JOURNAL OF VIROLOGY, Nov. 1976, P.425-435
Copyright0 1976 American SocietyforMicrobiology Printed inVol.20,U.S.A.No.2
Multiple
Methylated Cap Sequences
in
Adenovirus
Type
2
Early mRNA
SHU-ICHI HASHIMOTO AND MAURICE GREEN*
Institutefor Molecular Virology, Saint Louis University School ofMedicine, St. Louis, Missouri 63110 Received for publication 26 April 1976
The methylated constituents of earlyadenovirus2mRNA werestudied. RNA was isolated from polyribosomes of cells double labeled with
[methyl-3H]methionineand32pO4 from 2 to 7 h postinfectioninthe presence of
cyclohexi-mide.Cycloheximideensuresthatmethylation and processingareperformed by
preexisting host cell enzymes. RNA was fractionated into polyadenylic
[poly(A)I+ and poly(A)- molecules using poly(U)-Sepharose, and undegraded
virus-specific RNA wasisolatedbyhybridizationtoviral DNA in50% formam-ide at3700.Viral mRNA wasdigestedwithRNase T2andchromatographedon
DEAE-Sephadex in7 M urea. Two3H-labeled RNase T2-resistant
oligonucleo-tidefractionswithcharges between -5 and -6wereobtained,consistentwith twoclasses of5'terminalmethyl "cap"structures,
m7G(5')ppp(5')NmpNp
(cap 1)andm7G(5')ppp(5')NmNmpNp (cap 2) (Nmisaribose2'-O-methylation).The
puta-tive cap 1 contains m7G, Am, Gm, Um, and m6Am. Cap 2 contains all the methylated constituents of cap 1 plus Cm. The molar ratios ofm7G to 2'-O-methylnucleosides is about 1.0 for cap 1 and 0.5 for cap2, consistent with the
proposedcapstructures. Mostsignificant,compositional analysis indicatesfour
different cap 1 structures and at least three different cap 2 structures. Thus
there is a minimum of seven early viral mRNA species with different cap structures, unless each type of mRNAcanhavemorethanone5' terminus.In addition tomethylated caps,earlymRNA containsinternal basemethylations,
exclusively asm6A, asshown byanalyses of the mononucleotide (-2 charge)
fraction. m6A was presentinthe ratio of1mol ofm6Apper450nucleotides.Thus viral mRNA molecules contain two to three internal m6A residues permethyl cap, since there is on the average 1 cap per 1,250 nucleotides.
Thetranscription of DNA tumor virus genes
in infected and transformed cells provides a
model for analyzing mRNA biogenesis and
posttranscriptional regulation in eukaryotic
cells. The human adenoviruses, in particular
the well studied adenovirus2 (Ad2), are
espe-ciallyuseful for such studies. Ad2productively
infectscultured human cells (KBorHeLa)and expresses its genomein twostages,"early" (be-fore viral DNA synthesis at 6 to 7 h postinfec-tion) and "late" (14, 32). Early Ad2 mRNA is transcribed in the cell nucleus and processed in the absence of protein synthesis, so host cell enzymes must perform these functions (al-thoughthepossibilityof a virion methylaseor
RNA polymerase has not been excluded). Therefore, by studying early Ad2 mRNA syn-thesis we can learn about the cellular mecha-nisms that regulate both viral and cell gene expression.
Inthisconnection, the possible role of meth-ylationismostintriguing. It has recently been
demonstrated that mRNA from animal cells
(26) and viruses that replicate in eukaryotic cells (10) ismethylated. With RNA viruses that
multiply in the cytoplasm, methylation is
ex-clusively at the 5' terminus in two classes of
"capped" oligonucleotides, m7G(5')ppp(5')Nm
andm7G(5')ppp(5')NmpNm (Nm is a ribose
2'-O-methylation), respectively, termed cap 1 and
cap 2. The basepenultimate tom7G is Am (or
m6Am) in vesicular stomatitis virus mRNA (in vitroand in vivo) (1, 22, 29), and cytoplasmic
polyhedrosis virus mRNA (in vitro) (13),
whereas it isGm in reovirus mRNA (in vitro) (11). ThemRNA generated invitroby cores of vacciniavirus, acytoplasmic virus with a dou-ble-stranded DNA genome, contains two caps withpenultimate Am and Gm (34). All of these
invitrosynthesized viral mRNA's but vesicular stomatitis virus contain only cap 1 structures. The methylation of mRNA synthesized dur-ing late stages of infection by twoDNA tumor viruses thatreplicate in the cell nucleus, sim-ian virus 40 (SV40) (3, 18) and Ad2 (9, 21, 36), have been studied. Presumptive evidence for
425
on November 10, 2019 by guest
http://jvi.asm.org/
426 HASHIMOTO AND GREEN
SV40 mRNAcap1structureswithpenultimate
Am and Gmwasreported (18). LateAd2mRNA
hasbothcap1andcap2 structures (21,36). The cap 1 fraction was found to consist of 80%
m7Gpppm6Am and20%m7GpppAm (21);the cap 2 fraction wasnot fully characterized and
ap-pears tobe morecomplex. Unlike cytoplasmic
viruses,Ad2 (21) andSV40 (18)mRNA, aswell
ascellRNA(33),containinternal methylations,
exclusively m6A. In contrast to the restricted composition ofviralcaps,cellmRNAcontainsa
wide varietyofmethylcaps,ofbothtype 1and type 2: 27 different caps were isolated from
mRNA ofmousemyelomacells (6), and
multi-plecaps werefoundinmRNA ofHeLacells(12,
33) andmouseLcells (27).
Severalfunctions of the 5' terminal
methyla-tion of cell and viral mRNA have been
sug-gested, including: (i) ribosome binding for the
translation of mRNA (4, 23), (ii) recognition signals for processingof nuclear precursorsto mRNA (30), and(iii)protection against
exonu-cleases. Conceivably, different cap sequences
affect translation control, or reflect different
recognition signals for putative cleavage en-zymes or initiatingsequences forRNA polym-erase. Unfortunately, the large number of
mRNAspecies ineukaryotic cellslimits
experi-mentalapproachestofunctional analysisofcap structures.
The study ofAd2early mRNA methylation
offersanopportunitytoinvestigate thepossible
functions of different caps. Unlike late viral
mRNA methylation which may involve the
functioningof viralgeneproducts,early mRNA
methylation probably involves only host cell
enzymes. The genes encoding early mRNA have been mapped (28, 31), individual mRNA
sequencesfromeachgenehavebeenidentified (5, 7, 8; W. Buttner, Z. Veres-Molnar, and M. Green, J. Mol. Biol., in press), and
proce-dures have beendevelopedtopreparatively
iso-late discrete viral mRNA species (Buttner et
al., J. Mol. Biol., in press; 5a). Early Ad2 mRNAhas beentranslatedinvitro into at least sixpolypeptideswhosegeneshavebeen located (19). From six (Chin and Maizel, in press;
14a) to as many as 11 (W. S. M. Wold, M.
Green, and W. Buttner, in press) early
pro-teins (notnecessarilyviralcoded)have been
de-tected in vivo. Thus it is possible to correlate
the cap structure of each early mRNA with
their position on the viral genome and the
synthesis ofspecific viral proteins. Such
infor-mation can help elucidate possible functional
roles forindividualcaps. Wehaveanalyzedthe methylated components of early Ad2 mRNA, and find that it is methylated internally as
m6A,as areothermRNAmoleculestranscribed
in thenucleus. But unlike previously reported viral mRNA species, Ad2 early mRNA has a variety of different methyl caps, both type 1
and type 2. Compositional analysis indicates four type 1 caps andatleastthreetype2 caps. Both types of caps contain m6fAm. These data
are consistent with the possibility that each Ad2 mRNA species has a different cap struc-ture.
MATERIALS AND METHODS
Cell culture and virus infection. Exponentially
growing KB cells in suspension were cultured in
Eagle minimumessential medium (MEM) contain-ing 5%horseserum. Cells wereconcentratedto 3 x
106 to 5 x 106cells/ml by briefcentrifugation and infected in MEM with 100 PFU/cell of Ad2 (plaque 4, strain 38-2, free of adenovirus-associated viruses).
After 1 h ofadsorption, cells were dilutedwith 10
volumes of MEM (with horse serum) containing 25 ,tgofcycloheximideper ml. At2hpostinfectioncells were concentrated 10-fold, resuspended in phos-phate-free MEM containing one-tenth the concen-trationofmethionine and 25 j,g ofcycloheximideper ml, and labeled from 2 to 7 hpostinfection with 1 mCi Of 32P04 per ml and 200 ,uCi of [methyl-3H]methionineperml (13Ci/mmol, Schwarz/Mann). Cell fractionation and isolation of Ad2-specific RNA. RNA was isolated from thepolyribosomesof infected cells aspreviously described (5). Polyriboso-mal RNA was fractionated on polyuridylic acid
[poly(U)]-Sepharosecolumns intopolyadenylic acid
[poly(A)]+ andpoly(A)- RNA fractions (20) and
an-nealed in 50% formamide at 37°C with Ad2 DNA immobilized on membrane filters (5). In some exper-iments, hybridization-purified RNA was further
purified by rehybridization to Ad2 DNA. Poly(A)+
[3H]RNA used incontrol experiments was isolated
from polyribosomes ofuninfected KB cells labeled for15 hinthe presence of 20 ,uCi of[3H]uridine per ml (39 Ci/mmol, New England Nuclear). In addi-tional control experiments, [methyl-3H]-labeled
poly(A)+ RNA was isolated from polyribosomes of uninfected KB cells labeled for 12 h in the presence of 50 ,uCi of [methyl-3H]methionine (13 Ci/mmol, New England Nuclear). Ribosomal RNA used as markers for gelelectrophoresis wasisolated as de-scribed by Hashimoto andMuramatsu (15).
Polyacrylamide gel electrophoresis of viral mRNA.Hybridization-purifiedviralmRNA was an-alyzed by electrophoresis on gels (0.6 by 20 cm) containing 2.5%polyacrylamide, 0.5%agarose, and 0.2% sodium-N-lauroylsarcosinate (ICN Pharma-ceuticals) (5).Electrophoresis wasperformed for6 h
withconstantvoltage (150 V) at 5C with
recircula-tion of buffer. Ribosomal [3H]RNA markers were
coelectrophoresed with Ad2mRNA. Gels were frac-tionated into 2-mmportions with a Gilson gel frac-tionator (Gilson Medical Electronic Co.) and countedinAquasol (New England Nuclear) usinga
scintillationcounter.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
CAP SEQUENCES IN Ad2 EARLY mRNA 427 RNaseT2digestion andDEAE-Sephadex column
chromatography. 32P_ and [methyl-3H]-labeled vi-rus-specific RNA togetherwith 0.5 to 1 mgof carrier
yeast tRNA weredigestedwith 2unitsofRNase T2 (Sankyo Co., Ltd)irn0.5 mlof 0.05Msodium acetate (pH4.5)at37°C for15 h (15).One unitofRNase T2
was added, and incubation was continued for an additional 5 h. The digest was treated with 0.1 N HCl at 0°C for 5 h to cleave cyclic phosphodiester linkages. After neutralization, the solutionwas di-luted with5ml of 7 Murea-0.03 M Tris-hydrochlo-ride (pH 7.6) andloadedontoaDEAE-Sephadex
A-25column (0.4by30 cm).Fiveoptical density units
at 260 nm
(OD2N)
ofa pancreatic RNase digest ofyeast tRNA was added in the application buffer. Elutionwasperformed withalinear gradient of0to 0.4MNaCl(70mlof each)inapplicationbuffer. In
several experiments, elution with 200 ml of eluant
(100ml each) was utilized (16). TheOD2,0 ofeach fraction (1.4ml) was measured, and 100to 200Ml
wasanalyzedfor32p and3H counts perminute.
Compositional analysis of methylated cap struc-turesisolated from Ad2 mRNA. Cap1 (-5charge) and cap2(-6charge) fractions resolved by
DEAE-Sephadexchromatography were pooled, desalted by elution from DEAE-Sephadex (0.3 by2.5cm)with2 Mtriethylamine bicarbonate, dried, anddissolved
in 20 Mul of 50 mM Tris-hydrochloride (pH 8.5) -5
mMMgCl2 containing nucleoside markers (Am, Um, Cm, and m7G, approximately 1OD2. unitofeach). The mixturewasincubatedat37°C for4 hwith5 Mg each of venom phosphodiesterase andEscherichia
coli alkalinephosphatase (Worthington Corp.). The mixture wasreincubated at37°C for 1h after the addition ofanadditional5 Mgofeachenzyme.
Nu-cleosides were separated by ascending paper chro-matographyonWhatman no. 3MM paper strips (1.5 by46 cm) using 1-butanol-0.8 Mboric acid-concen-trated ammonia (2,000:270:8, vol/vol/vol) (2). The chromatogramwasdevelopedat roomtemperature
until the solvent had ascended 30 to 35 cm. The
paper wasdried, cut into 0.5-cmstrips, and eluted with 0.5 mlof0.01 NHClat37°Cfor 15 h.The four markerpeaks which were detected and identified by absorbance at 260 nm and UV spectrum at pH 2migrated inthe order Am, Um, Cm, m7G. The absorbanceat260 nmshowedthatelutionof
nucleo-sideswasquantitative. Gmwasidentifiedfrom
par-allelchromatograms of phosphodiesterase, alkaline phosphatase digests of methyl-3H-labeled RNase
T2stable dinucleotides, isolated fromKBcells ribo-somalRNA(16). Samples of each
eluan%t
(100 to 200 Ml)weredissolved in5mlofBiofluor(NewEnglandNuclear) andcounted in ascintillation spectropho-tometer. In some experiments (after samples for
counting were taken), fractions ofm7Gand m6Am peaks werepooled and analyzedon Dowex-50 col-umnsasdescribed below.
Dowex-1 andDowex-50chromatography.
Analy-sisofmononucleotideand capfractionsand
identi-fication ofm7G, m6A, and m6Am. The
mononucleo-tidefractions(-2charge)obtained by
DEAE-Sepha-dex column chromatography were pooled, diluted fivefoldwithwater, and loaded on a Dowex-1 column
(0.3 by 20 cm). The column wasequilibrated with
0.004NHClanddeveloped withaconcavegradient
formedwitha40-mlmixingchamber(0.004 NHCl) and a 20-ml reservoir(0.1MNaCl and0.01NHCl) (15). Constituentseluted in theorderCp, Ap,m6Ap,
inorganic phosphate, Up, and Gp. One-milliliter fractions werecollected and suitablesampleswere
counted.m6Ap-containingfractionswerepooled, re-chromatographed on Dowex-1, depurinated, and chromatographed onDowex-50 asdescribed below. Further identification of m7Gandm6Am fromcaps
and m6Ap fromthe -2fraction was performedby
Dowex-50 column chromatography using a micro modificationoftheprocedure of IwanamiandBrown (17). The nucleosideornucleotide fractionwas
depu-rinated by treatment with 1 N HCl for 60 min at
100°C. The sample was diluted to 0.02 NHCl, ap-pliedonto aDowex-50 column(0.2by20cm), washed
with 0.02 NHCl, and eluted with0.5NHCl. After 20 fractions (0.6ml each) were collected, elutionwas
performed by 0.7 N HCl. The elution sequence of
purineswasGua, m7Gua, m'Ade, Ade, m6Ade. RESULTS
Isolation from polyribosomes of poly(A) terminated early Ad2 mRNA double labeled
with[methyl-3H]methionineand32PO4.To
ob-tain early Ad2 mRNA for analysis of methyl-atedconstituents, cellswerelabeled withhigh concentrations of P2PO4 (1mCi/ml) and
[methyl-3H]methionine (200
ACi/ml)
from 2to7hpost-infection in thepresenceof cycloheximide. Cy-cloheximide was usedbecause it increases the yieldof early viral mRNA(5, 25);it alsoblocks late viral mRNA synthesis, and prevents the synthesis of earlyviral-codedproteins, thus as-suring that host mechanisms were utilized to
process (methylate) early viral RNA tran-scripts (unless there is a virion methylase). From2 to 3 mgofpolyribosomal RNA with 32P specific activity of about 2 x 105 cpm/,ug was
isolated from1liter of infected KB cells (3 x 105
to 5 x 105 cells/ml). Polyribosomal RNA was fractionated into poly(A)+ and poly(A)-
mate-rial using poly(U)-Sepharose; analysis offive preparations showed that about 40% of
[32P]RNA waspoly(A)+ and 60%waspoly(A)-.
Viral mRNA was selected byhybridization to
Ad2DNA in 50%formamideat37°C.In a
repre-sentativeanalysis (Table 1), 15.6% ofpoly(A)+ RNA and 2.0%ofpoly(A)- RNAformedhybrids. In control experiments, generally less than
0.1%ofpoly(A)+RNAisolated fromuninfected
cells hybridized to Ad2 DNA (e.g., Table 1). This indicates that contamination of early poly(A)+ viral mRNA with cell mRNA after
hybridizationshould be less than 1%.
The integrity ofhybridization-purified viral
RNA wasroutinelycheckedby polyacrylamide gel electrophoresis. The typical gel pattern shown inFig. 1Aissimilar to thatreported pre-VOL. 20, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
428 HASHIMOTO AND GREEN J uo~
TABLE 1. Sekectionofpoly(A)+andpoly(A) Ad2 mRNAby hybridization toviral DNA
RNAhybridized
LabeledRNA Input (cpm) -__________
cpm
poly(A)+ (32P, [methyl_3H])a Ad2-infectedKBcells 9,644,000(32p) 1,515,000(32P) 15.6
poly(A)- (uP, [methy1_3H])a Ad2-infectedKBcells 73,670,000(32P) 1,529,000(32p) 2.0
poly(A)+ ([IHluridine)buninfected KBcells 5,516,000(3H) 3,000 (3H) 0.05
poly(A)+ ([methyl_3H])c uninfectedKB cells 43,000 (3H) 50 (3H) 0.11
a PolyribosomalRNA (2to3mg)isolated fromKBcells labeledwith [methyl_3Hlmethionine and
32P04
from 2 to 7 h postinfection with Ad2 was fractionated into poly(A)+ and poly(A)- RNA on a
poly(U)-Sepharosecolumn.One-tenth ofpoly(A)+RNAand one-half ofpoly(A)-RNAwereannealedtoAd2 DNA(50
.tig)
immobilizedonnitrocellulosefiltersin 1ml ofhybridizationbufferat370Cfor15h.bPolyribosomal RNA (2 to 3 mg) was isolated fromuninfected KB cells labeled with [3H]uridine and
fractionatedintopoly(A)+ andpoly(A)- RNA. One tenth of thepoly(A)+RNAwas annealed with50 /igof Ad2DNA asdescribedabove.
PolyribosomalRNA(2to3mg) wasisolatedfromuninfected KBcells labeledwith [methyl-3 Hlmethio-nine and fractionatedintopoly(A)+andpoly(A)- RNA. One-tenth of thepoly(A)+RNAwasannealedwith 50 .tLgofAd2DNAasdescribed above.
20 13S,andtwo
11S);
Buittner
etal. (J.Mol. Biol.,A in press) have preparatively isolated and
15
2Rs
18s 5smapped
five major early mRNA species (24S,I
~~~~~~~20.5S,
two 20S, andoneortwo14S).
10
Only
about 20% of total viral RNA waspoly(A)-
(Table 1).Rechromatography
on5
~~~~~~~~~poly(U)-Sepharose
showed that about 40% of5V
hybridization-purified
poly(A)-
viral RNA4
~~~~~~~~~~~~eluted
inthepoly(A)+
fraction; therefore, 10 to~~0 I
~~~~~20%
ofpoly(A)+
viralRNAwasnotretainedonB the first
poly(U)-Sepharose
column. Thegel
i~80
8 8 spattern
ofpoly(A)-
early
viralmiRNA
included28s18s 5s
~~~the
major
20-21S peak
(Fig.iB)
anddecreased60 levels of smaller viral mRNA
species.
The40-
methylated
components ofpoly(A)-
mRNAwere
analyzed
inparallel
withpoly(A)+
viral20
-MRNA,
as describedbelow.Resolution of methylated components
re-0 leased from Ad2mRNAbyRNase T2.To
char-0 20 40 60 80 acterize the
methylated
nucleotides ofearly
Slice number Ad2 mRNA, RNase T2
digests
wereprepared
FIG. 1.
Polyacrylamide
gel electrophoresis of and fractionated onDEAE-Sephadex.
RNasepoly(A)+(A) and poly(AP-(B) Ad2-specific early T2
hydrolyzes
phosphodiesterbondsthrough
a mRNA. Early polyribosomal RNA labeled with 2',3'-cyclicintermediateso thatphosphodiester 32P04and [methyl-3H]methionine was fr-actionated bonds in caps (pyrophosphate) and thoselink-intopoly(A)+andpoly(A)-RNAbychromatography ingthe 3' hydroxylof
internal
2'-O-methylatedon poly(U)-Sepharose, and each fraction was an- nucleotides are stable. Chromatography on nealed to membrane filters containing Ad2 DNA. DEAE-Sephadex in 7 M urea at neutral pH Viral RNA waselutedfr-om thehybridand
electro-reovslinuetdsonhebisfcag.
phoresed on polyacrylamide agarose gels togetherFgresolvsholignuletides
onAtehaexbss
chroarge.
with KB cell ribosomal
[PH]RNA
marker. Fgr hw h EESpae hoaograms of T2
digests
prepared froM 32P04 PlUSviously (8, 20, 25). Several small peaks were [
methyl-_3H]methionine
-labeled poly(A)-se-reproducibly
found between the major 20-215 lected Ad2nmRNA,
once (Fig. 2A) and twicepeak and 13-148 RNA, and a broad shoulder (Fig. 2B)
hybridization
purified.
Over 98% ofwasfoundbetween 215 and28S. This
complex
the32Pradioactivityapplied
tothecolumn waspattern includes at least five to seven viral recovered, and no further radioactivity was
mRNA molecules. Craiget al. (7, 8) have de- eluted with 1 M NaCi after completion of the tected atleastsevenearlymRNAspecies byan gradient. The mononucleotide
peak
(-2analytical
mappingprocedure (209,
19S, three charge)contained 99.6% of the total32P-labeled J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
CAP SEQUENCES IN Ad2 EARLY mRNA 429
0.1
c
0
(.0
C\M
.-O
w
0 z
m
a:
0
C/)
m
0.1
-U
I I 0K
0
0.2
-2
E
0
0.1 --Ia
3
2
CM I
x
6 4
2
20
40
60
80
TUBE
NUMBER
FIG. 2. Chromatography of RNase T2digest of 32p-,[methyl-3H]labeled Ad2 early mRNAon a
DEAE-Sephadexcolumnin7Murea atpH 7.6. Each enzymedigestwasco-chromatographedwith5OD260 unitsof
pancreaticRNase digest of yeast tRNA toprovideoligonucleotide markers as described in Materials and
Methods.3H cpm due to 32p spillover counts (about2%o)inmononucleotides fraction were notsubtracted.(A) Poly(A)+RNA (once hybridized); (B) poly(A)+RNA(twicehybridized); (C) poly(A)-RNA (oncehybridized).
material eluted from digests ofAd2 poly(A)+ mRNA, the dinucleotide (-3 charge), and
tri-nucleotide (-4 charge) peaks contained no
counts,andoligonucleotide peakswith
approxi-mate charges -5 and -6, respectively,
con-tained about 0.20%and0.25%ofthe 32p radioac-tivity (Fig. 2Aand 2B). From furtheranalysis
ofthe -5and-6chargedfractions(seebelow),
andfrom the positions of elution from the col-umn, wetentativelyconclude that the -5and
-6 fractions, respectively, represent5' termi-nal structures of the general type m7GpppNmpNp (cap 1) andm7GpppNmpNmpNp (cap 2), derived from Ad2 early mRNA. We
calculate 1 cap per 1,250nucleotides, basedon
the ratio ofthe radioactivityin32P-labeled caps
(-5 plus -6 fractions) to that of total Ad2 [32P]mRNA.
Analysis of Ad2 poly(A)- early mRNA (Fig.
2C) showed a slightly higher content of
frac-tions with aproximate charges of -5 (0.25%) and -6 (0.38%),probably because the
poly(A)-fraction containedsomeviral mRNAlacking 3'
sequences, perhaps duetodegradation. About
0.1%of the32Pradioactivity was presentinthe dinucleotide fraction and coeluted with a peak
of 3H radioactivity. This peak was probably
derived from contaminating rRNA; e.g., 3% contaminationwith rRNAwould produce 0.1% RNase-resistantdinucleotides (16).
Methylated constituents of cap1and cap2
fractions in Ad2 mRNA. The 3H-labeled
nu-A
~~-2
A
-4 -5 -6 -7-8B
40
60_2
80
1000
60 --2
-3 -4 -5 -6-7-8
- ~
VOL. 20, 1976
0.24-0.2
-4
I20 40 80
11
II
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.102.393.67.436.2]430 HASHIMOTO AND GREEN
cleoside componentsofcap 1 andcap 2isolated from hybridization-purified Ad2 mRNA were
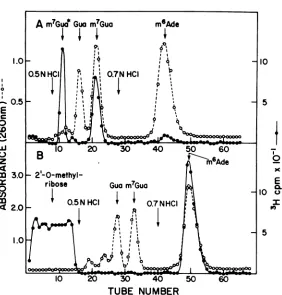
analyzed. Each cap fraction wasdigested with bacterial alkaline phosphatase and venom phosphodiesterase, and the resulting 3H-meth-ylatednucleosides were resolved by paper chro-matography.Figures3A-Dshow the chromato-grams of poly(A)+ cap 1, poly(A)+ cap 2,
poly(A)- cap 1, and poly(A)- cap 2,
respec-tively. Peaks 1, 4, 5, 6, and 7 are m7G, Gm, Cm, Um,and Am, respectively, as shown by co-chro-matography with marker nucleosides. Peak 1 waseluted from paper chromatograms and also identified as m7G by depurination and chroma-tography on Dowex-50. The elution position wasidentical to m7Gua prepared by depurina-tionof authentic m7G (Fig. 4A). Usually some
m7Gua in isolated caps was partiallyconverted
to the ring-opened form (m7Gua*, 2-amino-4-
hydroxyl-5-(N-methyl)formamide-6-ribosylam-ino-pyrimidine). This occurs during isolation
andanalysis ofcapsunderslightlyalkalinepH
conditions (pH 8 to 9). m7Gua* was identified
fromthe elution profile ofm7Gua* produced by
alkalinetreatment(1 NNH4OH,37°C,6h)and depurination ofpm7G. m7Gua*waselutedfrom Dowex-50 infraction2to4 after application of 0.5 N HCI. m7Gua*wasobtainedby depurina-tion of alkaline treated mRNA, whereas only m7Gua was obtained from untreated mRNA.
Peak 8 was expected to be a 2'--methylated adenosine derivative, becauseonly 2'-O-meth-ylated nucleosides (or 2'-deoxynucleosides)
would move with this solvent system and N6-methyl-2-deoxyadenosine moves the furthest among2'--blocked nucleosides (2). To charac-terize peak 8, the chromatogram spot was
eluted, depurinated, and chromatographed on
Dowex-50 (Fig. 4b). Equimolar amounts of 3H-labeledm6Ade and 3H-labeled noncharged
ma-terial, 2'-O-methylribosewerefound, asshown by co-chromatography with authentic m6fAde. Therefore, peak 8 is N6, 2'-O-dimethyladeno-sine. Peaks2 and3 were presentinsmall and
variable amounts and were absent from some
preparations;chromatographyonDowex-50
in-dicated thatpeaks2and3 arem6Ade and 2'-O-methylribose, presumably degradation
prod-ucts of N6, 2'-O-dimethyladenosine.
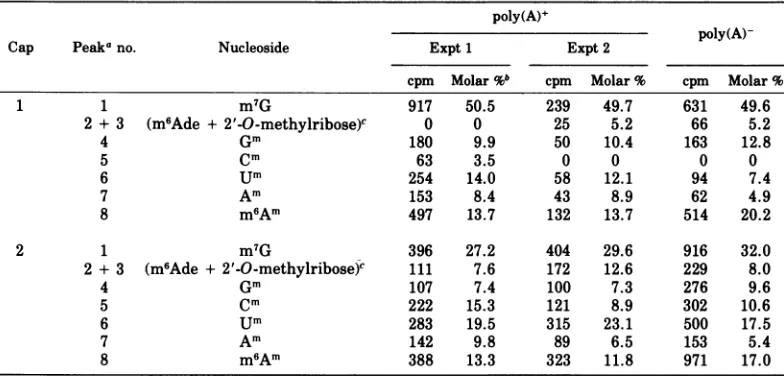
Table 2 gives the radioactivity and molar percent ofeachmethylatednucleosideincap1 andcap 2fractions fromtwodifferent prepara-tions of poly(A)+ and one preparation of poly(A)- Ad2 mRNA. Themolar ratio of m7Gto
2'-O-methylnucleosides was approximately 1.0
for the cap 1 fractions and 0.5 for the cap 2 fractions. Analysesof threeadditional prepara-tionsofearlyAd2poly(A)+mRNA (notshown)
10
10 I
0
E a. to
5
10
5
200
)2±34
X<,5 60 10 20 30
DISTANCE FROM ORIGIN
(CM)
FIG. 3. Paper chromatography of methyl-3H-la-beled cap1and cap2fractions of early Ad2 mRNA
after digestion with venom phosphodiesterase and alkaline phosphatase. Enzyme digestion and paper chromatography wereperformed as describedin Ma-terials and Methods. The shadowed circles indicate nucleoside markers, the numbers correspond to those
inTable 2, and thearrowsindicatethe solvent front. The paper was cutinto 0.5-cm pieces, and nucleo-sides were elutedin 0.01 NHCl and counted in a
scintillator.(A) Cap1 of poly(A)+Ad2 mRNA; (B) cap2 ofpoly(A)+Ad2mRNA; (C) cap1 ofpoly(A)-Ad2 mRNA; (D) cap2ofpoly(A)-Ad2 mRNA.
gave similarratios with some variation inthe molar percent ofindividual 3H-methylated
nu-cleosidesbut nodifferencesinthespecific meth-ylated components detected. This variation is
expected ifthe relative amounts of different viral mRNA molecules fluctuated indifferent preparations andifeachniRNAhasadifferent cap. Although trace amounts of Cm (less than 3% oftotal 3Hcpm) were found insome cap 1 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.499.272.464.68.431.2]CAP SEQUENCES IN Ad2 EARLY mRNA
0
E C 0 cqJ w 0
z
D
03
C,) co en
4
0
x
E a. U
I')
[image:7.499.104.386.66.361.2]TUBE NUMBER
FIG. 4. Identificationofm7Gand m6AmbyDowex-50columnchromatography. Peaks1and 8 inFig.3were eluted with 0.01 N HCI, dried,and dissolved in 0.1 (A)or(B)0.2 mlof1NHCl containing1 to 3 OD260
unitseachofpG,pm7G,and m6A markers. Thesamplewasdepurinated, diluted to 0.02 NHCl,andloaded
onaDowex-50column. The column was washed with 1 mlof0.02NHClandelutedwith 0.5NHCIand 0.7N HCI.Fractions (0.6ml) werecollected, OD2.was measured, and eachfraction was dissolved in 10 mlof Biofluorand counted. The position ofeach base wasestablished byitsUV spectrum.(A)Analysisofm7G; (B) analysis ofm6Am.
preparations (Fig. 2), the amount was
varia-ble, and Cm was absent from several
prepara-tions.Cm in cap1mayrepresentminor
contam-ination with cell mRNA ormorelikely
cross-contamination onDEAE-Sephadexwith cap2.
Onthe basis of these nucleosideanalyses, four
cap 1 and atleast three cap 2sequences exist:
m7G(5')ppp(5')Nmp
withNm asAm,Um,
Gm,orm6Am; and
m7G(5')ppp(5')NmpNmp
with Nm aspairs of Am, Um, Cm,Gm, orm6Am.
To ruleoutpossiblecontaminationwithcell
mRNA,poly(A)+ and poly(A)- viralmRNA
se-lected by hybridizationtoAd2 DNA was
puri-fied by a second hybridization to viral DNA.
RNase T2-stable cap 1 and cap 2 fractions wereisolated andanalyzed. Noqualitative
dif-ferences were found in the3H-methylated
nu-cleoside components of once and twice hybrid-ized viral mRNA.
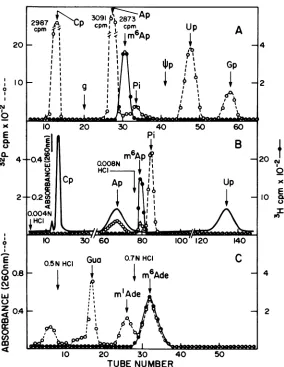
Analysesofthemononucleotidefraction. It
is not possible to determine directly whether 3H-labeled nucleotides are presentinthe
mon-onucleotide fraction (-2 charge) eluted from DEAE-Sephadex (Fig. 2) due to the large 32P counts which can mask small amounts of 3H counts.To detect internal basemethylationsin Ad2 early mRNA, the mononucleotidefraction from DEAE-Sephadex (Fig. 2) was chromato-graphedon aDowex-1 column (15).
The four 3P-labeled ribonucleotides eluted in the orderCp, Ap, Up, and Gp andpossessedno detectable 3H cpm; thus, the incorporation of methyl-3H into purinerings isnegligible (Fig. 5A). A distinct 3H peak was identified that eluted afterAp and beforeinorganicphosphate. The peak was furtherpurified by re-chromatog-raphy on Dowex-1 (Fig. 5B), depurinated, and
chromatographed on Dowex-50 (Fig. 5C). The
resuilting base was identified as m6Ade by com-parison with standards including m'Ade and m6Ade. The molar ratio ofm6Ap to total nucleo-tides in viral mRNA was 1 in 450, in both poly(A)+ andpoly(A)- RNA, asmeasured from the32Pcontent ofm6Ap elution from Dowex-1.
VOL. 20, 1976 431
on November 10, 2019 by guest
http://jvi.asm.org/
432 HASHIMOTO AND GREEN
TABLE 2. Composition of methylated oligonucleotides in Ad 2 early mRNA Cap 1 and Cap 2
poly(A)+
poly(A)-Cap Peaka no. Nucleoside Expt 1 Expt 2
cpm Molar%b cpm Molar % cpm Molar %
1 1 m7G 917 50.5 239 49.7 631 49.6
2 + 3 (m6Ade +2'-O-methylribose)c 0 0 25 5.2 66 5.2
4 Gm 180 9.9 50 10.4 163 12.8
5 Cm 63 3.5 0 0 0 0
6 um 254 14.0 58 12.1 94 7.4
7 Am 153 8.4 43 8.9 62 4.9
8 m6Am 497 13.7 132 13.7 514 20.2
2 1 m7G 396 27.2 404 29.6 916 32.0
2 + 3 (m6Ade + 2'-O-methylribose)C 111 7.6 172 12.6 229 8.0
4 Gm 107 7.4 100 7.3 276 9.6
5 Cm 222 15.3 121 8.9 302 10.6
6 um 283 19.5 315 23.1 500 17.5
7 Am 142 9.8 89 6.5 153 5.4
8 m6Am 388 13.3 323 11.8 971 17.0
aPeak numbers correspond to those in Fig. 3.
bMoles of each methylated nucleoside/100 mol of methylated nucleosides present in the cap. Two moles of
methyl residue of peak 8 were calculated as1mol of m6Am. Putative degradation products (peak 2 and 3) were notadded tom6Am values.
cTentative analysis (see text). Molar percent of these peaks was calculated with one mole of methyl
residue corresponding to one mole of the constituent.
DISCUSSION
Poly(A)-terminated polyribosomal. early mRNA was double labeled with
[methyl-3H]-methionine and32po4 in the presenceof
cyclo-heximide (to enhance the yield of early mRNA) andAd2-specificmRNApurified by hybridiza-tion to and eluhybridiza-tion fromAd2 DNA immobilized
onnitrocellulosefilters. The RNAwasdigested
with RNase T2 and chromatographed on
DEAE-Sephadex in 7M urea atpH 7.6. A
ra-dioactive fraction ofcharge -2 (mononucleo-tide)wasobserved, and twoadditional fractions
of charges -5 (cap 1) to -6 (cap 2). These
fractions were further characterized by paper
and column chromatography. m6Ap was the only methylated constituent in the
mononu-cleotide fraction. m7G andthe2'--methyl
de-rivatives ofA, G, U, andm6Awerefound in the cap 1fraction. Thecap2 fraction contained all
the constituents of the cap 1plus 2'--methyl C. The molar ratiosof m7G to the 2'-O-methyl-ated derivativeswere 1.0 and 0.5 for the cap 1
andcap2fractions, respectively. Sincethe cap
1 and cap 2 fractions apparently represent 5'
termini, these data indicate that there are at
leastseven different 5' termini, and therefore atleast sevenearly viral genes (unless a
spe-cific mRNA molecule can contain more than
onetypeof 5'terminus). Baseduponthe
analy-ses of 5' termini of vesicular stomatitis virus and reovirus mRNA's, we conclude that the.
general structures of these termini are:
m7G(5')ppp(5')Nmp(cap 1) and
m7G(5')ppp(5')-NmpNmp(cap 2). Four cap 1 structures exist with Nm = Gm, Um, Am, and m6Am. Three is a
minimumand25 is a maximum number for cap 2 structures (25 different pairs of five methyl-atednucleosides canexist). The existence of at
leastsevenmethyl caps is interesting in view of
the early Ad2mRNA mapping studies of Craig et al. (7, 8) and Buttner et al. (J. Mol. Biol., in press) which suggest a minimum of five to
eight early mRNAspecies. Inaddition,we have
recently obtained evidence of 11 Ad2-induced
early proteins, eight which were not detected in appreciable quantity in uninfected cells
(Woldet al., inpress). Thus, more than seven
earlygenes mayexist.
The significance of multiplecap structurein early Ad2 mRNA is unknown. Compositional
analysisindicates that differentcap structures
are notminor components.Analysis of lateAd2 mRNAby Moss and Koczot (21) gave a much
simpler pattern of two cap 1 structures
(m7Gpppm6Am and m7GpppAm); thecap 2 frac-tion was not fully characterized. The marked
differencesbetweenearly and late Ad2 mRNA
methylation patterns may suggest viral
in-ducedmodification. Two prominentfeatures of late Ad2 gene expression are the switch from
early to late viral mRNA synthesis and the
block in host cell protein synthesis. Although
most, if not all, late Ad2 RNA sequences are
J.
on November 10, 2019 by guest
http://jvi.asm.org/
CAP SEQUENCES IN Ad2 EARLY mRNA 433
20
0 N
0
x
E CM. U
a.
NV C,
0
x
E
a.
C.
XI
9E
o 10 30" 60 80 100"120 140
c 0.5NHCI Gua 0.7NHCI C
° 0.8 -
m6Ade
4w 0~I
w
~~~~~~~m
IAde
C-)
Z 04 2
4Zo.0
10 20 30 40 50
TUBE NUMBER
FIG. 5. Analysis of m6Ap in the mononucleotide fraction. The mononucleotide fraction of Fig. 2A was pooled, diluted fivefold with water, and loaded on aDowex-1 column(A). The column was equilibrated with 0.004 NHCland developed starting with fraction 20 with a concave gradient formed with 40 ml of 0.004 N
HClin themixing chamber and 20 mlofO0. MNaCIand 0.01 NHCIin thereservoir. One-milliliter fractions were collected,OD26 wasmeasured, and 0.1 ml ofeach fraction was counted. Each peak was identified by its UV spectrum at pH2 and 12 (0D260is not shown). Fractions 29 to 33 were pooled, neutralized, diluted fivefoldwith watercontaining 1OD26 uniteach of Cp, Ap, Up, andOpasmarkers, and loaded on aDowex-1
column(B). The column was equilibratedwith 0.004 NHCland elutionwas performed with 0.008 NHCl
after Ap was eluted. Fractions(1 ml) were collected and 0.3 ml of each was counted. Fractions 77 to 82 were pooled, dried, and dissolved in0.1 ml of 1 NHClcontaining 1 to 2 01)260 units ofpG, m'A andm6A as markers. The sample was depurinated and analyzed on a Dowex-50 column as shown in Fig. 4.
transcribed during early stages of infection,
they donotmature tomRNA andareretained
(ordegraded) inthe cell nucleus (31, 35). Late
after infection, the sameviral RNA sequences are processed to mRNA and transported to
polyribosomes. Conceivably this switch is a
consequenceofanalteredmethylationpattern.
Forexample, specificmethylated basesmaybe recognition sites for enzymes that transport mRNA out of the nucleus and onto polyri-bosomes, orforcleavage byputativeprocessing enzymes. Methylation may also "restrict"
de-struction of late viral mRNA precursor
mole-cules by nucleases. In addition, methylation
couldparticipate in the block of host cell pro-teinsynthesisthat occurslate after Ad2 infec-tion. Perhaps cell mRNA is not capped late
after infectionor capsare removedby specific pyrophosphatases, such as recently described
by Nuss et al. (24). Such enzymes may attack
cell caps which aresimilartosomeearlyviral mRNAcaps;late mRNA caps maybe different. Viral-induced enzymes that affect or
mod-ify methylation are aninterestingpossibility. VOL. 20, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.499.103.387.73.440.2]434 HASHIMOTO AND GREEN
The m7Gportion ofthe 5' terminal caphas been reported necessary for translation of mRNA (4, 23), but the biological functions of the penultimate 2'-0-methylation and
inter-nalm6A are not known. Since the methyl caps of each or most early mRNA's are different, perhapseach mRNA is metabolizeddifferently accordingtoits capstructure. Aless interesting explanation of multiple caps is that the type of 2'--methylated constituentsinthe cap is
irrel-evanttothe biological function of the cap. That is,the terminalm7G is presumably added post-transcriptionally, andthe penultimate methyl-ated base may result from nonspecific 2'-O-methylation after the capping reaction. Whether other internal base sequences located close to the 5' terminus serve as signals for capping, cleavage, or methylating enzymes is
unknown. Finally, we pointout that we have notstudiedthe effectofcell culture conditions
on the methylation patterns of early Ad2
mRNA, and it is possible that different results would be obtained underdifferent RNA label-ing conditions. However, preliminary analyses ofAd2 mRNA prepared without use of cyclo-heximide have revealed no substantial
differ-ences in methylation patterns from those
de-scribed here.
ACKNOWLEDGMENTS
Wethank W.Buttnerfor helpful discussion on prepara-tionof Ad2 early mRNA, S. Lowery for technical assistance, and W.S. M. Wold for critical evaluation of the manuscript. This investigation was supported by Public Health Serv-ice grant AI-01725 from the National Institute ofAllergy and Infectious Diseases and by contract NO1 CP43359 within the Virus Cancer Program of the National Cancer Program of the National Cancer Institute. S. H. was partly supported by Naito Research grant for travel ex-penses. M. G. is a Research Career Awardee
(5K6-AI-4739) of the National Institute ofAllergy and Infectious Diseases.
LITERATURE CITED
1. Abraham, G., D. P. Rhodes, and A. K. Banerjee. 1975.
The5' terminal structureofthemethylated mRNA
synthesized in vitro by vesicular stomatitis virus. Cell 5:51-58.
2. Ali-Arif, A., and M. B. Sporn. 1972. An analytical methodfor theseparation ofsugar-methylated ribo-nucleosidesfrombase-methylated andnonmethylated
ribonucleosides. Anal. Biochem. 48:386-393. 3. Aloni, Y. 1975. Methylated SV40 mRNAs. FEBSLett.
54:363-367.
4. Both, G. W., A. K.Banerjee, andA.J.Shatkin. 1975. Methylation-dependent translation of viral messen-gerRNAsin vitro. Proc. Natl. Acad. Sci. U. S. A. 71:2951-2955.
5. Buttner, W., Z. Veres-Molnar, and M. Green. 1974. Isolation of DNA strand-specific early messenger RNAspeciesincell infectedbyhumanadenovirus2 infected cells. Proc. Natl. Acad. Sci. U. S.A. 71:2951-2955.
5a. Chinnadurai, G., H. M. Rho, R. B. Horton, and M. Green. 1976. mRNA from the transforming segment of theadenovirus 2 genome in productivelyinfected
and transformed cells. J.Virol. 20:255-263.
6. Cory, S., and J. M. Adams. 1975. The modified 5'-terminal sequences in messenger RNA ofmouse mye-loma cells. J. Mol. Biol. 99:519-547.
7. Craig, E. A., M. McGrogan, C. Mulder, and H. J. Raskas.1975. Identification ofearlyadenovirus type 2RNAspecies transcribed from the left-hand end of thegenome. J. Virol. 16:905-912.
8. Craig,E.A.,S.Zimmer,and H. J. Raskas.1975. Anal-ysis of early adenovirus 2 RNAusing Eco R * Rl viral DNAfragments. J. Virol 15:1202-1213.
9. Fernandez, S. M., and H. J. Raskas. 1975.Methylation ofcytoplasmic adenovirus RNA synthesized early and late in productive infection. Biochem. Biophys. Res. Commun. 66:67-74.
10. Furuichi, Y.1974."Methylation-coupled" transcription by virus-associated transcriptase ofcytoplasmic poly-hedrosis virus containingdouble-stranded RNA. Nu-cleic Acids Res. 1:809-818.
11. Furuichi, Y., M. Morgan, S.Muthukrishnan, and A. J. Shatkin. 1975. Reovirus messenger RNA contains a methylated, blocked 5-terminal structure: m7G(5')ppp(5')GmpCp-. Proc. Natl. Acad. Sci. U. S. A. 72:362-366.
12. Furuichi, Y., M. Morgan, A. H. Shatkin, W.Jelinek, M.Salditt-Georgieff, and J. E. Darnell. 1975. Meth-ylated, blocked 5' termini in HeLacell mRNA. Proc. Natl.Acad. Sci. U. S. A.72:1904-1908.
13. Furuichi, Y., and K. I. Miura. 1975. A blocked struc-ture atthe 5' terminus of mRNA fromcytoplasmic polyhedrosis virus. Nature (London) 253:374-375. 14. Green, M., J. T. Parsons, M. Pina, K. Fujinaga, H.
Caffier, and I.Landgraf-Leurs. 1970. Transcription of adenovirus genes inproductively infected and in transformed cells. ColdSpringHarborSymp. Quant. Biol.35:803-818.
14a. Harter, M. L., G. Shanmugam, W. S. M. Wold, and M.Green. 1976. Dection of adenovirus type 2-induced early polypeptides using cycloheximide pretreatment toenhanceviral protein synthesis. J. Virol. 19:232-242. 15. Hashimoto, S., and M. Muramatsu. 1973. Differences
in nucleotide sequences of ribosomal RNA between
the liver and ahepatomaof C3H/He mice. Eur. J. Biochem. 33:446-458.
16. Hashimoto,S., M.Sakai,and M.Muramatsu. 1975.
2'-0-Methylatedoligonucleotidesinribosomal 18S and 28S RNAof a mousehepatoma,MH134.Biochemistry 14:1956-1964.
17. Iwanami,Y., andG. M. Brown. 1968. Methylated bases of transfer ribonucleic acid from HeLa and L cells. Arch. Biochem. Biophys. 124:472-482.
18. Lavi, S. and A. Shatkin. 1975.Methylated simian virus 40-specific RNA from nuclei and cytoplasm of infected BSC-1 cells. Proc. Natl. Acad. Sci. U. S. A. 72:2012-2016.
19. Lewis, J. B., J.F.Atkins, P. R.Baum,B.Solem, and R. F.Gesteland. 1976. Location and identificationof the genes for adenovirus type 2 earlypolypeptides. Cell 7:141-151.
20. Lindberg, U., T. Persson, and L.Philipson.1972. Isola-tion and characterization of adenovirus messenger ribonucleic acid in productive infection. J. Virol. 10:909-919.
21. Moss,B., and F. Koczot.1976.Sequenceofmethylated
nucleotidesatthe5'terminus of adenovirusspecific RNA. J.Virol. 17:385-392.
22. Moyer, S.A., G.Abraham,R.Adler,and A.K. Baner-jee.1975.Methylatedand blocked5'terminiin vesic-ular stomatitis virus in vivo mRNAs. Cell 5:59-67. 23. Muthukrishnan,S.,G.W.Both,Y.Furuichi,and A. J.
Shatkin. 1975. 5-Terminal7-methylguanosine in eu-karyotic mRNA isrequiredfor translation. Nature (London)255:22-37.
24. Nuss, D.L., Y. Furuichi,G.Koch,and A. J. Shatkin.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
CAP SEQUENCES IN Ad2 EARLY mRNA 435
1975. Detection of HeLa cell extracts ofa 7-methyl
guanosine specific enzyme activity that cleaves m7GpppNm. Cell 6:21-27.
25. Parsons, J. T., and M. Green. 1971. Biochemical studies
on adenovirus multiplication, XVII. Resolution of
early virusspecific speciesinAd2infected and trans-formedcells.Virology 45:154-162.
26. Perry, R. P., andD. A. Kelley. 1974. Existence of meth-ylatedmessengerRNA inmouseLcells. Cell 1:37-42.
27. Perry, R. P., D. E. Kelley, K. Friderici, and F.
Rott-man. 1975. The methylated constituents of L cell messengerRNA. evidence foranunusual clusterat the 5'terminus. Cell 4:387-394.
28. Philipson, L., U. Pettersson, U. Lindberg, C. Tibbetts, B.Venmstrom,and T.Persson. 1974.RNASynthesis and processing in adenovirus infected cells. Cold Spring Harbor Symp. Quant. Biol. 39:447-456. 29. Rose, J. K. 1975. Heterogeneous 5-terminalstructures
occur onvesicular stomatitisvirus mRNAs. J. Biol.
Chem. 250:8098-8104.
30. Rottman, F., A. Shatkin, and R. P. Perry. 1974.
Se-quencescontaining methylated nucleotidesatthe 5' termini ofmessengerRNAs: possible implications for
processing. Cell 3:197-199.
31. Sharp, P. A., P. H. Gallimore, and S. J. Flint. 1974. Mapping of adenovirus 2 RNAsequencesinlytically
infectedcellsand transformed cell lines. Cold Spring HarborSymp.Quant. Biol. 39:457-474.
32. Thomas, D. C., and M. Green. 1969. Biochemical stud-iesonadenovirus multiplication XV. Transcription of the adenovirustype2genomeduring productive
in-fection.Virology39:205-210.
33. Wei, C. M., A.Gershowitz, and B. Moss. 1975. Methyl-ated nucleotidesblock 5' terminus of HeLa cell mes-sengerRNA.Cell 4:379-386.
34. Wei, C. M., and B. Moss. 1975.Methylated nucleotides block 5'-terminus of vaccinia virusmessengerRNA.
Proc. Natl. Acad. Sci. U. S. A. 72:318-322. 35. Wold, W. S. M., M. Green, K. H. Brackman, M. A.
Cartas,and C. Devine. 1976. Genome expression and mRNA maturationatlatestagesofproductive adeno-virustype2infection.J. Virol. 20:465-477. 36. Wold, W. S. M., M. Green, and T. W. Munns. 1976.
Methylation of late adenovirus 2 nuclear and messen-gerRNA.Biochem. Biophys. Res. Commun. 68:643-649.
VOL. 20, 19.76
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 2.Poly(A)pancreaticSephadexMethods. Chromatography of RNase T2 digest of 32p-, [methyl-3H]labeled Ad2 early mRNA on a DEAE- column in 7 M urea at pH 7.6](https://thumb-us.123doks.com/thumbv2/123dok_us/1552758.107825/5.499.102.393.67.436/pancreaticsephadexmethods-chromatography-rnase-digest-methyl-labeled-early-column.webp)