Copyright ( 1975 AmericanSocietyforMicrobiology Printed inU.S.A.
Synthesis
of
Complex Forms of Bacteriophage iX174
Double-Stranded
DNA in a
Temperature-Sensitive
dnaC
Mutant of
Escherichia coli C
EVANGELIA G. KRANIAS1 AND LAWRENCE B. DUMAS*
DepartmentofBiochemistry and Molecular Biology, Northwestern University, Evanston, Illinois 60201 Received forpublication3March 1975
Fast-sedimenting forms of bacteriophage kX174 double-stranded replicative-form DNA observed in normal infections continued to accumulate at the nonpermissive temperature in a temperature-sensitive dnaC mutant of Esche-richia coli. These complex molecules accounted for up to half of the DNA
synthesized duringshortpulses at the nonpermissive temperature. Theywerethe
dead-end products ofDNA synthesis, not intermediates in normal
replicative-form replication. The data suggest that these
higher-than-normal-molecular-weight DNA moleculesresult from abnormal initiationof kX174replicative-form DNAreplication.
Two to four percent of the intracellular dou-ble-stranded replicative-form (RF) of bacterio-phage
OX174
DNA are multiple-length mole-cules (1, 11, 15, 16). Mostofthese are multiple-length continuous circles, although interlocked monomer circles are also observed. Both kinds ofstructures arefound to be theproduct of both DNA replication and DNA recombination. However, interlocked monomer circles are the predominant product of the DNArecombina-tion pathway, and multiple-length continuous
circles are thepredominantproductofthe DNA replication pathway (1).
Suchcomplex forms ofcircular DNAare not
unique to viruses. Multiple length forms of
bacterial plasmidDNAhave also been observed
(7, 8, 9, 10).
Wehave observed the accumulation of higher-than-normal-molecular-weight forms of
OX174
double-stranded DNA in a temperature-sensi-tive dnaC mutant of Escherichia coli at both the permissive and nonpermissive temperature forthe dnaCprotein activity. The dnaC protein isspecifically -equired for the initiation of DNA synthesis (2, 17). These complex forms ofOX174
RF DNA were also synthesized in theparental host strain, but not at the
nonpermis-sive temperature in a dnaEmutant of the same parental host. The dnaE gene product, DNA polymerase III, is essential for DNA chain elongation (5, 14). We conclude that complex
DNA molecules result fromabnormal initiation
of DNAsynthesis. The conditions under which these molecules accumulate are reported, and Present address: Department of Biochemistry. North-westernUniversity Medical School, Chicago,Ill.60611.
their relationship to the complex
OX174
RF DNA molecules observed by others in normal infections is discussed.MATERIALS AND METHODS
Bacteria and phage strains. LD301 and LD331 are,respectively,dnaEts,anddnaCtsmutants of H502 (uvrA-, thyA-, endI-) previously described (4, 12). kX174am3(gene E)is alysis-defective mutant.
Infection and preparationof cell lysates. These procedures have been described (4). In these
experi-mentsphage infection wascarriedoutinmedia
sup-plemented with 0.01
lsg
ofthymineperml. This lowconcentration of thymine was used to increase the
specific activity of [3H]thymidine-labeled
intracel-lular OX174DNA. In0.01jig ofthymineper ml
un-infected host cellsdouble in number before
multipli-cation ceases. The yield of kX174 am3 in this
concentration of thymine is 15% of that in 2 ug of
thymine per ml. The replication of OX174 am3 RF
DNA in the dnaC mutant host is as
temperature-sensitive in 0.01 ug ofthymineperml as in 2 ggper
ml.
Centrifugation analyses. The conditions of the
zone sedimentation analyses in neutral pH sucrose
have been described (12). Alkaline pH sucrose
gradi-entsconsisted of 5 to20%sucrose in 0.2MNaOH,0.8
MNaCl, 2mM EDTA, and0.1% Sarkosyl, pH 12.6.
Cesium chloride density gradient analysis was
carried out as describedbyMuller-Wecker et al. (13).
Thesampleswerespun in a Beckmantype 65 rotor at 45,000 rpm for 48 h at 20 C.
Electron microscopy. The intracellular phage
DNA wasextractedbyusing theprocedureofGodson
and Vapnek (6). The DNA was fractionated on
neutral sucrose gradients, dialyzed against 50 mM
Tris-hydrochloride, 1 mM EDTA, pH 8.1, at 4C,
and precipitated at -20C by addition of 2 to 3 volumes of ethanol.
412
on November 10, 2019 by guest
http://jvi.asm.org/
SYNTHESISOF COMPLEX
OX174
DNA 413The DNA sampleswere spread and shadowed by using thetechniquesdescribed by Davis et al. (3).
Chemicals. [Methyl-H
lthymidine,
56 Ci/mmol,was purchased fromAmersham/Searle. Mitomycin C,
chloramphenicol, egg white lysozyme, and protease (type VI) werepurchased from Sigma Chemical Co.
RESULTS
Synthesis of abnormally
high-molecular-weight
qX174
RFDNA. Previous experiments(12) showed that one of the major products of
DNA synthesis in a
4X174-infected
dnaCt8mutant host at 41 C, during the double-stranded RF DNA replication stage, was a
species that sedimented in a broad band at
about the same rate as
kX174
single-strandedDNAin high ionic strength medium (27S). We
monitored the synthesis of this DNA under
conditions where the specific activity of the
radioactive label was increased, and examined
its structure.
To determine whether this DNA was
single-stranded
phage
DNAorahigh-molecular-weight
form of
4X174
double-stranded DNA, liquidcultures of the
dnaCts
mutant LD331 wereinfected with
OX174
am3 inthe presence of 30,ug of
chlorampenicol
perml. Thisconcentrationof chloramphenicol inhibits single-stranded
synthesis,
allowing
prolonged RFreplication
(18).The
phage
DNAinthese cellswaslabeledwith
[3H]thymidine
at30and41 C. 3H-labeledDNA that sedimented faster than the
single-stranded
phage
DNAmarkerwas seeninlysates
ofthe infected cells
pulse-labeled
at30 C(Fig.
1A), and pulse-labeled after shifting to 41C
(Fig. 1B). The normal products of
OX174
RFDNA replication, RFI (21S) and RFII (16S),
were also observed. The 3H-labeled
fast-sedi-menting DNA was also observed after
longer
pulses, and after chases at 30 and 41 C (see
below).
The DNAinthe
fast-sedimenting
bandsfromsucrose
gradients
wassubjected
toequilibriumbuoyant density analysis (Fig. 2).
Almost all ofthe DNA in this band from
lysates
of cellslabeledateithertemperaturehad the
density
ofdouble-stranded DNA. The same was true of
the
fast-sedimenting
DNA fromlysates
ofcellsthat had been chased at both temperatures
(data not shown). It was not an aggregate of
normal
OX174
RF DNA held together by pro-teins since it was stable todegradation
by
nonspecific proteases followed
by
heating 20min at 56C (Fig. 1).Itwas nothost DNA since
no radioactively labeled DNA
sedimenting
atthe same rate in neutral
pH
sucrosegradients
wasfoundin extractsofuninfected host cellsat
30 or 41 C in identical experiments
(data
notshown).
We concludetherefore,
that ahigher-6
4
60-to I
a-
0z
%WOO
2
0
6
4
2
o)
-%f
10
20
30
FRACTION NUMBER
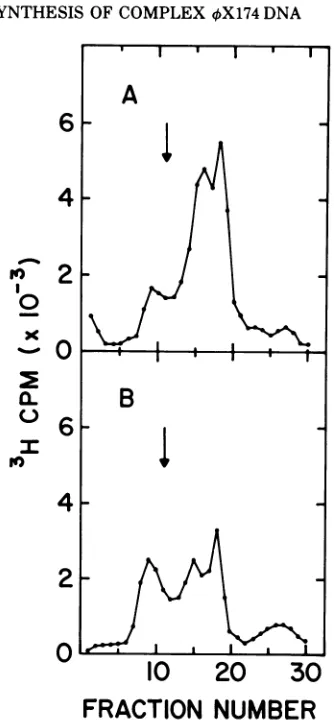
FIG. 1. Zone sedimentation of intracellular phage DNA from 4X174 am3-infected LD331. A liquid
culture of bacteria was grown at 30 C to a cell density of3 x 108 cells/ml on TPGA medium sup-plemented with 2
jug
of thymine per ml. The cells were collected by centrifugation and suspended in0.1volume of TPG medium. Mitomycin C was added
to0.1 mg/ml.After 20min in the dark the cells were collected and resuspended in 1 volume of TPGA medium supplemented with 0.01
Ag
of thymine per ml. Phage (10 per cell) and chloramphenicol (30,g
per ml) were added at zero time. After 55 min at
30 C 200MCiof [3H]thymidine were added to 20 ml of the infected culture. Two minutes later this portion of the culture (A) was rapidly chilled. At 55 min after
infection another 20-ml portion of the culture was
shifted to 41 C. Fifteen minutes later 200 ,uCi of
['H]thymidinewere addedfor 2 min, and the culture
was rapidly chilled (B). The chilled cells were col-lected, washed, and lysed. Thelysates were digested with protease, heated 20 min at 56 C, and sedi-mentedthrough neutral pH sucrose gradients (16 h at
4 C at 25,000 rpm in a Beckman SW27 rotor). Thirty fractions were collected from the bottom of each gradient. The amount of radioactivity in 0.2
ml of each fraction was measured. Arrows indicate the positions of added marker "2P-labeled virus
DNA.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.266.433.53.414.2]6
B
06
02
10
40
111,
60
-C
40
20
20
40
60
FRACTION
NUMBER
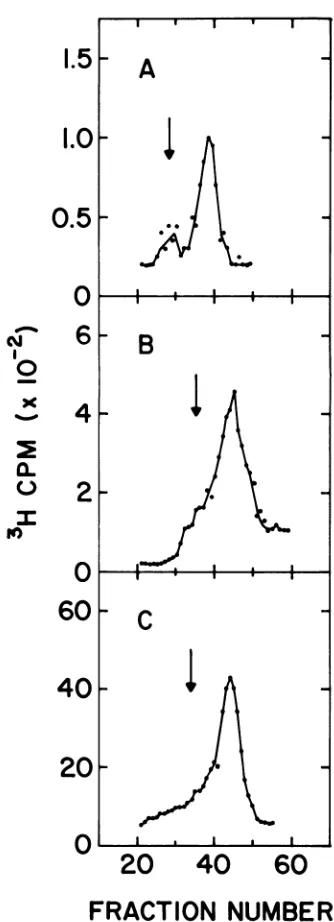
FIG. 2. Cesium chloride equilibrium buoyant den-sity analysis of the fast-sedimenting OX174 DNA.
TheDNA inthefast-sedimenting bands from neutral
pHsucrose gradients was dialyzed, concentrated by
alcohol precipitation, and spun in CsCI gradients. 32P-labeled
kX174
single-strandedDNA markerwasadded to each sample. Frame A represents the banding profile of the DNA from a lysate of kX174
am3-infected LD331 treated exactly asdescribedfor sampleA inFig. 1. FrameB represents the banding profile of the DNA from a lysate of infected cells
labeled for 30min at 41 Cbeginning at 5 minafter
the temperature shift-up. Frame C represents a
than-normal-molecular-weight form of
OX174
double-stranded DNA was synthesized in this
mutant hostboth at 30 and 41 C.
Theamount ofhigh-molecular-weight kX174
DNA synthesized in LD331duringshortpulses
at 30 Cwasaboutthe same in mediacontaining 1 and 0.01
Ag
of thymine per ml. This DNA represented 11 +3and16 ±3%, respectively,of the iX174 double-stranded DNAsynthesizedat these thymine concentrations. Similarly, thiscomplex DNA was observed in
OX174-infected
LD331 thathad notbeen treatedwith mitomy-cin C. It is not therefore an artifact of these experimental conditions.
In experiments similar to those described in Fig. 1, we measured the amounts ofthe
abnor-mally high-molecular-weight,
OX174
double-stranded DNAsynthesized at30and41 C, and the rates of synthesis of the
high-molecular-weightand normal RF DNAat 41 C relativeto
those at 30C. The data from threeexperiments
issummarizedinTable1. These data show that
the percentage of high-molecular-weight DNA
synthesized during short pulses at 30C was
about the same in LD331 and the parent host
strain H502. The rate of normal RFreplication at 41 C in LD331 was about 70% less than at 30 C. The rate ofsynthesisofthe
high-molecu-lar-weight
OX174
DNA in both hosts at 41 Cwas at least ashighasthatat 30C. Thesedata suggest that the dnaC gene product is not
TABLE 1. Synthesis of the high-molecular-weightand
normalforms ofqX174RFDNA at 41and30C in LD331 andH502a
%HMWb Rate at 41C/rate at 30 C
Expt-41 C 30 C HMW Normal Totalc
A, LD331 43 13 1.5 0.29 0.45
B, LD331 33 15 0.94 0.33 0.46
C, H502 12 16 1.2 1.7 1.6
aCellswere infected at 30C in 30
ltg
ofchloram-phenicol perml. Half ofeachculture was shifted to
41C at 45minafter infection.Equal portions of each
culture were pulse-labeled with [3H]thymidinefor 2
min(LD331)and 5 min (H502) at 20min(A) and (C)
and 30min (B) afterthetemperature shift. The rate
ofDNAsynthesiswas assumed to be proportional to
thecountsper minuteincorporated during the pulse.
bHMW, High molecular weight.
cTotal=HMWplus normal RF.
control where OX1743H-labeledRF DNA wasmixed with the 32P-labeled single-stranded DNA marker. Fractions were collected from the bottoms of the gradients. Arrows indicate the position of added
32P-labeledsingle-strandedOX174DNA marker.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.73.240.68.529.2] [image:3.503.266.457.415.507.2]SYNTHESIS OF COMPLEX OX174
essential for the synthesis of the
high-molecu-lar-weight form of /X174 DNA, although it is
forthe synthesisofnormal /X174 RF DNA (12).
Thus, at 41 C the synthesis of this complex DNA continues in the dnaC mutant host, while
the synthesis of normal RF DNA is markedly
inhibited. This results in an increase in the
percentage of high-molecular-weight DNA at 41 C.
Function of the complex forms of
4X174
DNA. We asked whether the complex forms of
iX174 double-stranded DNA were
intermedi-ates in normal RF replication or dead-end products of abnormal DNA synthesis. The
in-tracellular phage DNA in LD331 was
pulse-labeled at 41 C and chased at 30 and 41 C.
Fast-sedimentinghigh-molecular-weight kX174 DNA was again detected in the lysate ofthe
pulse-labeled culture (Fig. 3A). This complex
DNA was not chased into normal RF DNA at
either 30(Fig. 3B) or 41 C (Fig. 3C).
kX174
RFreplicationresumes at a near normal rate upon
shifting down to 30 C under these conditions
(unpublished observation). These datasuggest
thatthecomplexform of kX174DNA is not an
intermediate in normal RF replication, but
rather a dead-end product.
Synthesis of the complex form of
OX174
DNA in a dnaE mutant. In the temperature-sensitive dnaEmutantLD301fast-sedimenting
kX174
DNAwassynthesized
at30 C butnot at41 C (Fig. 4). ThissuggeststhatDNA
polymer-ase III-catalyzed chain elongation is necessary
for the synthesis of the high-molecular-weight
qX174 DNA.
Structure of the complex form of
OX174
DNA. All of our data relevanttothestructure ofcomplex form of
kX174
DNA suggest that it consists of the kinds of oligomeric forms ofOX174
monomeric RF DNAobserved byothers in infections ofnormal host cells(1, 11, 15, 16).When this DNA from sucrose
gradients
wasdialyzed, concentratedby alcohol precipitation,
and resedimented, we observed a partial
con-version to slower-sedimenting species. About
one-third still sedimented at the original rate
(approximately
30S),
while theremaindersedi-mented at 14 to30S(datanot
shown).
Previousstudiesshowed that
supercoiled
circular dimersof
OX174
RF DNA sediment at 29S, relaxedcirculardimers at21S,andhigherorderrelaxed
I0
x gO
0
10
20
30
FRACTION NUMBER
Fig. 1. At 40 min after infection 10-ml portions of the culture wereshifted to 41 C. At20minafter the
shift 100
ACi
of[(H]thymidine
were added to each. Five minutes later one portion was rapidly chilled (A), while the other two were filtered. The lattertwo were then washed with 10 ml of medium
con-taining 200jAg of thymine, 2 mgof thymidine, and
30 lAg of chloramphenicol per ml. The cells were
resuspended in 10 ml of the same medium and incubated 30 min at 30(B) and41 C(C). The cells
werecollected, washed,lysed,digested with protease, heated to 56 C for 20 min, and sedirrtented as
described inFig. 1.Arrows indicate thepositions of
added marker 32P-labeled 4X174 single-stranded
DNA. FIG. 3. Zone sedimentation of phage DNA
ex-tracted from 4,X174 am3-infected LD331
pulse-la-beled and chased during RF replication. The
con-ditionsfor culturingthecells,mitomycinC treatment, and infection were identical to those described in
1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.265.430.71.509.2]1
2
D
6
4
2
0
10 20 20
FRACTION NUMBER
FIG. 4. Zone sedimentation of phage DNA
ex-tracted from OX174 am3-infected LD301
pulse-la-beled during RF replication. The conditions were
identical to those described in Fig. 1. After the mitomycin C treatment the cellswere suspended in
I volume ofTPGA mediumsupplemented with 0.01 ,ug of thymineperml. Phagewereaddedata
multi-plicity of 10 and chloramphenicol at 30 gg per ml.
After45 minat30 Chalf ofthe culturewasshifted
to 41 C. At 20 and 45 min after the shift 10-ml
portions of the 30 and 41 C cultures were
pulse-labeled for 2 min with [3H]thymidine, 10 uCi per
ml. A, 20 min, 30 C; B, 20 min, 41 C; C, 45 min, 30C;D,45min,41C.
oligomers somewhat faster than 21S (11). Su-percoiled and relaxed monomeric RF DNA molecules sediment at21and 16S, respectively. Thus, before these manipulations the complex form of
OX174
DNA observed in the dnaCmutant host sedimented at a rate expected of
supercoiled dimeric and higher oligomeric forms ofRF DNA. After these manipulationsa hetero-geneous population of species sedimenting at
the ratesexpected of relaxed forms and
break-downproducts of these complex DNA molecules
was observed.
Zone sedimentation of the dialyzed,
concen-trated DNA from neutral pH sucrosegradients
in alkaline pH sucrose gradients showed three majorbands (Fig. 5). The DNA in the middle
band sedimented at the rate of denatured
monomeric RFI DNA. The DNA in the band
closesttothe bottomofthegradient sedimented
at the rate expected of denatured oligomeric
RFI DNA. The DNA in theslowest-sedimenting
band consisted of denatured single strands
derived from relaxed double-stranded DNA
molecules. When sedimented for longer times
(data not shown), the band was shown to be
more heterogeneous than that of an added
single-stranded circularDNAmarker. The data
suggested the presence of monomeric circular
and linear DNA strands, as well as oligomeric
single-strands.
When other preparations ofDNAfrom
neu-tral pH sucrose gradientswere spun to equilib-rium in CsCl gradients containing ethidium
bromide (100 ,ug/ml), about20% bandedat the
position ofsupercoiled DNA.
When dialyzed, concentrated DNA from
neu-tral pHsucrosegradientswasspread and
exam-8
2
6-0. 4
0
10
30
50
FRACTION NUMBER
FIG. 5. Zonesedimentation of the
high-molecular-weight kX174DNA in an alkaline sucrose gradient. LD331 was treated as described for sample B in Fig. 1. At 5 min after the shift to 41 C 200MCi of
['H]thymidine were added. Thirty minutes later the infected cells were collected and treated as described
inFig. 1. The DNA from the fast-sedimenting band
was dialyzed and concentrated by alcohol precipita-tion. This DNA sample was spun in an alkaline pH
sucrosegradient for 3.5hat 38,000 rpm in a Beckman
SW40 rotor at 5 C. Total radioactivity was measured
in each fraction collected from the bottom of the gradient.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.61.256.55.381.2] [image:5.503.290.429.321.541.2]16, 1975 SYNTHESIS OF COMPLEXOX174DNA
V
m
wr ti-N .t P,
PR
0-R, S-4W.A j;;.,Z,'.?16 slil.-o-W
VWI MW lb-Ij-'riAl 6i
WL ZP'Nou,
r-2-5.,.l.tu
J,?j5
kewO
icik
vyk
LIP *1 A6. 11-116
'IrP
Wr At.Z.401"ra.4
ir
64 VP
15c VAT
VPlfo.;v
Aar*pf'Lf; 4
%(4q *,A, '12
16 LA
[image:6.503.56.432.68.620.2]-No,
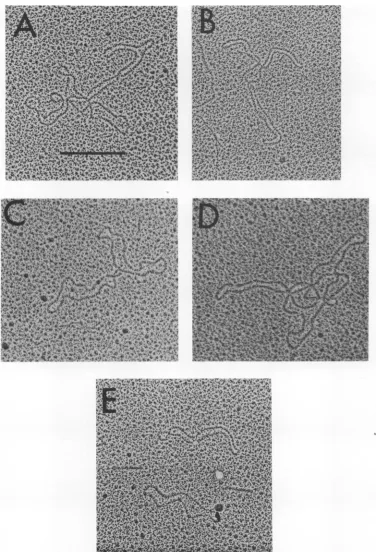
FIG. 6. Electronmicrographs of
OX1
74DNAfromLD331.A,Continuous tetramer;B,continuoustrimer;C,continuousdimer; D,trimer,probablycatenated;F,monomers.The barrepresents0.51gm.
on November 10, 2019 by guest
http://jvi.asm.org/
ined by electron microscopy, monomer and complex forms of
OX174
DNA were observed. The complex formsincludedtetramers, trimers,anddimers, some continuous, someapparently
catenated. Someexamplesareshown inFig.6.
These data suggest no unique structure for
the complex qX174DNAobservedinthednaC mutant host. The data indicate that these complex DNA molecules are similar in structure to at leastsome of those found by other inves-tigators in
OX174
infections of normal cells.DISCUSSION
High-molecular-weight double-stranded forms of
OX174
DNA were synthesized duringtheperiodofRFreplication (stage
II)
atboth 30and 41C in a temperature-sensitive dnaC
mu-tant host. dnaC mutants are defective in the
initiation ofDNAsynthesis (2, 17). At41Cthe rate of
OX174
RF DNAsynthesisinthismutant hostwas 30% ofthat at 30C (Table 1). Thehigh-molecular-weightDNA wassynthesizedat least
asfastat 41 C asat30 C. Thissuggests that the formation of these complex DNA structures is
not dependent upon the activity of the dnaC
initiation protein.
Our data suggest that this complex DNA is
the productofDNAsynthesis, rather than DNA
recombination.Itsformation required the
activ-ity of the dnaE protein, DNA polymerase III (Fig. 4). This enzyme is essential for normal
OX174
RF replication (4). Also, moreradioac-tively labeled high-molecular-weight
OX174
DNA was made at 41 C during a short pulse than at 30C, while less radioactively labeled RF DNA was made (Fig. 1, Table 1). If the
radioactivity labeled complex DNA were a
product of recombination between normal RF DNA molecules synthesized during the period
ofthepulse, the rate of its formationshould
de-crease at 41C.
The high-molecular-weight 4X174 DNA was
not chased into
kX174
RF I and RF II DNAmolecules, the products of normal RF
replica-tion, at 30 or 41 C (Fig. 3). This indicates that this complex DNA is a dead-end product of abnormal DNA synthesis, not an intermediate in normalRF replication.
High-molecular-weight forms of
OX174
dou-ble-stranded DNA have been observed
previ-ously in normal infections, and in infections where RF replication was inhibited by phage mutations and drugs (1, 11, 15, 16). Both catenatedandcontinuous circles wereobserved. Thesestructuresapparently canarise from both DNA replication and DNA recombination.
Closed circularcomplex DNA structures
repre-sent 2 to 4% of the supercoiled RF DNA
synthesized under these conditions (11). Those
complex
DNA molecules stable torepeated
centrifugation represent about 4% of the total
double-stranded RF DNA in the cell
(1).
In our experiments the complexDNA
mole-cules represented approximately 15% of the total
OX174
RF DNA synthesized at 30 C.Twenty to fifty percent of this complex DNA
was completely closed circular, as seen by
analysisin alkalinepHsucrosegradientsand in
CsCl-ethidium bromide gradients. About
one-third still sedimented at 30S at neutral pH after the initial purification on sucrose
gradi-ents.The levels ofcomplexkX174 DNA thatwe
observed in infections at 30 C are therefore
comparabletothose observed inprevious
inves-tigations, the difference being that we did not
select only the completely closed circular DNA fraction or only the fraction stable torepeated centrifugation.
Thus the kinds of DNA molecules that accu-mulate in thednaC mutant host at the nonper-missive temperaturehave also been observed in this host at the permissive temperature and in normal host cells. Thesehigh-molecular-weight
OX174
DNA molecules apparently result from abnormal initiation of RF DNAsynthesissince theycontinue to be synthesized at a normalrateunder conditions where the initiation ofnormal RF replication is inhibited. The abnormal initi-ationpathway is usedinfrequentlyin normal in-fections since only a small amount of this complex DNA is synthesized. This pathway predominates only when the normal one is inhibited, leading to an accumulation of
com-plex qX174DNA.
ACKNOWLEDGMENTS
These results are drawn from athesissubmitted byE. G. K. to Northwestern Universityin partialfulfillment ofthe requirementforthe Ph.D. degree. We thank R. Ryanforhis helpwith theelectron microscopy, andH. Swift for theuse of hiselectron microscope facilities.
This work was supported by a Public Health Service research grant (AI-9882) and research careerdevelopment award(AI-70,632) to L. B. D. fromthe National Instituteof AllergyandInfectiousDiseases.
LITERATURE CITED
1. Benbow, R. M., M. Eisenberg, and R. L. Sinsheimer. 1972.MultiplelengthDNAmoleculesofbacteriophage OX174.Nature(London)New Biol. 237:141-144. 2. Carl, P. L. 1970. Escherichia coli mutants with
tempera-ture-sensitive synthesis ofDNA. Mol. Gen. Genet. 109:107-122.
3. Davis,R.W.,M.Simon, andN.Davidson. 1971. Electron microscope heteroduplex methodsformapping regions ofbasesequencehomologyinnucleic acids,p.413-428. In L. Grossman and K. Moldave (ed.), Methods in enzymology, vol. XXI. Academic Press Inc., New York.
on November 10, 2019 by guest
http://jvi.asm.org/
SYNTHESIS OF COMPLEX
OX174
DNA 4194. Dumas, L. B., and C. A. Miller. 1973. Replication of bacteriophage 4X174 DNA in a temperature-sensitive dnaE mutant of Eseherichia coli C. J. Virol. 11:848-855.
5. Gefter, M. L., Y. Hirota, T. Kornberg, J. A. Wechsler, andC. Barnoux. 1971.Analysisof DNApolymerasesII and m in mutants ofEscherichia colithermosensitive
for DNA synthesis. Proc. Natl. Acad. Sci. U.S.A.
68:3150-3153.
6. Godson, G.N.,and D. Vapnek. 1973. A simple method of
preparing large amounts of OX174 RF supercoiled DNA. Biochim.Biophys.Acta 299:516-520.
7. Goebel, W. 1970. Studies on extrachromosomal DNA.
Replication ofthe colicinogenic factor colEl in two
temperature-sensitive mutants of Escherichia coli de-fective in DNA replication. Eur. J. Biochem. 15:311-320.
8. Goebel, W. 1974. Studies on the initiation of plasmid DNA replication. Eur. J. Biochem. 41:51-62.
9. Goebel, W., and D. R. Helinski. 1968. Generation of
highermultiple circular DNA forms inbacteria.Proc.
Natl.Acad. Sci.U.S.A.61:1406-1413.
10. Goebel, W., and J. Kraft. 1974. Complex col El DNA in Escherichia coli and Proteus mirabilis. Mol. Gen.
Genet. 129:149-166.
11. Gordon, C. M., M. G. Rush, and A. C.Warner. 1970.
Complexreplicative form molecules ofbacteriophage
*X174and S13su105.J.Mol. Biol. 47:495-503.
12. Kranias, E. G., and L. B. Dumas. 1974. Replication of bacteriophageOX174DNA in atemperature-sensitive dnaC mutant of Escherichia coli C. J. Virol. 13: 146-154.
13. Muller-Wecker, H., K. Geider, and H. Hoffmann-Berl-ing. 1972. DNA synthesis in nucleotide-permeable Escherichia coli cells. IV. Mode of1X174 replicative form DNAsynthesis and the templateinvolved. J. Mol. Biol. 69:319-331.
14. Nusslein, V., B. Otto, F. Bonhoeffer, and H. Schaller. 1971.Function of DNA polymerase III in DNA replica-tion. Nature(London) 234:285-286.
15. Rush, M. G., and R. C. Warner. 1968. Multiple length rings ofOX174and S13 replicative forms.III. A possible intermediate in recombination. J. Biol. Chem. 243: 4821-4826.
16. Rush, M. G., A. K. Kleinschmidt, W. Hellman, and R. C. Warner. 1967. Multiple length rings in preparation of
4X174replicativeform. Proc. Natl. Acad. Sci. U.S.A.
58:1676-1683.
17. Schulbach, W. H., J. D. Whitmer, and C. I. Davern. 1973.
Geneticcontrol of DNA initiation in Escherichia coli.
J. Mol. Biol. 74:205-221.
18. Sinsheimer, R. L., B. Starman, C. Nagler, and S. Guthrie. 1962. The process of infection with bacterio-phageOX174.I.Evidence for areplicative form. J. Mol. Biol. 4:142-160.
VOL. 16,1975