Copyright © 1977 AmericanSocietyforMicrobiology Printed inU.S.A.
Replication
Process
of the Parvovirus
H-1.
VII. Electron Microscopy of Replicative-Form DNA Synthesis
IRWIN I. SINGER AND SOLON L. RHODE III*
PutnamMemorial Hospital Institute for Medical Research, Bennington, Vermont 05201
Receivedfor publication 11 June 1976
The geometry of replicative form (RF) DNA synthesis of theH-1 parvovirus
wasstudied withthe electron microscope usingformamideor aqueous variations
of theKleinschmidt spreading procedure. H-1DNAwasisolated from human or
hamster cells infectedwith a temperature-sensitive mutant, tsl, whichis
defi-cient inprogenysingle-stranded DNAsynthesis at therestrictive temperature
(S.L.Rhode, 1976),thusminimizingpossible confusionbetweenRFandprogeny
DNA replicative intermediates (RIs). The purity of the isolated H-1 DNA, as
determinedby gel electrophoresis, ethidium bromide staining, autoradiography,
and digestionwithendoR EcoRI, was high. H-1 RF DNAs werelinear
double-stranded molecules, 1.53 ,m in length. H-1 RIs of RF DNA replication were
double-stranded, Y-shaped molecules, with thesame length as RF DNAs. The
replicationorigin was localized no more than 0.15 genome lengths from one end
oftheRFDNA, withreplication proceedingtoward the other end at a uniform rate. SimilarRFand RI molecules of dimer size were also observed. The length
of H-1 single-stranded DNA extracted from purified virions was measured
relative to thatof
OX174
and it had a verysimilar contour length, so that themolecularweightof H-1single-stranded DNA wouldbe at least 1.48 x 106 to 1.59
x 106 (Berkowitz and Day, 1974).
The parvovirus H-1 contains a single- ceedsfrom the end containing the origin to the
stranded(ss) DNA(22),and iscapable ofauton- opposite terminus at a uniform rate. The
omousreplication(12, 20, 21).During infection, branched RI DNA appeared to be entirely ds.
a double-stranded (ds) replicative form (RF) Dimer-length RF DNA molecules andRIswere
DNA is synthesizedand replicated semiconser- alsoobserved. The lengths ofH-1viral andRF vatively at a nearly exponential rate (13). Prog- DNArelative to those of
pX174
weremeasured,eny ssDNA is produced simultaneously, pre- and the molecular weight of H-1 ssDNA was
sumably by displacement fromRF DNA engag- determined tobe at least 1.48 x 106 to 1.59 x
ing in asymmetric DNA synthesis (14), and 106,similar to the value obtained by gel
electro-encapsidatedinto virions. Inthisstudywehave phoresis (16).Additional data on the location of
examined H-1 replicative form DNA and its the origin ofreplication has been obtained by
replicative intermediates (RI DNA) with the partialdenaturation mapping, and will be
pre-electron microscope to define the geometry of sentedinthe followingpaper of this series(19).
RF DNA replication. To minimize any
confu-sionwith progenyviralDNAsynthesis, we an- MATERIALS AND METHODS
alyzed the replicating DNA of a temperature- Virus and cells. Parasynchronouscultures of
sec-sensitivemutantof H-1,tsl, which isdeficient ondary hamster embryo fibroblasts or human NB
in progeny ssDNA synthesis, but not in RF cells were prepared and infected with tsl or
wild-DNAreplication (15). type (wt)
H-1
as previously described (16).Esche-Previous studies using ethidium bromide- richia coli H-502 and H-4714, and
OX174
(wt) andCsCl density gradient centrifugation, velocity am3 werekindly provided by R. L. Sinsheimer.
sedimentation, and gel electrophoresis pro- Viral DNApreparation. H-1 virion DNAwas
ex-duced noevidenceforcovalentlyclosed circular tracted from purified virus (12) labeled with H-1 RF DNA(13). Electron microscope visualiz- [3H]thymidine ([3H]TdR) by lysis in 0.2 N NaOH and ationof H-1 RF DNA reveals it to be alinear
centrifugation
inanalkalinesucrosegradient
(16).ationlofuH-i
R.
DAm
reveal.
itatosbe
alin
OX174
viral DNA was prepared as in (11), andmolecule,
1.53um
inlength.
Analysis of RIs XX174 RF DNA was produced as outlined byJohn-shows thattheyareY-shaped, with the origin son and Sinsheimer (8).
H-1
RF DNA was labeledofreplicationlocated no more than 0.15 genome andprepared as previously using the Hirt extraction
lengthsfrom oneend, and that replication pro- withPronase digestion (16). The Hirtsupernatants
713
on November 10, 2019 by guest
http://jvi.asm.org/
714 SINGER AND RHODE J. VIROL.
containing viral DNA were extracted with phenol stubs and their attached grids. The samples were
after RNase treatment and before fractionation in shadowed by melting a 2-cm segment of platinum-the preparative neutral sucrosegradients. Benzoyl- palladium (80:20) wireina vacuumof2x 10-5mm of ated DEAE-cellulose (BDC) chromatography was Hg; a visible droplet of molten metal was
main-carried out asdescribed (16). tained on the tungsten electrode for the 1-min Gel electrophoresis. Agarose gel electrophoresis shadowing period while the table rotatedat120rpm. incylindrical gels has been described (16). Vertical The grids were shadowedata50angle 5cmfrom the gel electrophoresis was performedon anEC470 elec- metal source, and were then easily removed from trophoresis cell (E-C Apparatus Corp., Philadel- their stubs with fine forceps. Micrographs were phia, Pa.) with a gel (0.3 by 12 by 16 cm) of 1% made with a JEM-7 electron microscope (J.E.O.L. agarose.The gel and electrode buffers were buffer E Co., Inc., Medford, Mass.) equipped with liquid-(40 mM Tris[pH 7.2]-20 mM Na acetate-1 mM nitrogen-filled cold traps at the diffusion pump and EDTA) asdetailed (5). Electrophoresis was carried on topofthe objective lens pole piece (immediately out withaconstant voltage of 100Vat16°C untilthe beneath the specimen), and a30-,tmgold foil objec-bromophenol blue marker was near the bottom of tive aperture. The lens currents and high voltage the gel. The gels were stained in E buffer with 5 ,ug (80kV) were turned on at least 1 h before studying of ethidium bromide per ml for1handphotographed grids at a magnification of 14,200, calibrated fre-under illumination with long-UV light. Autoradi- quently with a grating replica (E. Fullam, Inc., ographs were made of the gelafter vacuum drying Schenectady, N.Y.). The intermediate lens current with Kodak no-screen X-ray film NS2T, exposed (which controls magnification on this instrument) for 24 hat23°C. was neverchanged once a micrograph of the calibra-Electron microscopy. Viral DNA, which had tion replica was made; the output magnification been banded to equilibriuminCs2SO4, was dialyzed remained constant throughout the course of this against 0.1MTris(pH 8.5)-O.01MEDTA, broughtto work. Care was also taken to use a low
electron-0.2 M inNa acetate, precipitated with ethanol, and beam current (not exceeding 10
AA)
and a reduced dissolved in 50 to 100gl
ofthe latter buffer for amount ofcondensor illumination to minimize dam-electron microscopic study. Most of the DNA was age to the specimens. The electron micrographs prepared with the formamide technique. The spread- were enlarged 10 times by projection so that theingsolutior.contained10llofwater,10
gl
of an0.5- DNA molecules could be accurately traced; theirmg/ml solution of cytochrome c (Sigma, type III, in contour lengths were measured with a Dietzgen 0.5 M Tris-hydrochloride[pH 8.5]-0.05 M EDTA), planimeter, and expressed as the mean plus or
mi-5
gl
of DNA sample in 0.1 M Tris-hydrochlo- nus the99% confidence interval (CI). ride(pH 8.5)-0.01 M EDTA, and 25 ,ul of formamide(Matheson Scientific, Inc.). Immediately after thor- RESULTS ough mixing, the entire50
gl
of DNA solutionwasspread onto a freshly prepared hypophase of 20% Purification ofH-1 RF and RI DNA from formamide in 10 mM Tris-hydrochloride(pH 8.5)-i infected cells.
H-1
DNA was extracted from mMEDTA, in a Teflon-coated trough as describedtsl-infected
cultures of parasynchronous hu-by Davis et al. (4). DNA was also spread hu-by the man NB cells orsecondary
hamster embryo aqueous method asoutlined by the latterauthors. fibroblastsbythe method ofHirtaspreviously Parlodion-coated 300-mesh copper grids were diled bythemet
supertas wre sub-touchedto thehypophase surface 1 min after DNA detailed (16). The Hirt supernatants were sub-spreading; no talc was used. The grids were attached jected tovelocity
sedimentation in a prepara-to stubs to avoid damaging their parlodion films tivesucrosegradient,
andfractionswerepooled when handling them with forceps, sampling the asillustratedinFig. 1. Pool A contains primar-DNA, or during staining, dehydration, and shadow- ily monomer RF DNA, and pool B contains ing. To dothis, microscope slides were dipped into a monomer RF DNA, dimer RF DNA, and RI1%solution of parlodion in amyl acetate (wt/vol) and
molecules,
which sediment more rapidly than allowedtodry vertically. The parlodion films were monomerRF DNA (16). Mock-infected cultures stripped ontoadistilledwatersurface, and portionssimer
extrA ctedcultures
having asilver-gold interference color were pickedwere
similarly
extracted, and
theyieldof radio-up with awire loop, air dried, and placed onto the labeled DNA in these regions of the sucrose surface of agrid resting on a stub; theoverhanging gradient was less than 2% of that from H-1-edgesof the parlodionfilm firmly held the grid on infected cultures. Thehomogeneity
of the ra-the stub. This method minimizes damage to the diolabeled DNA to competitive hybridization parlodionfilm, which increases length variation of (13), or to cleavage by bacterial restriction en-the adherent DNAmolecules; contamination of the donucleases (16) indicated that theH-1
DNAs lowersurface of thegrid by the various solutions is were at least 90% radiochemically pure. How-alsoprevented. Thegridswere stainedimmediately ever itafter DNAsampling with freshly prepared 50 HM
e,
Spossible
thatmock-infected
cultures uranyl acetate-50 ,MHCl in90% ethanol (pH3.9)are
inadequate controls for
purity. For exam-for 30s, followed by a 10-s rinse in 90% ethanoland ple, the cytopathic effects ofH-iinfection might dehydration with 2-methylbutane for 10 s. Shadow- inducedegradation
ofcellularDNA,
although ing wasaccomplished with a Denton vacuum evapo- this was not observed when cells with prela-rator andarotary table, which accommodated the beled DNA were infected (13), orcontaminationon November 10, 2019 by guest
http://jvi.asm.org/
SYNTHESIS
l w 0 ' _ with
long-UV
light (Fig. 2a). As shown before15 _ A (16), the predominant bands are the monomer
RF DNA in the mixtureofA and B and in A
alone, dimer RF in B alone, and thepartially
cleaved dimer RF, EcoRI-A, dimer B, and B
fragments in the digest of A-plus-B mixture.
Thesestained DNAbandswereshownto
corre-10 spond to the radiolabeled DNA by an
autora-diograph of the gel (Fig. 2b). Thus, after the
sucrosegradient step, thepreparations consist
largely of monomer and dimer H-1 RF DNAs,
x asjudged by electrophoreticmobility and
speci-CL lficityofcleavage with EcoRI. It should be noted
I / thatthemonomerand dimerRFbands,aswell
asthose of the EcoRI-A and -Bfragments,
ap-5/
pear as doublets in the ethidiumbromide-stained gel, butnot inthe autoradiogram. This
difference in resolution isprobably due to our
deliberate overexposure ofthe autoradiogram
to
visualize the
light
bands
inthe
EcoRIdigest.
B I A The occurrence of two distinct
EcoRI-B
frag-0 , ments has
already
been documented(16);
the5 10 15 20 25 possible existence of doublet monomer and
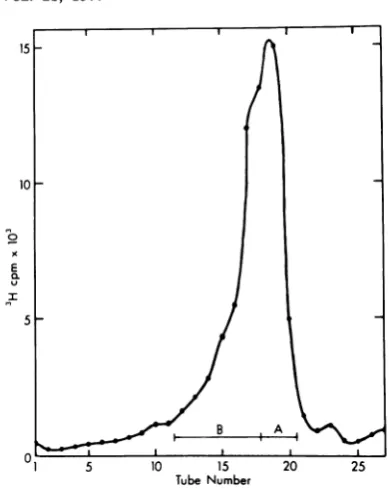
di-FIG. 1. Preparative sucrosegradient
ofPH]BUdR
mer RFs and EcoRI-A fragments iscurrentlycontaining H-1 RF DNA. Parasynchronous cultures under
investigation.
Also,thematerial remain-of hamster embryo cells infected with tsl H-1 at a ing at the origin of thelanes containing DNA multiplicity of infection of 5 to 10 PFU/cell were from fraction Bof the sucrose gradientis pre-incubated at 39.5°C. The cultures were treated with sumed to be entangled molecules not greater FUdR (10.5m.g/ml)
14 to 14.5 h p.i., and labeled than dimer size, since electron microscopywith[3H]BUdR as inResults. Viral DNA extracted never revealed longerDNAs in fraction B, and
by the Hirt method was redissolved in 50 mM thisentrapmentisnotconsistentlyobserved. It Tris(pH 7.5)-i mMEDTA and treated with pan- ispossible that this material is associated with creatic RNase (50
mg/ml)
for 30 min at 37°C. RNasewas removed by phenolextraction and the DNA was
protein,
but webelieve
that this is unlikely,precipitated with 0.15 MNaCl and 2.5volumes of sice theDNA was digested withPronase, and
ethanol at-20°C for 16 h. The DNA was redissolved extracted with phenol.
inthegradient buffer and sedimented in a 5 to 20% The DNA prepared for electron microscope sucrosegradient for 18h at24,000 rpm,4°C,in an analysis was density labeled with [3H]bromo-SW27 rotor as done previously (16). Fractionsofi ml deoxyuridine([3H]BUdR). Specifically, H-1 tsl-werecollectedthrough the bottom ofthe tube, and 20- infected NB cultures at39.5°C were incubated
pi
aliquots used to determine the positions of radio- with medium containing 5'-fluorodeoxyuridine activity.Regions were pooled as illustrated, precipi- (FUdR) and BUdR(10-5
M) 14 to 16 h p.i. tated with ethanol, and redissolved in 20 mM and then FUdR with [3H]BUdR (2gCi/ml,
Tris(pH8.0)-i
mM EDTA-0.15% Sarkosyl before a X 106 M) 16to
18 hp.i. Similarly,
ts-l
isopycniccentrifugation inCsSoe4.
The directionofinfected
hamster
embryo
cells at
39.5°C
were
sedimentation isfromrighttoleft. incubated with FUdR and BUdR 14 to 14.5 hp.i., and then FUdR and[3H]BUdRat 14.5 towith
degraded unlabeled
cellular DNAmaybe 16 hp.i. After Hirtextraction and sucrosegra-inexcessof the viral DNA. Such contamination dient centrifugation, the A and B pools (from
wasruledoutby comparing
radiolabeled
DNA Fig. 1) werebanded to equilibrium in Cs2SO4tototal DNA byagarosegel electrophoresis. H- gradients (Fig. 3A and B). All DNA greater
1tsl viralDNAlabeled for 12 to16hpostinfec- than hybriddensity (arrow indicates adensity
tion (p.i.) with 32p was prepared through the of 1.440g/cm3)waspooled,anddialyzedagainst
sucrose gradientstep. Equal portions ofpool A 10 mMTris(pH
8.5)-i
mM EDTA forstudy withand
pool
B (Fig. 1) were analyzed separately theelectronmicroscope.Inthis way,theprepa-andcombined bothwith and withoutdigestion ration was enriched formolecules that had
rep-with endoR EcoRIinaslabgel(3 mmby12cm licated one or more times in the presence of
by 16 cm) of1% agarose. The gel was stained BUdR, and controlexperiments indicatedthat
with ethidium bromide, and the fluorescent 98% ofcontaminating light DNA wasremoved.
DNA was photographed under illumination Thecompositionofthe final dense H-1 DNA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.49.244.55.300.2]716 SINGER AND RHODE J. VIROL.
,..-~U-_~~~~~~~~
FL
FIG. 2. Verticalslab-gelelectrophoresis ofH-iDNAaftercentrifugationinneutralsucrose. Parasynchron-ousNBcultureswereinfectedwithtslH-iat39.50Candlabeled with32P04from12to16hp.i.(16). The viral DNA was extracted, treated with RNase,and subjectedtopreparative centrifugation in a neutralsucrose
gradient.Fractions werepooledasinFig.1,andthe DNAwasdissolvedin 100p.lof10mMTris(pH7.5)-b mMNaCl-0.1 mMEDTA. Aliquotsofl10
MI
ofpoolsAand Bwerecombined andadjustedto50 mMNaCl,10 mMMgCl,,
and1 mMdithiothreitol, anddigested with100 UofEcoRIfor1 hat370C. The reaction wasstopped byadditionof20
tii
of2.5%sodiumdodecylsulfate-50%glycerol-10 mMEDTA.Slab-gel electropho-resiswascarriedoutasdescribedinMaterials and Methods. Thegel contained, from lefttoright:poolB(20pi),poolA (20pI), EcoRI-digestedA +B(10/.dof each) mixture,and A +B(20
PIl
each) mixture.Twentymicrolitersofeachsamplecontained theyield ofDNAfromabout2 X107NB cells. Thegelwasstained with ethidium bromide, and theDNAwasvisualized withlong-UVlight(a),andbyautoradiography(b). It has been shown(16) that thespecificH-iDNAspeciesobservedare(indescendingorder): dimer RF (DIRF),RF withattached EcoRI-Bfragment(RF +RIB,partialdigestofDIRF), RF,EcoRI-Afragment(RIA),dimer EcoRI-Bfragment(DI RIB),and EcoRI-Bfragment (RI B).
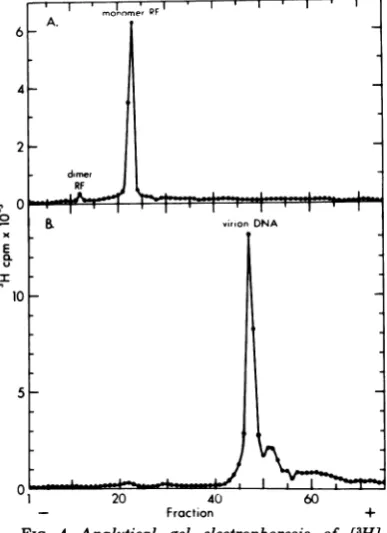
preparation
was examinedby
agarosegel
elec- Relative contourlengths
ofH-i andOX174
trophoresis (Fig.
4A) and found to be almost viral ssDNAs.Preliminary
electronmicro-entirely
monomerwithsmall amountsofdimerscopic
examination ofH-i ssviral DNAshowedRF DNA. The 13H]TdR-labeled virion DNA that it is
linear,
and hasacontourlength
simi-used in thisstudy
was alsoanalyzed by gel
lartothat of circular pX174 viral DNA. Sinceelectrophoresis
as shown inFig.
4B. The elec- we wanted to determine the molecularweight
tropherogram
of thelatterpreparation
isdomi- ofH-i ssDNA relative to4)X174
DNAby
mix-nated
by
ahomogeneous
peak,
with a smallering
thesepreparations
andmeasuring
the ratioportion
offaster-migrating species
assumed to of theirlengths
onthesamegrid,
itwasneces-be
fragmented
molecules. It should be noted sary to ensure thatsignificant
breakage
ofthatthe virion ssDNA
migrates
faster than theOX174
circleswasnotoccurring
sothatbroken ds RFunder these conditions and that there is (linear)4X174
ssDNA would not be confusedno evidence of virion DNA in the RF DNA with
H-i
ssDNA. We therefore examinedpreparation
(Fig.
4A). A very smallpeak
of4X174 ssDNA,
and found that the extent ofuncertain
significance
attheelectrophoretic
po- circlebreakage
was9%.These viral DNAsweresition of RF DNA is noted in the virion DNA then mixed such that the concentration ofH-i
electropherogram.
This could arise from an- DNA was twice that of the4X174;
themaxi-nealing
of V strands to traces (0.25%) of C mum amount of linear4X174
DNAcontami-strands,
butthis hasnot yetbeen proven.nating
the H-i viral DNApool
was thereforeon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.61.451.56.314.2]RF DNA
717
l l mum molecular weight of H-1 viral ssDNAis
10 A. D1.48 x 106 to 1.59 x 106, based on the 4X174
l1.50 ssDNA
molecular-weight
determination of1.59/1
45 x 106(1). In addition, we did not observecircu-1.45 larization (evidence of terminal
self-comple-1.40mentarity)
after incubation of H-1 viral DNA5
I5
- underannealing
conditions(50%
formamide-50
{\1.3 mM Tris-5 mM EDTA for 1.5 h at 23°C),
fol-lowed byimmediatespreading from50%
form-- amideonto20%formamide; theelectrophoretic
x JC\4 profile also was
unchanged
after thistreat-E O - ' ' s
~
t8 ment.X0- B. ,
Geometry
ofH-1 RF and RI dsDNA's.Re-1.50 gions of the
Cs2SO4
gradients containing
puta-tiveRI and RF(Fig. 3A andB), which exhibited
1\.45
[3H]BUdR-substituted
H-1 tsl DNA ofgreater1.40 than hybrid density, were chosen for electron
5 \ - l.40 microscope study, since it would be unlikely for
1-.35 the host cell to produce any DNA of this density
duetothe semiconservative nature of cellular
DNA synthesis, and the short
labeling
timesemployed. The
fully
substituted DNA frompool
_ s s , < s B (Fig. 3B) contained ds linear RF DNA of
1 5 10
15
20 monomerand dimerlengths (Fig.
5B),
andY-Tube Number
FIG. 3. Isopycnic centrifugation of H-1 tsl DNA
A.
moromerPFpools A and B from Fig. 1. The viral DNA pools 6 prepared by velocity sedimentation in the neutral
sucrosegradient (Fig.1)weresedimentedto
equilib-riumingradientsofCs2SO4aspreviously described 4_ (16). (A) Pool A; (B) pool B. Centrifugation condi-tionswere48hat35,000 rpmin atype 40fixed-angle rotor at 10°C. Fractions of 0.2 ml were collected 2
through the bottomofthetube, and
10-p.
aliquots 2weredriedon25-mmfilterpaper disksforassayof dimer
radioactivity. DNA greater thanhybrid density (ar- RF rows) waspooled, dialyzed against 0.1 M Tris(pH 0
8.5)-10mMEDTA, adjustedto0.3MNaCl,precipi- virionDNA tatedwithethanol,and redissolved in 50to100
pl
of E0.1 M Tris(pH8.5)-10 mM EDTA forelectron mi-croscopy. The direction ofsedimentation is from I
right to left. 10_
3% (DNA spreading solution contained 50%
formamide; hypophase contained20%
formam-ide). Figure5A is arepresentative
micrograph
5of this mixture, and histograms of the
mea-sured H-1 and 4X174 viral DNAs
(Fig. 6)
ex-hibit thesamemaximumpeaks(at 1.0,Lm)and
verysimilarcontour
length
variation.Both H-1and4X174 viral ssDNA's hada mean
length
of 01.0 amunder these formamide-spreading con- 1 20 40 60
ditions. We also
spread
a mixture ofH-1 and - Fraction +4X174
ssDNA's from a solutioncontaining 30% FIG. 4. Analytical gel electrophoresis of[3H]-formamide
ontoahypophase
with 10% formam-BUdR-containing
H-1 RF DNA and[3H]TdR-ide. Under these coditions, themean
length
of labeled H-1ssviral DNA.H-1RFDNA,
prepared
asHde.
Under
these96
codition,0umen
e 143 g-+- 99 in Fig. 3 (mixture ofpools A and B), and[3H]TdR-H-I
ssDNAwas0.96 ± 0.03z (n = 143 + 99 labeledvirion DNA were analyzed in a cylindricalCI) and thatofX174DNA was1.03 ± 0.02
ymI1
gel (0.6 by 15 cm) of 1.4% agarose (16). Electropho-(n = 123). Since gel electrophoresis revealed retic conditions were 30 Vfor 17.5 h at23C. (A)H-1some
fragmented
molecules(Fig. 4B),
themini- RFDNA; (B) H-1 virion DNA.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.52.240.51.315.2] [image:5.501.256.450.316.583.2]718 SINGER AND RHODE J. VIROL.
4
14~~~~~~~~~~~4
FI. Elcto
mirgah f-
Npedfo
0 ommdno2%fraie
a mV I R'Sieff
( *
of
l * t .24 : v DJ~~-~~ <<'s S
,z,-1*
E~
; S--*".'UIr 3
(Fig.5a7Ta
m,
m.;;*.; rlength
wa~1.53 -+ 0.0 um -n= 1 1) that of th1 2.9 0r2um( 2 o hie.Iiily
>-t
ie
;* 2 2 s t &*
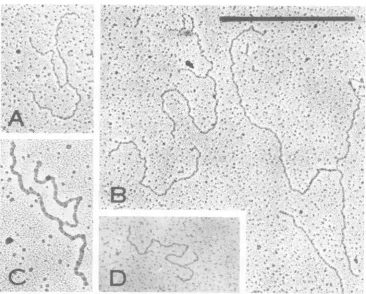
N-.,FIG 5 ElectronmicrographsofH-1DNAspreadfrom 50%formamide onto 20%formamide.Bar =nIzm.
(A)Mixctureof linear ss wtH-i andcircular ss4X174 viral DNAs; (B)tsl H-i ds RFDNA from pool B (Fig.
3B) (synthesized at the restrictive temperature,39.5°C), exhibiting monomer- and dimer-length molecules; (C)tsiH-i (39.5°C)ds RFmonomer(i)and ds RIsreplicated14%(2) and52%o(3)obtained from pool B (Fig. 3B) .
shaped RIs of both monomer and dimer size branch to that of the unreplicated base) were (Fig.
5C
and 7A-D). The mean RF monomer 1.55 ± 0.10gum
(n = 51) for the monomer, and length was 1.53 ± 0.04,um (n = 111); that ofthe 2.94 ± 0.22,rm
(n = 12) for the dimer. Initially, dimer was 3.10 ± 0.28 ,um (n = 16). Gorre- tsl H-i RIs were purified from synchronizedsponding
mean RIlengths
(each length
was hamsterembryo
fibroblastsorhuman NBcells.calculated by
adding
thelength
ofthelongest
The RIs of thesetwo groups wereidentical inon November 10, 2019 by guest
http://jvi.asm.org/
---vE ,-I .--- 20%I I of the length of the longer of the two
20 - arms; RF DNA lengths varied within 20% of the
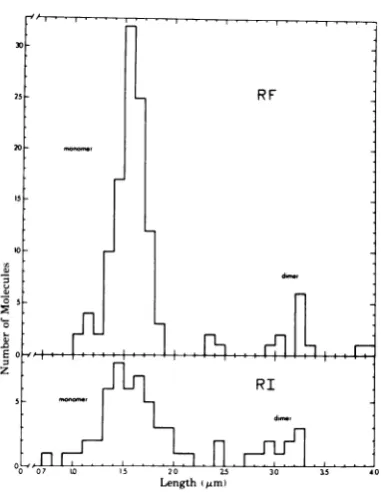
mean RFlength). Histogramsofthese RF and RI molecules exhibit analogous contour length 1S
*X-174
distributions ofmonomerand dimer DNAsub-populations (Fig. 8),
but thepercentage
of RIdimers (19%) was 1.4 times higher than thatof
RFdimers (13%). The molecular weight ofH-1
10_ monomer ds RF is 2.95 x 106, based on our
measurements of purified circular ds RF
OX174
spread separately under replicate conditions: X
1.65 ± 0.05 ,m, n = 15. Analysis of the
mon-@5. omer RF DNA peak (pool A) of the sucrose
gradient after Cs2SO4 purification (Fig. 3A)
X yieldedverysimilar
results, except
that theRIM o ~ E § § @ l @ | § contentwassixfold lower than in the RI
region,
%
30
__
pool B(Fig.
3B).
E _- Since it has been
reported
that ssDNAap-z pearsthinner and more twisted than dsDNAin
H-1 formamide preparations (4, 23), we expected to
observe obvious differencesinstrand
morphol-20 - ogy between RF, RI, and viral DNAs.However,
the appearance of H-1 ds RFs and RIs was
indistinguishable from that of ssH-1 or
OX174
viralDNAs (Fig.5A-C); therefore,the
possibil-ity that H-1 RI DNAs contain ss regionscould
not be excluded on the basis of their appearance
usingformamide-spreading solutions. We
con-sequently
studied
H-1 RI and RF DNAmole-cules
(isolated by BDCchromatography,
16)using the aqueous technique, which causes ss
o
L/ r-. .0 . regionsof the DNAtocondense into bushes (4).0.6 l.0o.5 All RI (Fig. 7C) and RFtsl H-1 DNA was fully
[image:7.501.48.242.63.399.2]Length(pm) extended and lacked bushes with this
method,
FIG. 6. Histograms of the contour lengths of cir- so that
H-1
RI and RF DNA must be largelyds;
cularsskX174 and linear ss H-1 wt viral DNAs. the mean RI length was 1.730.15
gm
(n
7),
These DNAs were combined in a spreading solution andthat of theRFwas1.46
±0.13
um(n 28)with 50% formamide, and cospread onto a hypo-
ungthis thnique.
Sgb e rophase containing 20%formamide; all micrographs using this technique. Single bushes were
ob-were taken from a single grid. The mean lengths of served on molecules of RF length when aqueous both H-i and X1 74 viral DNAs were 1.0
gm
(n = preparations of wild-type H-1 RI DNA were 50, 99% CI = +0.04,um for X1 74; n =97, 99% CI examined;these
moleculesarepresumably
in-=
±+0.05
,m for H-1).Notethat theH-1histogramis termediates in progeny ssDNA replication.skewed toward the lower end ofthe range, corre- A frequency distribution of
H-1
RIs spread sponding to the leading shoulder ofthe main analyti- from 50% formamide onto 20%formamide,
andcal-gel electrophoretic peak (Fig. 4B). ordered according to the proportion replicated,
isshownin Fig. 9; only unambiguously forked
size and distribution of replication fork posi- molecules whose
lengths ranged
from 1.0 to2.0tions, andweretherefore
pooled
for finalanaly-
,um(the
monomerlength
range)wereincluded.sis. Replicating molecules
constituted
33% of The amount ofreplication
varied from 0.15 tothis RF
population.
Seventeen percent of the 0.88 genomelengths;
the random distribution ofmeasuredRIpool exhibitednodifference inthe replicationforks indicated that the rate of DNA
lengths oftheir daughter branches, whereas
replication
was uniformthroughout
thispor-the remainder of por-the RIs had slightly different tionof the H-1 genome. The originof
replica-daughter-branch lengths, whosevariation was tion is thus within 0.15 genome units of an
approximately
that expected due to methodo-end(s)
of the DNA. No"eye"
structures werelogical errors,
determined
by measuring the seen, andreplication
was observed toproceed
distribution of RFmonomerlengths (75% of the
unidirectionally throughout
73%of the RFmol-RIshad
daughter-ann
lengths
that differedby
ecule. Of additionalsignificance
isthefinding
on November 10, 2019 by guest
http://jvi.asm.org/
720 SINGER AND RHODE J. VIROL.
4...,,
0
7 Eet mcorphs ofre' sl - U-DNA(l f-0 lt
Aure
ofi -
r
. w r 50 fmiFIG.
7Fg.3).Eletro
mtrorahmoofesRrpcaintsolatH-ib BDNA(isolatedfomgculture
atdtheparestritiv
tempera-microscopy bytheaqueousmethod, which is 60%replicated.Note that allportionsofthe moleculearefully
extended, and thatno bushesarevisible. (D)tsl ds RImonomerreplicated18%,from poolB (Fig.3B).
thatnoneofthe dimer
length
RImoleculeswasformer
method,
H-i
DNAwaspurified
by
sub-more than 50%
replicated.
Very
similardistri- stitution of[3H]BUdR
for TdRduring
morebutions of the
replication
forkposition
werethan
one round ofreplication,
andisolating
obtained with the aqueous
spreading
tech-molecules
greater
thanhybrid
density
fromnique.
Cs*SO,
gradients.
Unlike cellularDNA,
viralDISCUSSION
RF
DNAundergoes multiple
rounds ofsemi-DISCUSSION
~~~conservative
replication
during
the shortperiod
In this
study,
we examinedH-i
viral,RF,
of
density
labeling,
so thatselecting
DNA and RI DNAby
electronmicroscopy.
Viral greater thanhybrid
density
excluded 98% of DNA wasprepared
frompurified
virionsby
anycontaminating light,
cellular DNA.Simi-alkaline sucrose
gradient
centrifugation.
Gellarly,
RIDNAmolecules becomeradiolabeled,
electrophoresis
of virion DNA revealedit to beand
thepurity
ofthesepreparations
appearslargely homogeneous
inmigration
withasmallhigh
on the basis of controlexperiments
withproportion
(approximately
16%) of faster(pre-
mock-infectedcultures,
H-i-infected
cellsusing
sumably
shorter)
molecularspecies,
possibly
displacement hybridization
(13),
andhomoge-generated
by radiolysis.
RFand RI DNA wasneity
ingel
electrophoresis
before and afterextracted from infected cultures
by
the Hirt restriction endonucleasedigestion
(in
thispa-method,
sedimented in neutral sucrose gra- per). The second method ofpurification
utilizeddients,
andpurified
furtherby isopycnic
cen-the
tendency
of RI molecules to bind to BDCtrifugation
inCs2SO4,
orby
BDCchromatogra-
tightly by
virtue of their ssDNAcomponents,
phy
andisopycnic
banding
inCsCl.
With theand
theirelutability
therefrom with caffeine.on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.77.444.66.360.2]DNAs from
Cs2SO4
density gradients.Further-30 _ : more,thelengths ofH-1RF and RI DNAs have
consistently been the same (1.5
gm)
through-25 _ RF out our electron microscope analysis, and this
DNA also exhibits a
specific
pattern
ofloops
after partial denaturation; the lengths of its
20 . :
EcoRI
fragments are similarly uniform afterKleinschmidt preparation (19). These results
would not be possible if our RF and RI DNA
were heavily contaminated with random-sized
fragmentsof cellular DNA.
10- _Our results show that the RF dsDNA of tsl
H-1
replicates
viaalinearY-shaped
(branched)
RI DNAthatappearstobe
largely ds;
thesumofthe lengths of the unreplicatedportionof the
RI plus one replicated branch is equal to 1.5
E,m,
the RFlength.
The initiation site for thisz n RF replication islocatedwithin 15% of one or
JLl1RI both ends of the RFDNA;the replication fork
5_ , 8
appeared
toproceed alongmostof the RImole-cule at a uniform rate. Wedid not observe any
ss
regions
in numeroustsl H-1 RIDNAmole-07 Lo 20 30
'5 'o
culesusing
the aqueoustechnique,
whichLength Am) makes such regions appear as bushes (4). How-FIG. 8. Histograms of the lengths of linear tslH-1 ever, small ss sections may be present in
H-1
ds RF DNA molecules, and Y-shaped ds RI DNAs RIs because they were preferentially retained (length = sum of longest branch plus unreplicatedagainst
salt elution from a BDC column, base), isolated fromhamster embryofibroblasts or which separates dsDNA molecules from those human NBcells(Fig. 3B) at the restrictive tempera- csspres
dsDNA Branchedfrom Rls ture, and prepared for electron microscopy bycontaiing
ssregions (16). BranchedlinearRIs the 50%/20%formamide technique.Both RF and RI have been similarly visualized using electron populations exhibit subgroups of monomer and di- microscopy for parvovirus LU III (18), ade-merlengths having very similar variation. novirus 2 (2), adenovirus type 5 (6), herpessimplexvirus type 1 (HSV-1) (17),and the
bac-This method has proved successful for isolating teriophage T7 (24). ssDNA segments were
con-RImolecules in a variety of DNA viruses (17), spicuous in the RIs of adenovirus 5 and phage
andwaspreviously usedtopurify H-1 RI DNA
T7,
butwere notobserved inadenovirus2and(16). HSV-1although theRIs of both ofthese viruses
Itispossiblethat mock-infected culturesare werealsopurified withasimilarprocedure
us-unsuitable controls for nonviral DNA contami- ing
benzoylated naphthoylated
DEAE-cellulosenation, sincethe cytopathic effects of H-1 infec-
chromatography.
Replicative loops ("eye"tionmay causefragmentation of cellular DNA.
forms)
werefoundinHSV-1and T7 in additionWe
previously found
noevidence for
such an toY-shaped
RIs. We didnotobservereplicative
effect (13), using host cells with
prelabeled
loops
inH-1 RIDNAmolecules. This isproba-DNA. Asa further testfor cellular DNA con-
bly
due tothe location of the initiationsite-tamination,the RF and RI DNA regions (pools very close to the end of the DNA
(within
0.23Aand B)of thesucrosegradientweresubjected ,m), sothatany putative
loop
would betran-to agarose
gel electrophoresis,
and the total sient and inconspicuous; thereplication
of atDNAwasidentified by staining with ethidium least73%of the H-1genome wasobservedtobe
bromide. The
predominant
areasstainedcorre- unidirectional.However,
we can not excludespondedtothelabeled H-1monomerand dimer the possibilitythat anend of the RFmolecule
RF DNA, as determined by
autoradiography.
might
bereplicated by
asecond forkmoving
inThus, there is noevidence for significant con- a direction opposite tothat observed. The
ac-taminationof theseH-1 RFand RIDNAprepa- companyingpapers in this series (16, 19) also rations with unlabeled cellularcontaminants, present evidence
indicating
thatthe initiationunless thisputativeDNAisthe size of H-1 RF, sitefor RFreplicationislocalizednear aunique
and has anEcoRI cleavage site at the same endof theRFDNA molecule.
locationas inH-1 RF.Thisunlikelypossibility RFand RI molecules of dimerlength (3.0 ,um)
was dealt withby selecting BUdR-substituted werealso foundinourstudy. However,not one
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.501.48.239.62.309.2]722 SINGER AND RHODE J. VIROL.
raonTT ofGenome
40.(
35I
30r
25
O.
FIG. 9. Ordered distribution of replication fork positions in tsl H-i ds-RI monomer DNA molecules purified fromhumanNBcellsorhamsterembryo fibroblastsat39.50C(Fig.3B). RIs between1.0and2.1 gm long(sumofthelongestarmplustheunreplicatedbase) werenormalized andrankedaccordingtotheamount
ofreplication that had occurred. Thepositionofthereplicationfork isdistributedrandomlyalongthe H-i
genomeregionbeginning0.15fractional unitsfromoneendoftheDNA andterminating0.12unitsfromthe other end.
ofthe measured RI dimers had
replicated
more branched molecules due to chance end-to-side thanhalf of itslength.
Sincethemajority
of RFapposition
oftwo RF molecules would be pro-dimers arecomposed
ofmonomers linkedby
portionaltothe numberof molecules thatinter-hydrogen
bonding
rather thanby
covalent sected each other. Based on the resolution of bonds(16),
it islikely
that dimer RI DNAs themethod,
thewidth,
andthemeanlengths
of dissociate when thereplication
fork reaches a themolecules,
weestimate that 5%ofthe total gap at thecenter of the molecule.intersecting
molecules would appear to be One-fourth of the observedH-i RI molecules branched Rls. The number ofX-shaped
inter-had sister branches whoselengths
differedby
secting
molecules in the hamsterembryo
RF 20 to 45%, which was greater than the 20% DNApreparation
was26, so that theexpected
variation observed in RFlengths.
This differ- number of branchedY-shaped
molecules duetoence could be caused
by
eitherradiolysis
or chancewas1.3.Since 34Y-shaped
RImoleculesphotolysis
due toincorporation
oflarge
were observed in thispreparation,
it is veryamounts of [3
H]BUdR,
by
theasymmetrical
unlikely
that this artifact accounts for moredistribution of small ssDNA
regions
indaugh-
than a smallpercentage
of the H-i RI DNAs. ter arms, orby
thelarger
relativeerrorsintrin-Also,
the distribution of RI DNAlengths
wassic to
measuring
shorterlengths.
Some of the very similarto that of the RFmolecules;
thisY-shaped
DNAsweobserved couldbe artifacts would beunlikely
if asignificant
number ofresulting
from the chance association of the end branched moleculeswereduetochanceend-to-ofoneRFmolecule with the side of another RF side
overlapping.
DNA. The number of molecules
appearing
as The meanlength
ofss viral DNA extractedon November 10, 2019 by guest
http://jvi.asm.org/
[image:10.501.148.379.66.379.2]from purified wt H-1 virions and prepared in NewYork.
the presence of 30 to 50% formamide was at
5-
Edgel, M. H.,C. A. Hutchison III, andM.Sclair.1972. least0.96
to 1.0 am. The molecular weight of SpecificendonucleaseRfragmentsofbacteriophage least 0.96 to 1.0 ,um. The molecular weight of XX174deoxyribonucleic acid. J. Virol.9:574-582.this ssDNA is 1.48 x 106 to 1.56 x 106, based on 6. Ellens, D. J., J. S. Sussenbach, and H. S. Jansz. 1974.
the measured length andthe previously deter- Studiesonthe replication of adenovirus DNA. III.
mined molecularweight of 4X174ssviral DNA Electron61:427-442.microscopy of replicating DNA. Virology
(1)present as aninternal standard. The latter 7. Garon, C.F., K. W. Berry, and J. A. Rose. 1972. A
values agree closely with the figure of2.95 x unique form of terminal redundancy inadenovirus
106 obtained for themolecular weight of
H-1
ds DNA molecules. Proc. Natl. Acad. Sci. U.S.A.RF
DNA,
which wascalculated independently 69:2391-2395.using
'OX174
ds RF DNA as a reference, and 8. Johnson, P. H., and R. L. Sinsheimer. 1974. Structureusing ~X174 ds RF DNA as a reference, and of anintermediatein the replication of bacteriophage
that of 3.26 x 106, measured with respect to 4X174 deoxyribonucleic acid: the initiation site for
specific lambda phage fragments using gel elec- DNAreplication.J.Mol. Biol.83:47-61.
trophoresis
(16). It isof
interest to notethat the 9. Koczot, F., B. J. Carter, C. F. Garon, and J. A. Rose.meanlengthofH.1 ss viral DNA is signifi- 1973. Self-complementarity of terminal sequences
mean lengthz of H.-i ss viral DNA iS signii- within plus or minus strands of
adenovirus-associ-cantly shorter than that of the corresponding ds ated virus DNA. Proc. Natl. Acad. Sci. U.S.A.
RF DNA when spread under the same condi- 70:215-219.
tions using 50%formamide
(ss/ds
ratio =0.67). 10. Mayer, F., A. J. Mazaitus, and A.Puthler.
1975.Elec-tron microscopy of simian virus 40DNA
configura-The
ssDNA/dsDNA
length ratios ofa number tion under denaturationconditions. J. Virol.15:585-of DNA viruses are significantly less than unity 598.
(3, 7, 10). Finally, we did not observe the circu- 11. Pagano, J. S., and C. A. Hutchison. 1971. Small
circu-larization ofssH-1 viral DNA after incubation lar viral DNA: preparation andanalysis,p. 79-123.In
K.Maramorosch and H.Koprowski(ed.),Methods in
under annealing conditions, which occurs in virology, vol. V. Academic Press Inc., NewYork.
the caseof another parvovirus, adenovirus-as- 12. Rhode, S. L. 1973. Replication process of theparvovirus
sociated virus, and is thought to result from H-1.I. Kinetics inaparasynchronouscell system.J.
terminal
self-complementarity
(9).
Inthe pre- Virol.11:856-861.terminalself-complementarity (9). Inthepre- 13. Rhode, S. L. 1974. Replication process of the parvovirus
vious paper, evidence was presented that one H-1.II.Isolation and characterization ofH-1
replica-end of the RF DNA molecule, which is a repli- tive form DNA. J. Virol. 13:400-410.
cation terminus, is self-cohesive, andmaygive 14. Rhode, S. L. 1974.Replicationprocessof theparvovirus
rise todimer molecules in "tail-to-tail" linkage
H-1. III.
Factors affectingH-1
RF DNA synthesis.J.Virol. 14:791-801.
(16). On thebasis of this study, itappearsthat 15. Rhode, S. L. 1976. Replication process of theparvovirus
the end near the origin of replication is not H-1. V. Isolation and characterization of
tempera-complementary
to the other endasinthecase ture-sensitiveH-1mutantsdefectiveinprogenyDNAof adenovirus-associated
virus. Further evi- synthesis. J. Virol.17:659-667.16. Rhode,S. L.1977.Replicationprocess of theparvovirus
dence documenting the dissimilarity of the ter- H-1.VI.Characterizationofareplication terminusof
miniof H-1RFDNAispresentedinthe follow- H-1replicative-form DNA. J. Virol.21:694-712.
ing paper on the partial denaturation mapping 17. Shlomai, J., A. Friedman, and Y. Becker. 1976.
Repli-ofH-1 RF molecules. cative intermediatesVirology 69:647-659. of herpes simplexvirusDNA.
ACKNOWLEDGMENTS 18. Siegl, G., and M. Gautschi. 1976. Multiplication of par-vovirusLuIIIin a synchronized culture system. III. Thiswork was supported by Public Health Service grant
Repicatin
ofsviraloDNA.dJcuViro
1781-5.eCA-07826-11 from the National Cancer Institute, and a
Replication
ofviral
DNA. J.Virol.
17:841-853.CA-erous2gift
from theaiona
anc
.ins
a 19. Singer,I.I., and S.L.Rhode.1977.Replicationprocessgeeosgif from
th GieFonation.
of theparvovirus H-i.
VIII.Partial
denaturation
Wegratefullyappreciate theexcellenttechnical assist-mappingpandolocalizatonVofItheareplicationurign
anceof RobertCostantinoandJessica Bratton, and thank H-ilcative-for
DN thelictron
micros
Kay A. 0.Ellem andHelene Toolan for critically readingJH
rol.
21:724-731.this
secetaiaduie.'
manuscript, andVirginiaHaas and JaneenPratt for 2VTrol.
21:794-731.20. Toolan, H. W. 1968. The picodnaviruses: H,RV and secretarial duties. Lami, and M. S. Hopkins. 1969. Single-stranded LITERATURE CITED DNAfrom theparvovirus
H-1.
Virology39:617-621.ogy,vol. 6. Academic Press Inc., NewYork. 1. Berkowitz, S. A., and L. A. Day. 1974. Molecular 21. Toolan, H. W. 1972. The parvoviruses, p.410-425.InF.
weight of single-stranded fd bacteriophage DNA. Homburger (ed.), Progress in experimental tumor High speed equilibrium sedimentation and light scat- research, vol. 16. Kager, Basel.
tering measurements.Biochemistry 13:4824-4831. 22. Usategui-Gomez, M.,H. W.Toolan,N.Ledinko,F. Al-2. Bourgaux-Ramoisy, D., J. Robin, and P. Bourgaux. Lami,and M. S.Hopkins.1969.Single-strandedDNA
1974.ReplicatingDNAof adenovirus type2.Can. J. from theparvovirusH-1.Virology39:617-621. Biochem. 52:181-189. 23. Westmoreland,B.C.,W.Szybalski,and H. Ris. 1969. 3. Bujard, H. 1970. Electron microscopy ofsingle-stranded Mapping of deletions and substitutionsin heterodu-DNA. J.Mol.Biol. 49:125-137. plex DNA molecules ofbacteriophagelambdaby elec-4. Davis, R. W., M.Simon,and N. Davidson. 1971. Elec- tronmicroscopy.Science 163:1343-1348.
tron microscopeheteroduplex methods formapping 24. Wolfson,J.,and D. Dressler. 1972.Regionsof single-regions of base sequencehomologyinnucleicacids, p. stranded DNAinthe growing pointsofreplicating 413-428.InL.Grossman and K. Moldave(ed.),Meth- bacteriophage T7 chromosomes. Proc. Natl. Acad. odsinenzymology, vol. XXI. Academic PressInc., Sci. U.S.A.69:2682-2686.