Copyright( 1973 AmericanSocietyforMicrobiology Printed in U.S.A.
Selection of
Temperature-Sensitive
Mutants
During Persistent
Infection: Role in Maintenance
of
Persistent Newcastle Disease Virus
Infections
of L
Cells
OLIVIA T. PREBLE ANDJULIUS S. YOUNGNER
Department of Microbiology, School of Medicine, University of Pittsburgh, Pittsburgh, Pennsylvania 15261 Receivedforpublication 30March 1973
Virus mutants
(NDVpi)
recovered from L cells persistently infected with Newcastle disease virus (NDV, Herts strain) are temperature-sensitive (ts) at43 C, although the wild-type virus (NDVO) which initiated the persistent infection replicates normally at that temperature. To study the relationship between thets marker of
NDVpi
and the otherproperties which distinguish this virus from NDVO,NDVpi
ts+ revertants were selected at the nonpermissive temperatureandNDVOtsmutantsweregenerated by treating NDVOwith nitrousacid.Spontaneously-occurringtsmutantsin theHerts NDVpopulationwerealso
isolated. The different viruspopulationswerecharacterizedwithregardtoplaque size, virulence foreggs, and thermalstability of infectivity, hemagglutinin, and
neuraminidase. The
NDVpi
ts+revertants,althoughnolongertemperature-sensi-tive, retained
NDVpi
properties, whereas both spontaneously-occurring and mutagen-inducedtsmutantsremainedwild-type in their other properties. These findings showed that the properties which characterizedNDVpi
wereindepend-ent of the ts marker. However, the ts marker and the other markers of
NDVpi
were coselected during the persistent infection, and the combination of thosemarkers appearedtobeimportant in theoutcome of NDV infection of L cells.
NDVpi
replicated productively in L cells, whereas NDVO, theNDVp0
ts+ revertants, and the spontaneously-occurring ts mutants all yielded covertinfections in L cells.The role of the selection ofts mutantsin persistent infection
was confirmed as follows: L cells were persistently infected with
NDVpi
ts+revertants andNDVOts mutants. Virus recoveredfromthe persistently infected cultures aftereight cellpassages wasalways temperature-sensitive and of smaller
plaque size than the parental virus in chicken embryo cell cultures. Similar results were obtained with virus recovered from L-cell cultures persistently infected with two other velogenic strains of NDV, the Texas-GB and Kansas-Man strains. Theseresultsstrongly suggest that selection ofts mutants during the persistent infection was not random and played a role in establishmentor
maintenance of the persistent infection, orboth.
Henle and his co-workers (7) demonstrated that persistent infection of mouse Lcellscould bereadilyestablished with theVictoria strain of Newcastle disease virus (NDV). Early experi-ments indicated that the virus re-isolated from the persistently infected cultures differed from the parental virus in both plaque type in permissivechicken embryo(CE) cells (7) and in virulencefor Lcells (15).Thacore and Youngner (23) found that stable small-plaque mutants could be isolated from mouse L cells (LND\)
persistently infected with the Herts strain of NDV and thesemutants
(NDVpi)
differedfrom the parental Herts virus (NDVo) in several respects: plaque size in CE cells, virulence for chicken embryos, virulence for L cells, and thermal inactivation profiles of several virion-associated activities (23-25). In addition, all of the HertsNDVpi
clones were shown to be temperature-sensitive (ts) mutantswith defects associated with RNAsynthesis at the nonper-missive temperature (10). Complementation481
on November 10, 2019 by guest
http://jvi.asm.org/
could not be detected between any
NDVpi
clones primarily as aresultofviralinterference at an early step in replication at either the permissive ornonpermissivetemperature (11).
The present report deals with therelationship between the temperature-sensitive marker of
NDVpi
andthe other properties whichdifferen-tiate
NDVO
andNDVpi.
The followingtypes of virus were compared:NDV,i
ts+ revertants selected at the nonpermissive temperature; NDVO ts mutants generated by treatment with nitrous acid; and spontaneously-occurring ts mutants in populations of Herts NDV. The properties of each type of virus were character-ized by the methods used previously to defineNDVp,
andNDVo
(10, 23).To determine if selection of ts mutants of NDV is involved in the establishment and maintenanceofpersistent infection, the ability
of
NDVpi,
NDVpi
ts+ revertants, and NDVO tsmutants to initiate persistent infection in L cells was tested. In addition, persistent infec-tions established by using two other strains of NDV, theTexas-GB and Kansas-Man strains, yielded temperature-sensitive mutants and the properties of the viruses selected in these per-sistent infections are also reported.
MATERIALS AND METHODS Cells. Primary CE cell cultures and acontinuous lineof'mouse L (clone 929) cells werepropagatedand maintained in Eagle minimal essential medium (MEM) plus4%calfserum (27).
Viruses. The origins and properties ofNDVO and the NDVpi clones derived from the Herts strain of NDV have been described in detail previously (10, 23). Two velogenic NDV strains, Texas-GB and Kansas-Man, were obtained from Samuel Baron, National InstitutesofHealth.
Isolation of NDVpi ts+ and NDVo ts clones.
NDVp,
ts+ revertants were cloned as follows: Stockpreparations ofNDVpi clones20, 25, 29, and38 were diluted in medium and assayed at 37 and 43 C. Well-isolated plaques obtainedfromplatesincubated atthe nonpermissive temperatureweretransferredto CEcell monolayersin 2-ozprescription bottles.After 24to 48hat37C, when cell destruction wascomplete, infected fluids were harvested and assayed for infec-tivity at 37 and 43C. Clones with a high plaquing efficiency at43 C (43C titer/37 Ctiter > 10-1)were plaque purified once again. Working stocks of five clones isolatedbythisprocedurewerepreparedinCE cell monolayer culturesin 32-ozprescription bottles. Mutagen-induced NDVO ts mutants wereisolated after treatment of NDVO with NaNO2 in acetate buffer, pH4.7, accordingtothemethod describedby Granoff(6). Well-isolated plaques were picked from terminal dilution plates of samples showing about 105 survival, transferred to CE cell monolayers in 2-oz bottles, and the virus progeny were tested for plaquing ability at 37 and 43 C. Working stocks of
fourcloneswith lowplaquingefficiencies (<10-2) at
43C were prepared in CE cell monolayers in 32-oz bottles.
Spontaneous NDV ts mutants were isolated from uncloned Herts NDV. Infected allantoic fluid was
seriallydiluted and plaquedat high dilutionsonCE cellsat37C. The agaroverwell-isolatedplaqueswas
removed,transferredto1.0ml ofMEM,and thevirus
was allowedtoelutefrom the agarat 4C overnight.
Hertsclones (351) wereisolated and screened bythe
procedure described below.
Screening ofNDV clones for temperature-sen-sitivity. CE cell monolayers were grown in plastic
trays(disposo-trays,Linbro ChemicalCo.)containing
24 wells of 24-mm diameter. Each well was seeded with1.5ml ofastandarddilutionofprimary CEcells;
about 106 to 2 x 106 cellswere presentin each well when themonolayerswereconfluent. The cells in each well were infected with 0.1 ml of undiluted virus
suspension from isolatedplaques.CE cells in control wells were mock-infected or infected with NDVo or
NDV,i
clone 35. Replicate welled-trays were in-cubatedat37and 43 C.Theinfectedmonolayerswereexamined under a microscope for cytopathology (CPE) which correlates well with virus replication (Youngner, unpublished data); cultures with CPE
always showed increased virus production, whereas cultures without CPE were usually found to have
producednovirus progeny. When CPEwascomplete
inwellsincubatedat37Candabsentin thereplicate
wells incubated at 43 C (usually 24-48 hafter infec-tion), fluids were harvested from both wells and assayedat37C.Virus cloneswithreplication efficien-cies of10-2orlessat43Cwereconsideredpotentialts mutantsandworkingstocks of viruswerepreparedin CE cells.
Infection of CE or L cells. The procedures for infectingcells in monolayer cultures and for harvest-ing released (RV) and cell-associated virus (CAV)
used in the experiments reported here have been described in detail elsewhere (10). Briefly, virus inoculumwasadsorbedtoCEorL-cellmonolayers for 1 h at 37 C. The infected monolayers were washed twice, fed with fresh medium, and incubated atthe appropriate temperature. At various times, culture fluids(RV)orinfected cellswereharvestedandvirus infectivity was determined by plaque assay in CE cellsat37 C.
CharacterizationofNDV clones.The characteri-zation of the NDV,i ts+ clones and the NDVO ts mutants wasperformed by methods used previously
to differentiate NDVpI and NDVO (10). Thethermal inactivation ofviral infectivity, hemagglutinin, and neuraminidase activities was determined bydiluting stock virus preparations into phosphate-buffered sa-line (PBS, 0.1 M, pH 7.2), and heating at the appropriatetemperature (10, 23, 25).
Establishment ofpersistently infected L cells. L-cellmonolayer cultures (5 x 10' to 6 x 106 cells) in 2-ozprescription bottleswere infected with0.1 PFU/ cell oftheappropriateNDVclones. Thecultures were washed twice, fed with 7 ml of warm MEM, and incubated at 37 C. This was considered "passage zero"(P0). Cultures withnoobviouscytopathology by 72h weresubcultured(P,) by scraping thecells from J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
the glassinto 2 ml of freshMEMandtransferring0.3 to0.4 ml of thiscellsuspension (5 x 105to 10' cells) into new 2-oz bottles. The infected cultures and
mock-infected control cultures were subcultured every7datysafter that(P2, P3, etc.). Bothsupernatant
fluids and cells were assayedfor infectivecenters at
the time ofsubculturing. At PO, P,, P4, and
P.,
the interferon activity in supematant fluids was alsodeterminedbytheplaque-reduction method(27). To confirm the establishment of persistent infec-tions, cultures at P.were challenged with vesicular stomatitis virus(VSV) (MOI = 0.1PFU/cell)orwith
NDVp, clone 35 (MOI = 5PFU/cell). Cultures were
infected with challenge virus for1 h at37C, rinsed, and fed. Culture fluids harvested 24 h later were
assayedonCE cells. VSVplaques werecountedat 2 days and were easily distinguishable from NDV
plaques counted3to4days afterinfection.
The reisolation ofclones from LND1' cultures was
also performed at P.. Cultures were assayed for
infectivecentersatthetime ofsubculturing for
P,,
the plaque size was measured, and the agar overwell-isolated plaquesfromtheassayplateswasplacedinto
1.0 ml of MEM at 4C overnight. The virus eluted fromtheseplaqueswasscreened for
temperature-sen-sitivity by the "welled-tray" technique described above.
RESULTS
Isolation of
NDVpi
ts+ revertants and NDVo ts mutants. The objective of this set of experimentswastodetermineifthetsmarker ofNDVpi
waslinkedtoanyof theotherpropertiescharacteristic of
NDVpi.
To obtainNDVpi
ts+ revertants, stock preparations ofNDVpi
wereplated on CE cells, overlaid with agar and
incubated at 43 C. If
NDVpi
plaques whichappear at the nonpermissive temperature are
due torevertantsrather thantoleakiness, then isolation of
NDVpi
plaques formed at 43C should result in selection of ts+ clones. From cultures plated with clone 38, 6 plaques wereisolated and fromplates infected with clone 20, 10plaqueswereisolated. Of these 16plaques, 4
(NDVpi
ts+ 20R3, 38R1, 38R4, 38R6) hadrepli-cationefficiencies at43Cgreaterthan 10- and
were kept for further study.
NDVO ts mutants wereisolatedafter treating
NDVo
stocks with nitrous acid as described inMaterials and Methods. The kinetics of virus inactivation by HNO2wereverysimilartothose obtained by Granoff (6). In three experiments, 70plaqueswerescreenedfor
temperature-sensi-tivity and 4 clones (NDVO ts 17, ts29, ts31, ts
43) had replication efficiencies of less than 10-1 at43 C.
Theplaquing efficiencies of cell culture
prep-arations of the
NDVp1
ts+ revertants and the NDVots mutantsarecomparedtotheplaquing efficiencies of allantoic fluid preparations of parentalNDVpi
and NDVO (Table 1).Revert-ants from both
NDVpi
clones 20 and 38 were clearly no longer ts, whereas the nitrous acid-induced NDVO ts mutants were ts to varying degrees. Three of the NDVO ts mutants pro-duced largeplaquesinCE cellsat 37 C,whereas NDVO ts 31 produced small plaques under the same conditions. All of theNDVp,
ts+ clones retained thesmall-plaquecharacteristicatboth 37and43 C. Plaquesizeand temperature-sensi-tivitytherefore appear to be independent char-acteristics.Thegrowth patternsofthe
NDVpi
ts+ revert-ants and the NDVO ts mutants at 37 and 43C were confirmed by one-cycle growth curves in CE cells. The growth curvesofNDVO ts 17 are showninFig.1;the resultswiththeother NDVO ts mutantswere essentially thesame. Replica-tion curves at 37 C were very similar to those previouslydescribedforNDVO (10, 23).At 43 C, however, no replication ofNDVO
ts 17 was detectable, although NDVO replicates normally at 43 C. Thereplication
ofNDVpi
ts+ 38R6 in CE cells at 37 and 43C is shown in Fig. 2. At 43 C, significantvirusreplicationwasobserved, with peak virus production (-30% ofthe 37 C yield) occurring12h after infection.The results also suggest that the ts+ revertants may be defective in a maturation step at 43C since mostofthevirus remains cell-associated.Properties of
NDVpi
ts+ andNDVO
ts clones. The new virus isolates were character-izedtodetermineif reversionofthetsmutationof
NDVpi
or introduction of a tsmutation intoTABLE 1. Plaquing efficiencies ofNDVpits+ revertantsandNDVomutantsinCE cells at37and
43C
PFU/ml Plaquing
Virus efficiency
37 C 43 C (43C/37C)
NDVOa 4.7x 10' 1.7 x10' 0.36
NDVpi Cl20b 4.0x 10' 7.7x 10' 1.8x 10-4
NDVpiCl38 1.5x 10' 1.0x 107 6.7x 10-9
NDVP1
ts+
20R3c 1.5x 106 6.1 x105
0.41NDVPI
tS+38R1 6.6x 106 5.1x 10' 0.78NDVpi
tS+38R4 1.0x107
6.1 x 106 0.61NDVpits+38R6 7.9x 101 6.4x 10' 0.84 NDVOts17d 3.2x 107 3.3x 102 1.0x 10-5 NDVOts29 1.4x 107 1.0x 10J 7.2x 10-5 NDVOts31 3.4x 101 3.5x 104 1.0X
10-i
NDV.ts43 1.1x 107 1.5x
105
1.4x 10-2aWild-type cloned Herts NDV, infectedallantoic
fluid.
'ts Mutantfrom persistent infection, infected al-lantoicfluid.
cts+ Revertants isolated from
NDVp1
clones, in-fected CE cellculture fluid.dtsMutants fromNDVo treated withnitrousacid, infectedCE cell culturefluid.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.255.451.421.654.2]108
temperature
and assayed for residual activitycompared
tounheated controls. TheNDVpi
ts+ revertantsremained"NDVi'"
despite
reversion towild-type
withregard
toreplication
at 43C;
7 / virus
infectivity
was not stable toheating,
07 whereas both hemagglutinin (HA) and
neura-minidase activitieswerestable underconditions that
completely destroyed
the HA andneura-minidase
activity
of NDVo(Table
3).
In aE6 separate experimentwith the NDVOtsmutants,
10 the results showed that these mutants remained
"NDV," even
though they
could nolonger
replicate
at 43 C.Clearly,
the ts mutation ofNDVpi
can revertindependently
fromthemuta-s ~
tion(s)
which result in altered thermalstability
lo % of virion-associated activities. Conversely,
in-troduction of a ts mutation into NDVO was not
sufficient to
change
thesephysical properties
of the virion.4'
Replication
ofNDVpi
ts+ andNDVo
ts 2 4 6 8 0 12 14 6 18 20 22 24 clones in L cells. To determine iftemperature-HOURS AFTER INFECTION
sensitivity
was linked to theabilityofNDVpltoFIG. 1. Single cycle yield ofNDVOts17 inCEcells
108
at 37 and 43C. CE cell cultures were infected and 0 . incubated at 37 and43 C. At various times, RV(0,0) and CAV (A, A) were determined. Results with NDVOwere very similarto those obtainedpreviously
(10). Filledsymbols, 37C; opensymbols, 43C. 7
10
NDVO altered the other
physical
orbiological
properties of the virus. Previous
experiments
(10, 23) have shown that NDVO is more lethal
for fertile eggs than
NDVpi.
As shown inTable 62, the
NDVpi
ts+ clones tended to behave like -\NDVpi
and the NDVO ts mutants tended to _behavelike
NDVo, although
the correlation was Enot absolute (e.g., clones ts+ 38R1 and ts
31).
X This suggeststhatvirulenceforeggs maynotbe 5related to
temperature-sensitivity.
Also,
no ab-solute correlation exists in this case between plaquesize oftheinfecting
virusand its lethal-ity for eggs,although
this correlation has beenreported for other NDV strains (13,
16).
4 Dead and live eggs were harvested and the 10 virus in theinfectedallantoic fluidwasassayed
at 37 and43 C. With all clones except NDVOts31, virus from infected eggs wasofthe
expected
plaque size and
temperature-sensitivity,
and 3the allantoic fluids were used as a source of 2 4 6 8 10 12 14 16 18 20 22 24
virus for subsequent experiments.
NDVo
ts 31was not stable and was not used in further HOURS
AFTER
INFECTIONexperiments. FIG. 2. Single cycle yield of NDVj, ts+ 38R6 in CE NDVO and
NDVpi
also differmarkedly
in cells at 37 and 43 C. CE cell cultures were infected thermal inactivationprofiles
of virion- withNDVO
or ND V, ts+38R6 and incubated at 37 orassociate
activities.
To det e te. 43 C. At varioustimes,
RV(0, 0) and CAV (A, A)associatedl activities. TO determine the were harvested andassayedonCEcells at 37C. The
thermostability
profiles
ofNDV,j
ts+revertants results withNDVowere very similar to those obtained and NDVOts mutants, stockviruspreparations
previously (10). Filled symbols, 37 C; open symbols, diluted in PBS were heated at theappropriate
43 C.484 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.57.250.58.308.2] [image:4.495.263.452.312.586.2]replicate efficiently in mouse L cells,
single-cycle yield experiments with the ts+ revertants
were performedinmouseLcells infected with1
to5PFU/cell. The results with
NDVpi
ts+38R4,NDVo,
andNDVpi
clone 38areshown in Fig. 3A,B,C, respectively. The results with the other ts+ clones were not significantly different from those obtained with ts+ 38R4. The data shown
for NDVOand
NDVpi
were verysimilartothosepreviously published (10, 23). The growth
curves of the ts+ clones were intermediate
between those of NDVO and
NDVpi,
with the overall shape of thecurve moresimilartothat of NDVO. The large accumulation of CAV, charac-teristic of the covert infection of L cells by NDVO, wasalsoseen in Lcells infected with ts+ 38R4. Less than 5% of the CAV present in infected cells at 8 to 12 h after infection was [image:5.495.254.449.83.243.2]released into the medium by 24 h after infec-tion. With
NDVpi
clone 38,however,peak virusTABLE 2. Lethality ofNDVP, ts+revertants and NDVO ts mutants for embryonated eggsa
No.dead/no.ofeggsinoculated Virus
24h 48 h 72h
NDVO 1/27 27/27
NDVPiCl20 1/24 7/24 14/24
NDVpits+ 20R3 0/24 0/24 17/24
NDVPiCl38 0/24 2/24 18/24
NDVpi
ts+38R1 0/24 13/24 24/24NDVpits+38R4 0/24 1/24 24/24
NDVpi
ts+38R6 0/24 2/24 20/24NDVO tS17 1/24 17/24
NDVOts29 0/24 23/24
NDVOts31 0/24 0/24 15/21
NDVOts43 0/24 24/24
aEaCh of 2 dozen eggs was
inoculated
into the allantoic cavity with 104 PFU of virus. Theinfected eggswereincubated at 36 C and cumulative dead eggs wererecorded at 24, 48, and 72 h. [image:5.495.101.397.292.436.2]4 a 12 16 20 24 4 8 12 16 20 24 4 8 16 20 Z HOURS AFTERINFECTION
FIG. 3. Replication ofNDVO, NDVp1 ts+38R4andNDVpiclone 38 in mouse L cells. MouseL-cell cultures were infected(MOI = 1-5PFU/cell) and incubated at 37 C. At various times, RV (0) and CAV (A) were de-termined. A, NDVpits+38R4; B,NDVo; C, NDVpiclone 38.
TABLE 3. Thermostability ofvirion-associated activities ofNDVpits+revertants andNDVO ts mutants
Experiment Fractionsurvivingainfectivityafter: Fractionremaining Fractionremaining
ExPeriment Virus HAUafter35min neuraminidase after
no. 30min 60
min
at 55 Cb 35min
at 55Cb1 NDVO 0.36 0.15 <0.2 <0.1
NDVpiCl20 3.1x 10-3 3.8 x 10-4 0.5 0.80
NDVP4
ts+20R3 4 x 10-3 4x 10-4 NDC NDcNDVp,
Cl38 1.9x 10-2 4x 10-3 0.71 1.0NDVpits+ 38R1 0.022 1 x 10-3 1.0 2.5
NDVpits+38R4 0.01 1.5x 10-3 1.0 2.5
NDVpits+38R6 8.7x 10-2 8.9x 10'- 0.7 0.95
2 NDVO 0.75 0.35 <0.2 <0.1
NDVots17 0.64 0.12 <0.2 0.086
NDVots29 0.41 0.15 <0.2 0.405
NDVots 43 0.67 0.21 <0.2 0.162
aCEcell-grownvirusdiluted 1:10inPBS andheated for 30 or 60 min at 50 C. bEgg-grown virusdiluted1:50 inPBS and heated for 35 min at 55 C.
cNotdetermined; titer ofegg-grown virus was not high enough to perform these tests.
.. 11 - 1.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.495.49.449.484.638.2]production did not occur until 16 h
postinfec-tion and 60% of the accumulated CAV was
released by 24 h after infection. These results suggested that the ability of
NDVpi
toreplicate in L cells may be linked to the ts defectofthevirus.
A similar experiment with the nitrous acid-inducedmutants,however, indicated that thets
marker alonemaynotbe sufficient fora
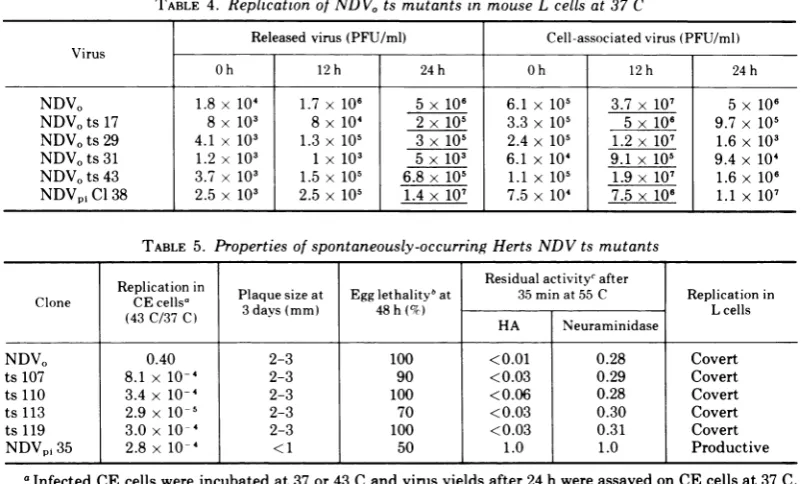
produc-tive replication cycle of NDV in L cells. The NDVOtsmutants,like the parental NDVO virus, alsoproduced acovertinfection in Lcells using
an input MOI of 1 to 5 PFU/cell (Table 4). A
comparisonofthe underlinedvalues for RV and CAV for each virus listed in Table 4 revealsthat with both NDVO and each of the NDVO ts mutants, accumulationofCAVwasmaximalat 12 h after infection, and that less than 10% of thisinfectivity wasreleased as maturevirus. In allcases,CAV declinedbyatleast50% between 12 and 24 h after infection without a concomi-tant increase in released virus. The fate of the CAV in the covertinfection of L cellsby NDVO, NDVO ts mutants or
NDVpi
ts+ revertants remains unknown.Properties of spontaneous ts mutants. To investigate further the relationshipbetweenthe
ts marker and the evolution of
NDVpi,
351 clones of Herts NDVwereisolated and screenedby the welled-tray technique for ability to
replicate at 43C. This technique determines the number of ts clones present in the virus population tested but does not measure the
mutationrateofthis marker. Fourspontaneous ts mutant clones were obtained. The reversion
frequencyof thets markerforthese clones was
<3 x 10-5 in all four cases. The replication efficiencyinCEcellsat43 Cforeach clone and other properties ofthe spontaneously-occurring
mutants are shown in Table 5. All of the
spontaneous mutants could be classified as
"NDV0": they produced large (2-3 mm) plaques in CE cells at 37 C; the HA and neuraminidase were not stableto heating, and they produced a covert infection in mouse L
cells in whichalargeamountofreplicatedCAV
was neverreleasedinto the medium. Thesedata
confirm that withnaturally occurring clones,as
well as with mutagenized clones, no
relation-ship exists between temperature-sensitive markers and other properties associated with
NDVp0.
One other result is worth noting. The
spon-taneous ts clones were tested for the abilityto
synthesize virus-specific RNA in CE cells at 43 C by using methods which have been de-scribed in detail (10). All four spontaneous mutantsfailedtosynthesize virus-specificRNA TABLE 4. Replication ofNDVO ts mutants in mouse L cells at 37 C
Released virus(PFU/ml) Cell-associated virus (PFU/ml) Virus
Oh 12h 24h Oh 12h 24h
NDVO 1.8x 104 1.7 x 106 5 x 106 6.1x 105 3.7x 107 5 x 106
NDVots 17 8 x 103 8x 104 2x 105 3.3 x 105 5x 106 9.7 x 105
NDVots29 4.1 x 103 1.3x 105 3 x 105 2.4x 105 1.2 x 107 1.6 x 103
NDV0ts31 1.2x 103 1x 103 5 x 103 6.1 x 104 9.1x 105 9.4 x 104
NDVOts 43 3.7x 103 1.5 x 105 6.8 x 105 1.1 x 105 1.9x 107 1.6 x 106
NDVpiCl38 2.5x 103 2.5 x 105 1.4 x 107 7.5 x 104 7.5 x 106 1.1 x 107
TABLE 5. Properties of spontaneously-occurring HertsNDVts mutants
Replicationin Residualactivityeafter
Clone CEcells" Plaquesizeat Egg
lethality"
at 35minat 55 C Replicationin3davs (mm) 48h(%) Lcells
(43C/37C) ~~~
~~~~~HA
NeuraminidaseNDVo 0.40 2-3 100 <0.01 0.28 Covert
ts 107 8.1 x 10-4 2-3 90 <0.03 0.29 Covert
ts 110 3.4x 10-' 2-3 100 <0.06 0.28 Covert
ts 113 2.9x 10-5 2-3 70 <0.03 0.30 Covert
ts119 3.0x 10'4 2-3 100 <0.03 0.31 Covert
NDVpi35 2.8 x10-4 < 1 50 1.0 1.0 Productive
aInfectedCE cellswereincubatedat37or43C andvirusyieldsafter24hwereassayedonCEcellsat37C. bEachof2dozenfertile eggswasinjectedintotheallantoiccavitywith 104 PFUand dead andlive eggswere
tallied after48h.
cStock virus preparations were diluted in PBS, heated at 55C and residual hemagglutinin and neuraminidase activitieswere assayedaspreviously described (10).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.495.54.455.383.625.2]at the nonpermissive temperature. In addition to the four spontaneous ts mutants, we have nowtested three nitrous acid-induced mutants,
and atleast 20ts mutantsisolatedfromLcells persistently infected withHerts NDV and allof thesets mutantswereRNA-.Apossible
excep-tion was NDVO ts 43 which synthesized about 16% of the wild-type level of RNA at 43 C; however,the reversionfrequencyof this mutant
wasfairly high, and the levelofRNAsynthesis obtainedmay have been duetothepresenceof revertants.
Persistent infection with
NDVpi
ts+ revert-ants andNDVotsmutants. The results ofthe experiments withNDVpi
ts+ revertants, NDVO ts mutants generated with nitrous acid, and spontaneousHertsNDVtsclones, coupled with previous results withNDVpi,
strongly suggested that selection ofts mutants within the persist-ent infection wasnotrandom andmight playarole in either establishment or maintenance of
the persistently infected state. To investigate this possibility, L-cell cultures were infected with NDVO,
NDVpi, NDVpi
ts+, and NDVo ts mutant clones, by using procedures similarto those used for establishment of the original LNDV cell cultures in 1967 (see Materials andMethods). Seventy-two hours later, all infected cultures appeared healthy and were subcul-tured
(Pj)
into new 2-oz bottles. The infected cultures retained their normal appearance and have been subcultured at the same rate ascontrol L cells once per week for over 30 passages.
Pertinent data on the early passages ofthe infected L cells are given in Table 6. Assay of
infected centersincellsand culture fluids from P0revealed thatverylittle, ifany,virus
replica-tion had occurred during the first 72 h of infection. In most cases, interferon synthesis
wasminimalduetothelow MOI(23).AfterP0,
most of the virus in the persistently infected cultures remained cell-associated and this was
reflected in the valueforcells perplaque. This
value fluctuatedwidely from sample tosample and from week to week, but average values
indicated that one cell in 100 or in 1,000 was
usually capableofyieldingvirus. These values
areinagreementwiththosepreviouslyreported forLNDV cultures (23).
By
P,,,
theoriginal virus inoculum hadbeen diluted by cellpassage appoximately 10.5logs. Since the original virus inoculum was 0.1 PFU per cell or about 3 x 105 PFU per bottle,infectivity atP15 wasclearly the result of virus
replication. The notation L(virus clone) was
adopted to denote each type of persistently infected culture, so that
LHertr
and LS8R1 werepersistentlyinfected with uncloned Herts virus
and
NDVpl
ts+38R1, respectively.Persistent infection, therefore, can
appar-ently be established by all NDV clones except
NDVpi. NDVpi
clone 20 induced significantlevels of interferon during P0 and was rapidly
eliminatedfromthecultures (Table 6); after
P,
the cultureswere consideredno longerinfected
and were discarded. Disruption of the cells by
sonic oscillation priortoplating hadnoeffecton
the number of infective centers per culture.
Other attempts to establish L-cell cultures persistentlyinfected with
NDVp1
have also been unsuccessful (Thacore and Youngner,unpub-TABLE 6. Earlypassagehistory ofL cellspersistentlyinfectedwith various typesofNDV
P 0 P, P15
Lcells in- PFUa/ml Cells"/ Interferon PFU/ml Cells/ PFU/ml Cells/
fected with: x10' plaque titerc x 10 plaque x 102 plaque
Herts 2.6 3.0x 102 <32 <1 1.3x 10' 0.2 3.6x 10'
NDVO 2.5 3.0 x 102 <32 <1 5.0 x 103 3.3 4.0x 102
NDV.ts17 4.5 1.0X 102 <32 <1 5.0x 103 0.2 3.7x 101
NDV.ts29 4.0 7.5 x 101 <32 <1 6.0x 103 2.6 3.6x 103
NDV.ts43 2.1 3.0x 102 <32 <1 3.0x 103 10 3.0x 102
NDVpi
Cl.20 1.0 7.0 120 <ld >106 dNDVp0
ts+20R3 0.4 5.0 x 102 <32 10 5x 102 8.0 5.6x 102NDVp1
ts+38R1 41 7.5 <32 100 40 5.8 7.8x 102NDVp,ts+38R4 110 6.0 64 64 2.5x 102 290 2.4x102
NDVp0ts+38R6 90 8.0 <32 1700 4.5 490 2.1X 102
aInfective virusinculture fluid(7 ml) assayedinCE cellmonolayers.
bInfected cellswerescrapedfromtheglassinto 2mlofMEM,counted inahemocytometer,andplatedfor infectivecenters onCE cell monolayers.
cInterferon inculture fluidsatselected passageswasassayedbytheplaquereductontechnique;valuesare
U/3ml.
dCultures concludedtobenolongerinfected and werediscardedafter
P3.
on November 10, 2019 by guest
http://jvi.asm.org/
PREBLE ANDYOUNGNER lished data). Although attempts to establish
persistent infection with
NDVpi
clones 25to 38werenotincluded in theseexperiments, one can assumethat theywould have behaved similarly
to clone 20. Subtle differences between these clonesand clone20exist (10)but the replication
patterns ofall
NDVpi
clones inL cellsare verysimilar.
As aresult ofpersistent infection withNDV,
LNDV cultures have been shown to become resistant tosuperinfection with unrelated
cyto-lytic viruses, such as VSV (3, 7, 14, 23). The
putative LNDV cultures and uninfected control L-cellcultures werechallenged atP8with VSV, NDVQ, or
NDVpi
clone 35, as described inMaterials and Methods. Plaques of NDV chal-lenge virus were produced 1 to 2 days earlier than NDV plaques caused by the endogenous, carried virus in the
LNDV
cultures.Theyield of challenge virus from each persist-ently infected culture is shown in Table 7. Clearly, VSV replicated only in the uninfected
L cells; the yield of VSV from anyof the LNDV
cultureswasat least 4logs lowerthan the yield ofVSV from the control L-cell cultures. Simi-larly, no virus replication was observed when
LNDVcultureswerechallengedwitheitherNDVO
or
NDVpi
clone 35. The interferon titers in allculture fluids harvested from P8 at the time of superinfection were less than 16 U per 3ml.
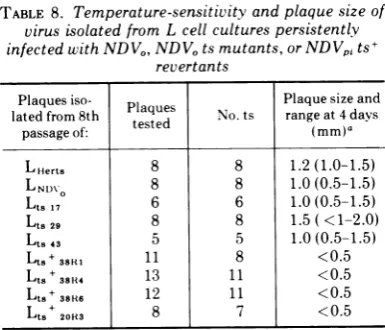
The data given in Tables 6 and 7 clearly demonstrate that the L-cell cultures were
per-sistently infected with NDV. It was also
ob-served during the course of the cell passages
described above that the time required for development of NDVplaques in infectivecenter
assays inCE cell cultures increased from 3 to4 daysto 6days; this suggested that selection of slower growing variants or mutants had
oc-curred. To test this possibility, and to deter-minethe temperature-sensitivity ofvirus
pres-ent, clones of virus were reisolated from each
persistently infected culture and screened for ability to replicate at 37 and43 Cby usingthe
"welled-tray" technique described in Materials and Methods. At 37 C, all infected monolayers showed extensive cytopathology by 48 h. In contrast, in one experiment, only 6 out of 65
monolayers infectedwithvirus clones from LNDV
cultures showed minor cytopathic changes at 43C, although allNDVO-infectedmonolayersat this temperature were completely destroyed.
Culture fluids from about 40% of the wells at 43 Cwereassayedforinfectivityandsignificant virus replication was detected only in fluids
from wells with degenerative changes in the cells.
Intwoexperiments,78plaque-purifiedclones from the various persistently infected cultures
TABLE 7. Yield of challenge virusfrompersistently
infectedL cells (P8) superinfectedwith VSV,NDVO,
orNDVp,clone35
Yieldofchallengevirus(PFU/ml x 103)
Lcells
NDV5i
VSV NDV, clone 35
Controla 5.0x 104 51 170
LHerts 0.7 2.7 1.0
LNDV 1.1 1.8 3.0
Lts
170
1.0 2.3 1.3Lts29 3.9 2.6 3.2
Lts43 1.6 1.3 3.1
Lts+
38Rt1
0.4 4.2 2.6Lts+ 381t4 <0.1 4.6 2.7
Lts+20tR3 0.5 3.2 4.8
[image:8.495.253.447.105.240.2]aUninfected control L cells with the same pass-agehistory asthepersistentlyinfected cultures. TABLE 8. Temperature-sensitivity and plaque size of
virus isolatedfromL cell culturespersistently
infectedwithNDVO,NDVOtsmutants,orNDVpits+ revertants
Plaquesiso- Plaques Plaquesizeand latedfrom8th tested No. ts range at4days
passage of: (mm)a
LHerts 8 8 1.2(1.0-1.5)
LNI)\ 8 8 1.0(0.5-1.5)
Lts17 6 6 1.0(0.5-1.5)
Lts29 8 8 1.5( <1-2.0)
Lt843 5 5 1.0(0.5-1.5)
Lt.+38RI 11 8 <0.5
Lts+38R4 13 11 <0.5
Lt.+38R6 12 11 <0.5
Lt.+20R3 8 7 <0.5
aNDVocontrol plaques were2to 3 mmat 3days,
whereas NDVpi clone35 control plaques were 1 to 2 mm at 4 days.
were screened for
temperature-sensitivity
and virus progeny harvested fromwelled-trays
at 37 C was also tested forplaque
size at 4days
in CE cells at 37 C, to determine if the virus reisolated at P8 was ofa different
plaque
mor-phology from the virususedtoinfect P0.Bythe
eighth cell passage, viruscarried inthe
persist-ently infected L cells was ts no matter which type ofvirus was used for infection
(Table 8).
Also, plaque size of all reisolated virus clones was decreased regardless of whether
large
orsmall plaque virus was used to initiate the persistent infection.
Persistent infection of L cells with other NDV strains. To test whether the selectionof ts virus was
peculiar
to the Herts NDV-L-cell system, twoother strains ofNDV wereused toinitiate persistent infection. L-cell
monolayers
were infected with the Texas-GB
(Tex)
or the 488on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.495.253.446.271.436.2]TSMUTANTS IN PERSISTENTINFECTION
Kansas-Man (Kan) strains of NDV, by using conditions for establishing persistent infection (see Materials and Methods). Wild-type clones
(Texo
andKan.)
and clones from persist-entlyinfected cultures(Texpi
andKanpi)
wereisolated andcharacterized.
The properties of Tex and Kan clones are
summarized in Table 9. The plaque size in CE cells of both
Texpi
andKanpi
cloneswassmallerthan that of the respective wild-types. The frequency of stable ts
Texpi
clones was muchhigher than the frequency of stable ts
Kanpi
clones. This pattern was true whether clones
were isolated early (after4-19cell passages) or
late (after 37-46 cell passages) after
establish-ment of the persistent infections and may
reflect differences between the Tex and Kan strains. Many
Kanp,
clones provedtobeunsta-ble after egg passage (Table 9) and
conse-quently high-titered stocks of those cloneswere
notavailable for use in RNA synthesis
experi-ments.All stabletsclonestested, however, were
RNA- at the nonpermissive temperature. All clones isolated from persistently infected cul-tures alsoreplicated moreefficiently in L cells
than did the wild-type Tex or Kan clones. In
contrasttoresults with Herts
NDVpi,
however,Texpi
andKanpi
virus clones isolated from persistently infected L cells were no differentfrom wild-type
Texo
andKano
clones in the thermal inactivation profiles of infectivity, he-magglutinin, and neuraminidase.DISCUSSION
The objectives of this investigation were to
determine the relationship between the ts
marker and the othercharacteristics of
NDVpi,
and to assess whether selection of such ts mutantsplayedarole ineitherestablishmentor
maintenance of persistent infection of L cells
with NDV. The experiments with
NDVpi
ts+ revertants, NDVo ts mutants induced with HNO2, and spontaneous ts mutants indicate clearly that thets marker can be either lostoracquiredwithoutnecessarilyaltering the physi-cal properties or the growth rate of the virus.
NDVpi
ts+clones retained thesmall plaque size and thermal stability profile ofNDVpi,
while the NDVOts mutantsremained"NDVo"
except for the ability to replicate at 43 C. Also, with spontaneous ts mutants of the population of uncloned Herts NDV, no correlation existedbetweenthetsmarkerandanyof theproperties
characteristic of
NDVp0;
all four clones dis-played the physical andbiological properties of NDVO. However, in L cellspersistently infected with NDV, temperature-sensitivity, small plaque size, and, at least with the Herts strain of virus, altered thermal stability wereco-selected.
Otherexperiments showedthat theabilityto replicate efficiently in L cells appeared to de-pendon thecombination ofthe ts marker with
the other properties of
NDVpi;
bothNDVpi
ts+ revertants and NDVo ts mutants produced acovert infectionin L cells. In addition,
NDVpi,
the only NDV type able to replicate produc-tively in Lcells,wasunabletoestablish persist-ent infection, whereas uncloned Herts virus, NDVO,
NDVpi
ts+ revertants, and NDVO ts mutantswereallabletoestablish and maintain carrier states in L cells. Virus reisolated at P8 from any of these persistent infections was ts andproduced small plaques in CE cellsat37 C. These results strongly suggestthat selection of virusts mutantsis notrandom andmayplayarole in maintenanceof thepersistent infection. Temperature-sensitive mutants of many
other enveloped RNA viruses, including
spon-taneous ts mutants (4), have beenstudied, and
TABLE9. Propertiesofwild-type Texas-GB and Kansas-Man NDV clones andofclones isolatedfromL cells persistently infected with these strains
Passagenumberin No. clonestsb/no. RNAsynthesisat Replicationin Clones l Lcells Plaque size (mm) clones tested 43 C Lcells
Tex, None 4-5 0/4 + Covert
Kan. None 4-5 0/4 + Covert
Texp0 4-19 2-4 14/18(14/14)c _d Productive
Texpi 43-46 2-4 20/24(9/10) - Productive
Kanp0
8 2-4 12/12(4/12) - ProductiveKanpi 37 2-4 12/12(4/12) NTe Productive
aPlaquesize inmminCE cellsafter 7daysat37C.
bYieldoforiginalcloneinCE cellsat43C yieldat37C <10-2orplaquetiter of
original
cloneat43C plaquetiterat37 C < 10'.cNumber oftsclones stable duringeggpassage pernumberoftsclones testedineggs; eggsstillalive 48 h
afterinoculationwereharvested.
dRNAphenotypeofstabletsmutantsgrown ineggs. eNT,nottested.
VOL.12,1973
on November 10, 2019 by guest
http://jvi.asm.org/
PREBLE ANDYOUNGNER in each case, characterization of the mutants
with regard to functional defect has revealed both RNA+ and RNA- mutants (1, 12, 22). Although other workers (2, 8) reported isolation of ts mutants of NDV, no biochemical analysis of the mutants has been described. Results presented here suggest that ts mutants of NDV, whether spontaneously occurring, isolated after mutagen treatment, or selected by persistent infection, are always RNA- (10). Several dozen ts mutants of NDV have been tested in this laboratory and all of the mutants consistently failed to synthesize any detectable virus-specific RNA atthe nonpermissive temperature under conditions where wild-type virus pro-duced large amounts of RNA. The functional defect in NDVpi ts mutants appears to be associated with RNA polymerase (11). The other ts mutants discussed in this report have not yet been analyzed in as much depth. The significance of findingonly RNA- ts mutants of NDV is difficult toexplain. The reason maylie in unusual genetic properties associated with NDV or in complicated virion protein interac-tions that allowonly certain limited variations instructure that are notlethal.
The mechanisms for the establishment and perpetuation of L-cell cultures persistently in-fected with NDV are largely unknown. The experiments described above explore the possi-bility that selection ofts virus mutants, which also differ fromwild-type virusinother proper-ties, may be involved in maintenance of the carrierstate.Otherworkers havealso noted that virus recovered from persistently infected cell cultures may be different in properties, e.g., plaque size and virulence, from the wild-type virusused to initiate the infection (5, 17, 18, 21, 26). In several instances reports have also ap-peareddescribingtsbehavior of virus associated with persistent infections. An attenuated mu-tant isolated from Frukto mousesarcoma cells persistently infected with Western equine
en-cephalitis (WEE) virus (18) also had a de-creased growth
potential
at 42C,
whereas the wild-type virus replicated normallyatthis tem-perature (19). When the attenuated strain waspassaged at 41 C and clones were selected for growth atthat temperature, theresulting popu-lation retained its attenuated characteristic. In contrast, when virulent revertants were se-lected, the reversion to virulence hadno effect on the ts marker(19). Thesestudies, therefore,
were similar to the studies with NDVpi ts+ revertants and
NDVO
ts mutantsreported here in which the ts and plaque type markers ofNDVpi
were foundtobeindependent mutations even though they were co-selected in the per-sistent infection. The studies with the WEEmutant support the hypothesis that mutations selected during persistent infection are not random. In another study, Nagataand cowork-ers (9) reported the establishment of BHK-21 cells persistently infected with Sendai virus. The viral maturation in these cultures was normal at 31 C, but ts at 37 C.
Onestudy has also linked carrier infections in vivo with selection of ts mutants (20). Plaque production by strains of foot andmouth disease virus isolated from carrier cattle was inhibited at 40 to 41 C, whereas plaque production by vesicular isolates was not affected by the in-creased temperature. Inaddition, plaque size on BHK monolayers was smaller with carrier iso-lates than with vesicular isoiso-lates.
Results from several different virus-cell sys-tems, therefore, suggest that a property of persistent infection is selection of mutants or variants with impaired ability to replicate at high temperatures. The demonstration of a small population of spontaneous ts mutants in Herts NDV preparations suggests that perhaps these virus clones are selected during early stages ofthe persistent infection of L cells with NDV and only later do the structural altera-tionsimplied bythealtered thermalstability of
NDVpi
evolve. It is possible that a low level ofthis type of virus is present even in cloned populations of virus, such as NDVO or the
NDVpi
ts+ revertants since passage througheggs is known to affect the stability of NDV populations (23; Preble and Youngner, unpub-lished data).
We are currently investigating the generality ofthe phenomena observed in the NDV-L-cell system; we are testing virus-carrier systems involving other paramyxoviruses, as well as viruses unrelated to NDV, in L cells and other cell lines to determine if the outcome of persist-ent infection is dependent on the particular virus or host cell used.
ACKNOWLEDGMENTS
The excellent technical assistance of DianeQuagliana is gratefully acknowledged.
Thisinvestigation wassupported byPublic HealthService
research grantAI-06264andtraining grantAI-00080fromthe
National Institute ofAllergyand Infectious Diseases. It was
conducted under the sponsorship of the Commission on
Influenza, Armed Forces Epidemiological Board, and was supported bythe U.S. Army Medical Research and Develop-ment Command, Department ofthe Army, under research
contract17-67-C7046.
Olivia Preble was a Predoctoral Fellow of the National
Institutes ofHealth underfellowship 5FO1 GM49499 from
the National Institute ofGeneral Medical Sciences.
LITERATURE CITED
1. Burge, B.W., and E. R. Pfefferkorn. 1966. Isolation and
characterization of conditional-lethalmutantsof
Sind-bisvirus. Virol. 30:204-213.
490
on November 10, 2019 by guest
http://jvi.asm.org/
2. Dahlberg, J., and E. H. Simon. 1969.Recombination in Newcastle disease virus (NDV): the problemof
com-plementing heterozygotes. Virol. 38:490-493. 3. Deinhardt, F., V. Bergs, G. Henle, and W. Henle. 1958.
Studiesonpersistentinfections of tissuecultures. III. Somequantitative aspects ofhost cell-virus
interac-tions.J. Exp. Med. 103:573-589.
4. Flamand, A. 1970. Etude genetique du virus de la stomatite vesiculaire: classement demutants thermo-sensible spontanesen groupesdecomplementation. J. Gen. Virol.8:187-195.
5. Gavrilov, V. I., D. M. Asher, S. D.Vyalushkina, L. S. Ratushkina, R. G. Zmieva, and B. G. Tumyan. 1972. Persistent infection of continuous line ofpig kidney cellswithavariantofthe WSN strainof influenzaA.
virus. Proc.Soc.Exp. Biol. Med. 140:109-117. 6. Granoff, A. 1961. Induction of Newcastledisease virus
mutantswithnitrous acid.Virology 13:402-408.
7. Henle, W., G. Henle, F. Deinhardt, and V. Bergs. 1959. Studiesonpersistent infections of tissue cultures. IV.
Evidencefortheproduction ofaninterferon in MCN
cellsbymyxoviruses. J. Exp. Med.110:525-541. 8. Kirvaitis,J., and E. H. Simon. 1965. Aradiobiological
study of the development ofNewcastledisease virus. Virology26:545-553.
9. Nagata, I., Y. Kimura, Y. Ito, and T. Tanaka. 1972. Temperature-sensitive phenomenon of viral
matura-tion observedinBHK cellspersistently infected with HVJ.Virology 49:453-461.
10. Preble, 0.T., and J. S. Youngner. 1972.
Temperature-sensitive mutants isolated from L cells persistently infected with Newcastle disease virus. J. Virol. 9:200-206.
11. Preble, 0.T., and J. S. Youngner. 1973.
Temperature-sensitive defect ofmutants isolated from L cells per-sistently infected with Newcastle disease virus. J. Virol. 12:472-480.
12. Pringle, C. R., and I. B. Duncan. 1971. Preliminary physiological characterization of temperature-sensitive
mutantsofvesicularstomatitis virus. J.Virol.8:56-61.
13. Reeve, P., and G. Poste. 1971.Studiesonthe cytopatho-genicityof Newcastledisease virus: Relation between virulence, polykaryocytosis and plaque size. J. Gen. Virol. 11:17-24.
14. Rodriguez, J. E., andW.Henle. 1965.Studieson
persist-ent infections of tissue cultures. V. Initial stages of infection ofL(MCN) cells by Newcastledisease virus. J.Exp. Med. 119:895-921.
15. Rodriguez, J. E., V. terMeulen, and W. Henle. 1967. Studiesonpersistent infectionsoftissuecultures.VI. Reversible changesinNewcastle diseasevirus popula-tionsas aresult ofpassageinL cellsorchickembryos.
J. Virol. 1:1-9.
16. Schloer, G. M., andR. P. Hanson. 1968. Relationshipof plaquesizeand virulenceforchickensof 14 representa-tiveNewcastle disease virusstrains.J.Virol. 2:40-47. 17. Schwobel, W., and R.Ahl. 1972.Persistenceof Sindbis
virusin BHK-21 cell cultures. Arch. Gesamte
Virus-forsch.38:1-10.
18. Simizu, B., and N. Takayama. 1969. Isolation of two
plaque mutantsof Western equineencephalitis virus differinginvirulence for mice.J. Virol. 4:799-800. 19. Simizu, B., and N. Takayama. 1971. Relationship
be-tweenneurovirulence andtemperaturesensitivityofan
attenuated Western equine encephalitis virus. Arch. GesamteVirusforsch.35:242-250.
20. Straver, P. J., and J. G. van Bekkum. 1972. Plaque production by carrierstrainsoffoot-and-mouth disease
virus inBHK-monolayers incubated atdifferent
tem-peratures.Arch. GesamteVirusforsch.37:12-18.
21. Takemoto, K.K., and K. Habel.1959.Virus-cell relation-shipinacarriercultureofHeLa cells andCoxsackie A9
virus.Virology7:28-44.
22. Tan, K. B., J. F. Sambrook, and A. J. D. Bellet. 1969. Semliki Forest virus temperature-sensitive mutants:
isolation andcharacterization. Virology 38:427-439. 23. Thacore, H. R., and J. S. Youngner. 1969. Cells
persist-ently infected with Newcastle disease virus.I. Proper-ties ofmutants isolated from persistently infected L cells. J.Virol.4:244-251.
24. Thacore, H. R., and J. S. Youngner. 1970.Cells persist-ently infected with Newcastle disease virus. II. Ribonu-cleicacid andprotein synthesisincellsinfected with
mutantsisolated frompersistently infected L cells. J. Virol.6:42-48.
25. Thacore, H. R., and J. S. Youngner.1971.Cells persist-ently infected with Newcastle disease virus. III. Ther-malstability ofhemagglutinin and neuraminidase ofa
mutant isolatedfrom persistently infected L cells. J.
Virol.7:53-58.
26. Walker, D. L.1964.Theviral carrierstateinanimalcell
cultures.Progr. Med..Virol.6:111-148.
27. Youngner, J. S., A. W. Scott, J. V. Hallum, and W. R. Stinebring. 1966. Interferon production by inactivated Newcastle disease virusincell cultures andin mice.J. Bacteriol.92:862-868.
VOL. 12,1973