine kinase mRNA was associated with theantisense RNAandrestrictedtothe nucleus when RNA complementary to the 3' end of thymi-dine kinase mRNA was present. Unlike the results reported with that model, however,wedid notdetect any ofthe sense transcript (ICP0) in latently infected mice.
We consider it unlikely thatthe major species ofthe LAT simply encodes a protein functioning in the latent phase of infection. The physical location of the major LAT in the latent phase argues against its being translated. The posi-tions of the identified ORFs vis-a-vis the 5' end of the transcript, however, suggest that the major LAT could be spliced to generate a readily translatable mRNA. It should be noted thatthere is no identifiable polyadenylation signal within 900 bases ofthe 3' end of the major transcript (18; Y.-F. Zhang and E. Wagner, unpublished data). Although
the smaller andpoly(A)+ LAT species may be products of
splicing, theirlowabundancehas made fullcharacterization
impossible at thistime. Inpreliminary attempts to identify an LAT-encoded translation product, however, we have syn-thesized a 14-amino-acid peptide onthe basis of a predicted antigenicsequence withinthe secondHSV-1LATORF, and antiserum wasproduced inrabbits. Althoughthe antibodies were highly reactive against the peptide in enzyme-linked immunosorbent assays (titer, >1:10,000), we have not yet detected antigens in immunohistochemical tests performed
on acutely or latently infected murine spinal ganglia (J. Stevens, M. Cook, and V. Dissette, unpublished data).
In conclusion, whatever function the HSV-1 LAT per-forms in latent infection, it is clear that it is abundantly expressed in neurons. We have recently found that this transcriptis expressed only at low levels in infected cultured cells (Wagner et al., in preparation). A definition of those elements of the LAT promoter-control regionwhichmediate cellular specificity will be of great interest. In addition to these studies, we are currently investigating theexpression
of this transcript undervarious conditions of infection and, perhaps mostimportantly, studying its function in the latent state.
ACKNOWLEDGMENTS
We thank M. Rice and V. Dissette for helping with technical details.
This work was supportedby Public Health Service grantAI06286 from the National Institutes of Health to J.G.S., E.K.W., and L.T.F. Furthersupport was provided by Public HealthServicegrant CA11861 from theNational Institutes of Health, American Cancer Society grants MV159 and MV372, the University ofCaliforniaat Irvine Focused Research Program in the Molecular Biology of Eucaryotic Viruses(E.K.W.), Multiple Sclerosis Foundation grant RG-1647-A1 (J.G.S.), and Public Health Service grant A120953 (L.T.F.) from the National Institutes of Health. A.T.D. was sup-ported by molecular sciences training program grant GM08042; W.M.F. is a predoctoral trainee supported by training grant T32-CA-09054.
LITERATURE CITED
1. Anderson, K. P.,R. J.Frink, G. B. Devi, B. Gaylord,R.Costa, and E. Wagner. 1981. Detailed characterization of the mRNA mapping intheHindIllfragment Kregionof the herpessimplex virus type 1 genome. J. Virol. 37:1011-1027.
2. Deatly, A. M., J. G. Spivack, E. Lavi, and N. Fraser. 1987. RNA from an immediate-early region of the type 1 herpes simplex virus genome is present in the trigeminal ganglia of latently infectedmice. Proc. Natl. Acad. Sci. USA 84:3204-3208. 3. Denhardt, D. T. 1966. A membrane-filter technique for the
detection of complementary DNA. Biochem. Biophys. Res. Commun. 23:641-646.
4. Draper, K. G., G. B. Devi-Rao, R. Costa, E. Blair, R. L. Thompson, and E. Wagner. 1986. Characterization ofthe genes encoding herpes simplex virus type 1 and type 2 alkaline exonuclease andoverlapping proteins. J. Virol. 57:1023-1036. 5. Fickett, J. W. 1982. Recognition ofprotein coding regions in
DNA sequences. Nucleic Acids Res. 10:5303-5318.
6. Frink, R. J., R. Eisenberg, G. Cohen, and E. Wagner. 1983. Detailedanalysis of theportion oftheherpes simplex virustype 1genomeencodingglycoprotein C. J. Virol. 45:634 647. 7. Galli, G., H. Hofstetter, and M. L. Birnstiel. 1981. Two
con-served sequence blocks within eucaryotic tRNA genes are major promoterelements. Nature (London)294:626-631. 8. Haase, A.,M. Brodie, L. Stowring, and H. Blum. 1984.
Detec-tion of viral nucleic acids by in situ hybridization. Methods Virol. 7:189-226.
9. Hill, T. J. 1985. Herpes simplex viruslatency,p. 175-240. In B. Roizman (ed.), The herpesviruses, vol. 3. Plenum Publishing Corp., NewYork.
10. Inoue, T., and T. R. Cech. 1985. Secondary structure of the circular formofthetetrahymenarRNAintervening sequence: a
technique forRNA structureanalysisusingchemicalprobesand reverse transcriptase. Proc. Natl. Acad. Sci. USA82:648-652. 11. Kieff, E., T. Dambaugh, M. Hummel, and M. Heller. 1983. Epstein-Barr virus transformation and replication. Adv. Viral Oncol. 3:133-182.
12. Kim, S., and B. A. Wold. 1985. Stable reduction ofthymidine kinaseactivityin cellsexpressing highlevels of anti-sense RNA. Cell42:129-138.
on November 10, 2019 by guest
http://jvi.asm.org/
13. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual, p. 109-121. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
14. Maxam,A.M., and W. Gilbert. 1980. Sequencing end-labeled DNAwithbase-specific chemical cleavages. Methods Enzymol. 65:499-560.
15. McPheeters,D.S.,A.Christensen,E. T.Young, G.Stormo, and L. Gold. 1986. Translational regulation of expression of the bacteriophageT4lysozyme gene. Nucleic Acids Res. 14:5813-5826.
16. Messing,J., and J. Vieira. 1982. A new pair of M13 vectorsfor selecting either DNA strand of double-digest restriction frag-ments. Gene 19:269-276.
17. Peden, K., R. Mounts, and G. S. Hayward. 1982. Homology betweenmammalian cell DNA sequences and human herpesvi-rusgenomesdetected byahybridization procedure with high-complexity probes. Cell 31:71-80.
18. Perry, L.J.,F.J. Rixon,R. D.Everett,M.C. Frame, andD. L. McGeoch. 1986. Characterization of theIE110genes ofherpes simplex virus type 1. J.Gen. Virol. 67:2365-2380.
19. Puga, A., E. M. Cantin, andA. L. Notkins. 1982. Homology between murine and human cellular DNA sequences and the terminal repetition of the 5' component of herpes simplex virus type 1 DNA. Cell 31:81-87.
20. Puga, A., and A. L. Notkins. 1987. Continued expression of a poly(A)+transcript of herpes simplex virus type 1 in trigeminal ganglia of latently infected mice. J. Virol. 61:1700-1703. 21. Rock, D. L., and N. W. Fraser. 1983. Detection of HSV-1
genome in central nervous system of latently infected mice. Nature(London) 302:523-525.
22. Rock, D. L., and A. B. Nesburn. 1987. Detection of latency-related viral RNAs in trigeminal ganglia of rabbits latently infected with herpes simplex virus type 1. J. Virol. 61: 3820-3826.
23. Sacks,W.R.,and P.A. Schaffer. 1987.Deletion mutants in the
geneencoding the herpes simplex virus type 1immediate-early protein ICPO exhibitimpaired growth in cell culture. J. Virol. 61:829-839.
24. Sanger, F., S. Nicklen, and A. R. Coulson. 1977.DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
25. Spector, D. J., T. R. Jones, C. L. Parks, A. M. Deckhut,and R. W. Hyman. 1987.Hybridization betweenarepeated region of herpes simplex virus type 1 DNA containing the sequences
[GGC].
andheterodisperse cellular DNA and RNA. Virus Res. 7:69-82.26. Spivack, J. G., and N. W. Fraser. 1987. Detection ofherpes simplexvirus type1transcripts during latent infectioninmice. J. Virol. 61:3841-3847.
27. Stevens, J. 1980. Herpetic latency andreactivation, p. 1-11. In F. Rapp (ed.), Oncogenesis and herpesviruses, vol. 2. CRC Press, Inc., BocaRaton, Fla.
28. Stevens, J. G., and M. L. Cook. 1971. Latentherpes simplex virus inspinal ganglia ofmice. Science 173:843-845.
29. Stevens, J.G., E. K.Wagner, G.B.Devi-Rao,M. L.Cook,and L. Feldman. 1987. RNAcomplementaryto aherpesvirus alpha genemRNA ispredominant in latently infected neurons. Sci-ence235:1056-1059.
30. Stow,N. D.,and E. C. Stow. 1986. Isolation and characteriza-tion of a herpes simplex virus type 1 mutant containing a deletion within the geneencoding theimmediate-early peptide Vmw110. J.Gen. Virol. 67:2571-2585.
31. Stroop,W.G.,D. L.Rock,and N. W. Fraser.1984. Localization of herpessimplex virus in the trigeminal and olfactory systems of themouse central nervous system during acute and latent infections byinsituhybridization. Lab. Invest. 51:27-38. 32. Thompson,R. L.,M. L. Cook, G.B.Devi-Rao, E.K.Wagner,
andJ.G. Stevens. 1986. Functional andmolecularanalyses of theavirulentwild-typeherpes simplex virus type1 strainKOS. J. Virol. 58:203-211.