0022-538X/81/040151-06$02.00/0 1
Characterization and
Classification
of Virus
Particles
Associated with
Hepatitis
A
III.
Structural
Proteins
JONDURI TRATSCHIN,' GUNTERSIEGL,'* GERTG. FROSNER,2AND FRIEDRICH DEINHARDT2
Institute of Hygiene and Medical Microbiology, University ofBern, CH-3010 Bern, Switzerland,1and Max vonPettenkofer-Institute of Hygiene andMedical Microbiology, D-8000Munich2,FederalRepublic of
Germany
Received 16 September 1980/Accepted22December1980
Hepatitis A viruswaspurified fromfecal samplescollectedatvarioustimes in
the incubationperiod of patients withnaturallyacquired hepatitisA. The proteins
ofparticles bandingataround 1.34 g/ml inCsClandsedimentingatabout 160S
wereradioiodinatedin vitro and separated by electrophoresisonpolyacrylamide
gels in thepresence of0.1%sodium dodecyl sulfate and8Murea. Under these
conditions, the capsid proteins resolved into four polypeptides with molecular weights ofapproximately 31,000, 24,500, 21,000, and9,000, respectively. A fifth protein of about 40,000 daltons in size and assumed to be equivalent to the
precursor polypeptide VPO of the picornaviruses was presentin particles
sedi-menting at only 150 to 155S and banding at around 1.33 g/ml in CsCl. The physicochemical characteristics ofthese particlesareconsistent with those of the provirion structures of picornaviruses. In several of the fecal samples, these particles representedaconsiderable fractionof all particlespresent. The signifi-cance of this finding with respect to the antigenicity of hepatitis A antigen extracted from stool specimens isdiscussed.
HepatitisAvirus (HAV) hasbeen identified repeatedly in samples of feces, bile, and liver collectedfrompatientsinanepidemic of hepa-titisorfrom
volunteers,
chimpanzees,
andmar-mosets in controlled experimentalstudies. The virus particles measured 27 nm in
diameter,
bandedataround 1.34
g/ml
inCsCl,
and sedi-mentedatabout 160S(3,7,
16-18).Furthernore,
thereis evidencetosuggest thatthegenome of HAV consists ofsingle-strandedRNA (8, 13, 17, 19). These physicochemical characteristics of thepurifiedparticlesstronglysupport the clas-sificationofHAVwith thepicornaviruses.
Intactpicornavirus
particles
containfour well-definedproteins.
Theavailable dataonthepoly-peptide spectrum of
HAV,
however,
areonly
partially in agreement with this
requirement.
Three
polypeptides
in the size range ofpicor-navirus structural
proteins
VP1 to VP3 wereregularly detected (5, 8), but only very recent experimentsrevealedtraceamountsof afourth, minor
polypeptide
migrating
in theappropriate position ofVP4 (6). Onthe other hand, Fein-stone et al. (8)reported
the presence of anadditional virus-associatedproteinwitha molec-ularweightof59,000.The size of thisproteinis muchlargerthan thatof theprecursor
polypep-tide VP0 (38 to 44 kilodaltons
[kD])
which ispresent inimmaturepicornavirus capsids. Here weprovide evidencethat HAVparticles collectedatvarious times inthe incubation pe-riodofpatientswithnaturallyacquiredhepatitis Ashowaproteinspectrumcompatiblewith that of knownpicornaviruses.Our results alsosuggest that variations in the protein pattern can be relatedtothe relative concentrations of imma-ture, provirion-like structures andmature viri-onsintheHAV
samples
underinvestigation.
MATERIALS AND METHODS
Origin and purification of viruses. The fecal specimens used as a source of HAV were collected fromanindividual involved inanepidemicofhepatitis
AinGermany (sampleno. 1) (9) and frompatientno.
2, whocontracted the infectionduringholidaysinthe Mediterraneanarea.Acompleteseries offecalsamples
from patient no. 2 was available from days -9 to 4
(day0wastheonset ofdisease) (8a).Samplesno.2 11/5,no. 2 11/6,andno. 211/9, collected 5, 4,and 1
daybefore theonsetofdisease,respectively,wereused asasource of HAV particles. Stool ofpatientno. 1 was obtained on the 6th day before the onset of jaundice. Theserological and physicochemical char-acteristicsofparticlesfromthe lattermaterial,aswell as the methodused for purification, have been de-scribed indetailpreviously (18, 19).
All stepsin the purification andconcentration of
151
on November 10, 2019 by guest
http://jvi.asm.org/
Radioiodination of viralproteins. Forlabeling
of virionproteins with theluI-Bolton-Hunterreagent (1), virusparticlessuspended in0.1Msodiumborate (pH8.4)-imMEDTAweredisruptedin the presence of 1% sodiumdodecylsulfate (SDS) by boilingfor1 min. Samplesof50 to 100
Id
ofthe SDS-complexedpolypeptides were then iodinated with100 to 200,uCi
of the "nI-labeledBolton-Hunter reagent ina silicon-ized glass tube at 00C for 90 min under standard conditions(1). Labeledproteinswereseparated from low-molecular-weight componentsby gelfiltrationon a Sephadex G-10 column equilibrated with elution buffer(50 mMTris-hydrochloride [pH 6.8],0.2%
gel-atin) and prerun with 0.5 ml of a 10% solution of bovineserumalbumin.
For iodination of proteins by the chloramine-T method, virusparticles were dialyzed against 50 mM sodiumphosphate (pH 7.5) beforedisruption by boil-ing. Iodination under the conditions given by Carthew and Martin (4) was allowed to take place at room temperature for 1 min, and labeled proteins were separated from unreacted iodide by gel filtration as described above.
The"25I-labeledBolton-Hunter reagent (specific ac-tivity, 2,000 Ci/mmol) and ['251]iodide (carrier-free) wereobtained from the RadiochemicalCentre,
Amer-sham, England.
Gelelectrophoresis.Samplesoflabeled HAV pro-teins werepreparedin theappropriatesamplebuffers ofthe gel systems described below and, in general, wereheated to 90°C for 15 min beforeloading onto thegels. Three types of gel and buffer system were used forelectrophoresis:(i)SDS-polyacrylamidegels inthe discontinuousbuffer systemdescribed by La-emmli(12); (ii)SDS-polyacrylamide gels in the same discontinuous buffer systemsupplemented with either 1or 4M ureaby the method of Bowen et al. (2); and (iii)amodification of the continuous phosphate buffer systemcontaining8Murea asdescribed by Summers etal. (21). In the latter system, both the gel and the
runningbuffer contained 0.1 Msodium phosphate (pH
8.0)-0.1% SDS-8 M urea. Protein samples were pre-paredinthesame bufferplus 1%
,f-mercaptoethanol,
2%SDS, 8 M urea, 20% sucrose, and 0.002% bromo-phenol blue. Electrophoresis in discontinuous gels was carriedoutinslab gels (120 by 130 by 1.5 mm) at 80 V pergel, whereas electrophoresis in continuous phos-phate-urea gels was performed in glass tubes (5 by 130 mm)at aconstantcurrent of7 mA pertube. Proteins were fixed and stained in 1.5% Coomassie blue-1% amidoblack-45% ethanol-9% acetic acid and destained in 15% ethanol-7.5% acetic acid. Gels were finallysliced into 1-mmsections, and the radioactivity was determined withoutfurther treatment of the slices in aPackard PRIASAuto-Gamma counter.
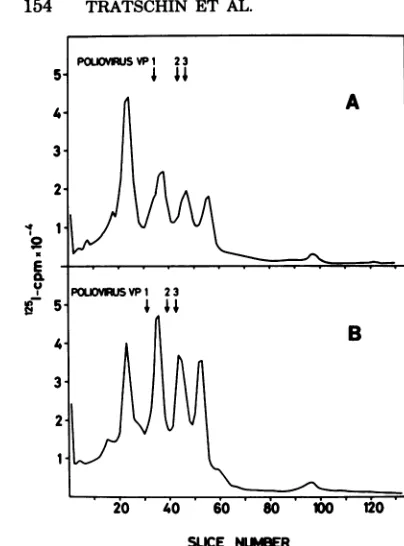
tions, only particles purified from stool no. 1 gave rise to a polypeptide spectrum that was more orless consistentwiththatof the marker
picornavirus (Fig. 1). It consisted of four poly-peptides tentatively labeled VP1 to VP4 and about35, 31, 26, and 7 kD in size. Moreover, two additionalpolypeptides with apparent molecular weights of 45,000 and 48,000 could be distin-guished. The latter polypeptides were also pres-ent inall HAVpreparations purified from fecal specimens of patient no. 2. In general, they con-stituted themajority of labeled proteins in these samples, whereasthe polypeptides migrating in thepositions of polioviruspolypeptides VP1 and VP4could barely be detected or were not
pres-ent atall.
A variation in several experimental param-eters (e.g., the technique usedfor
radioiodina-tion, treatmentofsamplesbefore electrophore-sis, origin and concentration ofacrylamide,and
addition of 1 and4 M urea to the discontinuous
gel system) hadnosignificanteffect onthe qual-itative andquantitative aspectsofthe polypep-tide pattem of individual samples. However, electrophoresisin acontinuousphosphatebuffer system containing 0.1% SDS and 8M urea
re-sulted in thefollowingdrasticchanges (Fig. 2). Besides ahigh-molecular-weight peak,three ma-jor proteinsand afourth minorproteincould be detected regularly in all HAV samples. All of thesepolypeptides appearedtobesmaller than the respective structural proteins ofpoliovirus (Table 1). Theonlyhigh-molecular-weight poly-peptideresolved in thepresence of 8 M ureahad an apparent molecularweight ofabout 40,000. This is clearlywithin thesize rangeof the pre-cursorpolypeptideVPO ofwell-characterized pi-comaviruses.
PolypeptideVPOofpicomavirusesis present insignificant quantities onlyin"empty" capsids or in immature virusparticles. Thesetypes of particle differ from the mature virion in their buoyancy andsedimentation behavior (14). As buoyantdensity centrifugationofaliquots ofthe HAV samples in shallow CsCl gradients indi-cated (Fig. 3), only particles purifiedfromstool no. 1and knowntobealmost devoid ofthe VPO-likepolypeptidehad abuoyancy behavior com-parableto that ofpoliovirus; yetpeak
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURAL PROTEINS OF HAV
m0 x
E Ur
In
SUCE NUMBER
FIG. 1. SDS-polyacrylamidegel electrophoresis of the capsid proteins ofpoliovirustype2(A) and of HA V particles extractedfromstoolsamplesno.1(B),no.211/5 (C), and no. 211/6 (D). The purified viruses were disrupted in the presenceof1%SDSby heatingto100°C for 1 min before radioiodination with the Bolton-Hunter reagent.Labeledproteinswereagain heatedto100°Cfor2min(1%SDS,2.5%
,8-mercaptoethanol),
and
polypeptides
wereseparated byelectrophoresisinadiscontinuous 15%SDS-polyacrylamide slab gel by the methodofLaemmli(12).Arrows indicate thepositions ofpolypeptidesVPOtoVP4ofpoliovirus.trations ofhepatitis A antigen accumulatedata
slightly lower density (1.336 g/ml) than did the marker picornavirus (1.34 g/ml). In contrast,
virussamples extractedfrom the fecal specimens of patient no. 2 and containing the VPO-like protein in considerable quantity regularly bandedatdensities of below 1.336 g/ml. Insome of these samples, there wasalso evidence that suggested thepresenceofmorethanoneparticle
species (e.g.,no.211/6inFig. 3). Sedimentation studies yielded consistent results (data not
shown indetail).Particles extracted from sample no. 1sedimented at 157S,whereas 150to 154S wasrecorded for HAVcontained in the stools of patientno.2.
DISCUSSION
The resultsreported in this paper arein
ac-cordancewith the observations of Coulepisetal.
(5) and Feinstone et al. (8) in as far as they
confirm the existence of three major protein
components in HAV capsids with molecular weights similarto those of the structural
poly-peptides
VP1 to VP3 ofpicomaviruses.
More-over,there isevidencetosuggestthat theHAV particles also containaminorpolypeptide com-ponentmigrating
intheposition
ofpicomavirus
polypeptide
VP4, as well as ahigh-molecular-weight
protein
comparable
insizetothe precur-sorpolypeptide
VPO. Theconcentration ofthe latterproteins,
however, varied withtheorigin
of thevirus
particles.
Rigorous
experimental
conditionsproved
tobenecessaryfor
resolving
thepolypeptide
spec-trum of HAV in a reliable andreproducible
manner. At least one of the
high-molecular-weight
peaks
of the spectrumdisplayed
in dis-continuousgel
systemsdisappeared
upon elec-trophoresis in the presence of 0.1% SDS-8 M urea, and radiolabeled material lost from that regionapparently
addedtotheconcentrationofpolypeptides
VP1toVP4(e.g.,
no.211/6
inFig.
1 and 2).
Therefore,
it may be concludedthat theformation
ofaggregatesofone or morestruc-tural
polypeptides
ratherthancontaminationof virussamples
with unrelated orvirus-specific
VOL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.494.46.447.58.348.2]
w-E
a.
SUCE NUMBER
FIG. 2. Polypeptides ofHAVparticles extracted fromstoolsamplesno.211/6(A) andno.211/9(B)
as revealed by electrophoresis incontinuous 12.5%
polyacrylamidetubegels containing8Murea.HAV
particlesweredisruptedasdescribedinthelegendto
Fig. 1, radioiodinated by the chloramine-T
proce-dure,andheatedto90'Cin thepresenceof2% SDS-1%f3-mercaptoethanol-8Mureafor15min.Arrows indicatethepositions ofunlabeledpoliovirus poly-peptides coelectrophoresed in the same tubes and
locatedbyproteinstaining.
butnonstructuralproteins (e.g., the equivalents of the 52- and 57-kDproteins detected in foot-and-mouth disease [15] or Mengo virus [22] particles, respectively) might be responsible for the appearance ofhigh-molecular-weight com-ponentsintheprotein pattem of HAVas occa-sionally revealed in Laemmli gels (8).
Ifwe assume thatHAV is atypical picorna-virus, it is obvioustorelate the 40-kD polypep-tideresolved in SDS-ureagelstotheprecursor polypeptide VPO. This protein is known to be
present inprocapsids (empty capsids) of picor-naviruses and, in the process of maturation of thevirus, is cleaved intoVP2 and VP4 (14). The onlytype of particle in which VPO, VP2, and VP4 have been found simultaneously and in
various concentrations isanimmediate precur-sor, the provirion, ofmature enterovirus parti-cles (10, 11, 20). Provirions contain a full com-plement of viral RNA, sedimentat130to150S,
VP4 9,000±2,000
aTentative classification.
b Polioviruspolypeptides
(VP0,
41kD;VP1,32kD;VP2, 28 kD; VP3, 25 kD; VP4, 8kD) were used as
internalmarkers and detectedby stainingwith Coo-massie blue. Additional molecular weight markers werebovineserumalbumin(68kD),gammaglobulin,
H-chain(50kD), ovalbumin(43 kD),pepsin (35kD), chymotrypsinogen (25.7 kD), papain (23 kD), cyto-chrome c (11.7 kD), and insulin B chain (3.4 kD). Numbers representmeanvalues ofseven determina-tions.
and band atlower densities inCsCl than does the mature virion. The analogies between the proteinpattern and the buoyancy and sedimen-tationcharacteristics ofthese structures and the HAVparticlesunderinvestigationarestriking. Therefore,it isconceivable that the 40-kD pro-tein indeedrepresentstheequivalent of the pre-cursor
polypeptide
VPO. At the same time it must be concluded that themajority
of HAV particlespresent in the fecalsamples
collected frompatientno. 2 areprovirionstructures.Since provirions of picornaviruses probably are not infectious (11), the concentration of physical particles allows no reliable conclusion to be drawn about the infectivity of a certain stool sample. This pointmustbeconsidered when the many unsuccessful attempts to infect cell cul-tures with HAV-containing fecal extracts are evaluated.Finally, itiswellknown thatmature
picorna-viruscapsidscarry acharacteristic immunolog-icalreactivity definedasD (orN) antigen. The absence or loss ofVP4 from these particles is
equivalent
totheappearanceofanentirelynew setofsurfacedeterminants,theC(or H) antigen (14). Theantigenicityof provirionparticleshas not been determined. In light of our protein data, however,it istemptingtospeculateon the extenttowhichthe use of uncharacterized fecal extracts as asourceofhepatitisAantigen could affectthesignificance and reproducibility of the available serological tests for HAV anti-bodies.ACKNOWLEDGMENTS
We thank Giovanna CattaneoandDanielWasem for ex-cellent technical assistance.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.494.45.247.62.335.2]OF HAV
0
x
a.
u I
5
0
5
4 X
K
3
YI
2
FRACTIONS
FIG. 3. Buoyantdensity centrifugation of mature poliovirions (A) and HAVparticles (0)purifiedfrom
stoolsamples no. 1(A), no.211/5 (B),and no. 211/6(C) inCsCL.[3H]uridine-labeled poliovirus(10
fi)
and unlabeled HAVparticles (50id)
weremixed with 7.9mlof CsCI (p= 1.34g/ml;made up in 50 mMTris-hydrochloride[pH7.41-0.1% Sarkosyl NL97)andcentrifugedtoequilibrium in a Beckman 50 Ti fixed-angle rotor at40,000 rpm at 15°C for 60 h. A total of 63fractionswere collectedby bottom puncture of the tubes.
Samples of each fraction were assayed for3Hradioactivity by liquid scintillation counting and for the presence of hepatitis A antigen in theradioimmunoassay. Poliovirus was assumed to band exactly at a
density of1.34g/ml.
This studywas supportedby grant 3.945-0.78 from the Schweizer Nationalfonds and grant De 261/1 from the DeutscheForschungsgemeinschaft.
LITERATURE CITED
1. Bolton,A.E.,and W.M.Hunter.1973.Thelabellingof proteinstohigh specificradioactivityby conjugation to
a"2'I-containingacylatingagent.Biochem. J. 133:529-538.
2. Bowen, B., J. Steinberg, U. K. Laemmli, and H. Weintraub. 1980. The detection ofDNA-binding pro-teinsby proteinblotting.Nucleic Acids Res. 8:1-20. 3. Bradley,D.W.,C.L.Hornbeck,E. H.Cook,and J. E.
Maynard.1977.Purification ofhepatitisAvirus from chimpanzeestools. J. Virol. 22:228-231.
4. Carthew, P.,and S. J. Martin.1974.The iodination of bovine enterovirusparticles.J.Gen.Virol. 24:525-534. 5. Coulepis,A.G.,S. A.Locarnini,A. A.Ferris,N. I.
Lehmann, andL.D.Gust.1978.Thepolypeptidesof hepatitisA virus.Intervirology10:24-31.
6. Coulepis,A.G.,S. A.Locarnini,andL.D.Gust.1980.
Iodination ofhepatitisAvirusrevealsafourth
struc-turalpolypeptide.J.Virol.35:572-574.
7.Feinstone, S. M., A. Z.Kapikian,and R. H.Purcell. 1973. HepatitisA:detection by immune electron
mi-croscopy ofaviruslikeantigenassociated with acute
illness. Science 182:1026-1028.
8. Feinstone, S. M., Y.Moritsugu,J. W. K.Shih,J.L. Gerin, and R. H. Purcell.1979. Characterization of hepatitisAvirus,p.41-48.InG.N.Vyas,S. N.Cohen, and R. Schmid (ed.), Viral hepatitis. Abacus Press, Turnbridge Wells,Kent.
8a.Frdsner, G. G., F. Deinhardt, M. Roggendorf, R.
Scheid,B. Bayerl,and R. Zachoval. 1980. Labora-torydiagnosisofhepatitisAinfection,p.17-26.InL.
Bianchi, W.Gerok, K. Sickinger, and G. A. Stalder (ed.), Virus and theliver. M. T. P. Press, Lancaster, England.
9. Fr6sner, G. G., L R. Overby, B. Flehming, H. J. Gerth, H.Haas, R. H. Decker, C. M. Ling, A. J. Zuckerman, and H. R. Frosner. 1977. Seroepide-miologicalinvestigation of patients andfamily contacts inanepidemic of hepatitis A. J. Med. Virol. 1:163-173.
10. Guttman, N., and D. Baltimore. 1977. Morphogenesis ofpoliovirus.IV.Existence ofparticlessedimentingat 150S andhavingthe properties of provirion. J. Virol. 23:363-367.
11. Hoey, E.ML,and S. J. Martin.1974. Apossible precursor containingRNAof a bovine enterovirus, the provirion II.J. Gen. Virol. 24:515-524.
12.Laemmli, U. K. 1970. Cleavage of structuralproteins duringtheassemblyof the head ofbacteriophageT4. Nature(London)227:680-685.
13.Provost, P.J., B.S. Wolanski,W. J.Miller, 0. L Ittensohn, W. J. McAleer, and M. R. Hilleman.
1975.Physical, chemical andmorphologicdimensions of humanhepatitisAvirusstrain CR326.Proc. Soc. Exp.Biol. Med. 148:532-539.
14.Rueckert,R.R.1976.Onthestructureand morphogen-esis ofpicornaviruses,p. 131-213. In H.Fraenkel-Conrat and R R.Wagner (ed.), Comprehensive virology, vol.6.
PlenumPublishing Corp., New York.
15. Sangar,D.V., D. J.Rowlands,D.Cavanagh,and F. Brown. 1976.Characterization of the minor polypep-tides in the foot-and-mouth disease particle.J. Gen. Virol. 31:35-46.
16. Schulman,A.N.,J.L.Dienstag,D.R.Jackson,J. H.
Hooflagel, R. J.Gerety,R.H.Purcell,and L F. Barker. 1976. Hepatitis Aantigeninliver, bile, and stool ofchimpanzees.J.Infect. Dis.134:80-84.
38,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.494.59.435.62.277.2]Siegl, G.,
classification of virus particles associated withhepatitis nentsofthe virus particle. Virology57:531-542.