JOURNAL OFVIROLOGY, July1981,p. 82-86
0022-538X/81/070082-05$02.00/0 Vol.
39,No.1
Structural
Changes
in BHK
Cell Plasma
Membrane Caused
by the
Binding of Vesicular Stomatitis Virus
LARRY D. ALTSTIELtANDFRANK R.LANDSBERGER*
TheRockefellerUniversity, New York, New York 10021
Received6January1981/Accepted 6April1981
Spin label electron spinresonance techniquesusinganitroxide derivative of
stearic acidwereusedtodetectchangesinplasmamembranestructurecausedby
thebinding of vesicular stomatitis virus(VSV)tocellplasmamembranes of intact
BHK-21 cells. Theresults indicate thatbindingof VSVtocell surfacereceptors
causes anincrease in theobservedrigidityof theplasmamembranelipidbilayer.
Thischange in membrane structure, which appearstobe causedby the
cross-linking ofreceptorsin theplane of the plasmamembrane,could bepreventedby
treating the cellswithcolchicine before addition of virus and could be reversed
bytreating the cells with colchicine after addition of virus. Cells treated witha
monovalent, water-soluble derivative of VSV G-protein
(G.)
did not show anincreaseinplasmamembranebilayer rigidity. However,additionofanti-VSV
G-protein immunoglobulin G to cells pretreated with
G.
caused an increase inplasma membranebilayer rigidity.Thisincreasedrigiditycould also be reversed
by the addition of colchicine.Fluorescencemicroscopywasusedtodetermine the
distribution offluorescein-labeled VSVparticlesonthecell surface after addition
of virus.Approximately 30minafter addition ofvirus,discreteareas onthecell
surface showedfluorescentstaining,which coalescedtoapical regionsof thecell
afterapproximately40min.
Theinitialeventinthereplicative cycleofan
animal virus is theadsorption of the virus
par-ticle tothe host cellplasmamembrane (8, 30).
This attachment involves specific interactions
between the virion and the plasma membrane
surfaceproteins andlipids capableoffunctioning
asvirusreceptors(13,15,30).Virusbindingcan
inducechanges in the lateral distribution of cell
membrane receptors by causing the formation
of virus receptor patches in the plane of the
plasma membrane (3, 14, 15). Attachment of
influenza and Sendai virusesorlectins alters the
fluidity ofthe membrane lipidbilayer ofavian
erythrocytes (31, 32). This effect requires the
cross-linking ofreceptors andcan beinhibited
withmicrotubule-disruptingdrugs (31, 32).
Weinvestigatedplasma membrane structural
changescausedbythebindingoftheenveloped,
negative-strand virus vesicularstomatitis virus
(VSV)totissueculture cellsby using spinlabel
electron spin resonance (ESR) methods. VSV
bindstocellsvia anenvelope-associated surface
glycoprotein (G-protein) (4, 5, 6, 22). ESR
spec-traof anitroxide derivativeofstearic acid
incor-t Presenincor-t address: Deparincor-tmenincor-t of Cellular and
Develop-mentalBiology, The Biological Laboratories, Harvard Urni-versity,Cambridge, MA 02138.
porated into the plasma membrane of intact
BHK-21 cells indicate that the plasma
mem-brane lipid bilayer becomes more rigid upon
adsorption of VSV. This increase inrigiditycan
be inhibitedby colchicine.
Ligands thataremonovalentorhavereduced
valencyaregenerally unabletoinduce clustering
ofreceptors in the plane of the plasma
mem-brane (12, 18, 23, 36). Cells infected with VSV
produceawater-soluble glycoprotein that is
de-rived fromG-protein(20, 21, 28, 29). This soluble
glycoprotein,
G.,
appears tobemonovalent(un-published data). It isreported here that
treat-mentofcells withG8 doesnotcause a change in
plasma membrane lipid bilayer fluidity. When
immunoglobulin G(IgG)directedagainst
G-pro-tein is added to cells pretreated with G8, the
plasmamembrane lipid bilayer becomes more
rigid. By measuring the distributionof
fluores-cently labeled VSV on the cell surface, it was
found thatboundvirusformeddiscrete clusters
on the cell surface and that these patches of
bound virus moved toapical regions of thecell
approximately 40 min after addition of virus.
These results suggest that binding of VSV to
cellplasmamembranereceptors involved a col-chicine-sensitive lateralrearrangementof recep-torsin theplane of the plasmamembrane. 82
on November 10, 2019 by guest
http://jvi.asm.org/
VSV ATTACHMENT TO BHK CELLS
MATERLALS AND METHODS Cells. MonolayersofBHK-21cellsweregrownon
Falcon (FalconPlastics,Oxnard, Calif.) plastictissue cultureflasks inreinforced Eaglemedium containing 10% tryptose-phosphate broth and 10% calf serum
(16).
Virus. Stocksof theIndianaserotypeof VSVwere
maintained andassayed on BHK-21 cells (7). Virus was prepared by infecting BHK-21 cells with VSV
suspendedinEaglemediumatamultiplicity of
infec-tionequalto0.1 PFUpercell. Aftera2-hadsorption
period,theinoculumwasremovedandreplacedwith
Dulbecco medium. Thesupernatantfromthe infected cellswasremovedapproximately14hafter infection
at 37°C. Celldebris wasremoved bylow-speed
cen-trifugation (2,500 xg,20min).Viruswasrecovered
from theclarifiedsupernatantmediumbyhigh-speed centrifugation (25,000x g,2h).Thepelleted viruswas
purifiedbydensitygradient centrifugationonlinear5 to40%potassiumtartrategradients(26).
Preparationofsolubleglycoprotein.The
super-natant medium recovered after virus pelleting was
brought to 70% saturation with ammonium sulfate. Theprecipitated protein wasrecovered by
centrifu-gation,and theresulting pelletwassuspendedinwater anddialyzed against 10mM Tris0.150 M NaCl(pH
V\WN\CO
COOH
C5 N
10GAUSS
7.4) buffer (TBS). The dialyzed protein mixture was subjected to high-speed centrifugation (100,000xg, 6 h) to remove insoluble aggregates. Final purification wasachieved bySepharose 4B gel filtration chroma-tography and eluting with TBS.
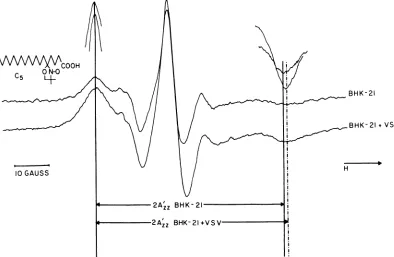
Spinlabeling. The cellswereharvested by gentle scraping. They were washed with phosphate-buffered saline, and10 1A of1-mg/ml C5 stearic acid spin label [2-[carboxypropyl]-4,4 dimethyl-2-tridecyl-3-oxazoli-dinyloxyl] (structure shown in Fig. 1) in ethanol was added to the cellpellet. The cells were immediately washed withphosphate-buffered saline and aspirated into acapillary pipette. ESR spectra were recorded with a Varian E-12 ESR spectrometer. All spectra wererecorded at room temperature.
Fluorescent labeling and fluorescence micros-copy.Fluorescein isothiocyanate atafinal concentra-tion of100FsMwasaddedtoa2-mg/ml suspension of VSV in 10 mM sodium phosphate buffer (pH 8.5). Aftera12-h incubation period at 4°C, the virus sus-pension was dialyzed against phosphate-buffered sa-line to remove the unbound label, and the labeled viruswasrepurified as described above. The labeled virus in excess was added tocellsgrown ongla cover slips. At 5-min intervals after addition of virus, the coverslipswere mountedon glass microscopeslides and washed withphosphate-buffered saline, and the
BHK-21
BHK-21+VSV
H
FIG. 1. ESR spectraoftheC5 spinlabelincorporatedinto theplasmamembranelipidbilayerofBHK-21 ceUs. Thestructureofthespinlabel is shown in the upperleft. The upperspectrumis thatofthespinlabel incorporatedintocontrol BHK-21 cellplasmamembranesofintact cells. The lowerspectrum is thatofthe spin label in theplasmamembraneofintact cells which had been treated with 100 PFUof VSVpercellat 37°Cfor30minbeforebeingharvested andspinlabeled. The distance between the outermostpeaks in the spectrumisdefinedas2A'=.Theoutermostpeaks ofthespectrawereamplifiedto aid in the determinationof
2A'z2.
83 VOL. 39, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.46.440.320.577.2]84 ALTSTIEL AND LANDSBERGER
cells wereexamined for bound fluorescence (9).
Incu-bations with virusweredoneat37°C,andmicroscopy wasdoneatroomtemperature.
Reagents.Fluoresceinisothiocyanatewasobtained from SigmaChemicalCo.,St. Louis,Mo. Spinlabel
waspurchasedfromSyva, PaloAlto,Calif. All other chemicals used were the highest grade obtainable. Anti-VSVG-proteinIgGwasisolatedbyion-exchange chromatographyonDEAE-cellulose (27)fromrabbit
antiserumagainstpurified VSVG-protein.Rabbit
an-tiserumwaskindlyprovidedbyJohnLenard, College of Medicine andDentistryof NewJersey,TheRutgers MedicalSchool.
RESULTS
Figure1shows the ESRspectraoftheC5 label incorporated into the plasma membrane of
un-treated BHK-21cellsand of BHK-21cells which
had been treated with 100 PFU of VSVpercell
inEagle medium withoutserumat37°C for 30 min before harvesting and spin labeling. As dis-cussedpreviously in detail, it appearsthatthe
ESR signal detected from C5-labeled intact BHK-21 cellsis due tospin labelincorporated into theplasma membranelipid bilayer andnot to spin label incorporated into cytoplasmic membranes (25). Thebinding of VSV caused a
slight, but easilydetectable, increaseinthe dis-tancebetween the outermostpeaks of the
spec-trum (2A'2. defined inFig. 1), indicating an in-creaseinthe averagerigidity of the membrane
lipidbilayer (17, 19, 24). The ESRspectra there-fore suggest that binding of VSV to BHK-21
cellsresults inastructuralchange in the plasma
membranelipidbilayer. Thisstructuralchange reachesamaximumapproximately 30 min after
addition ofvirus.
BHK-21cellsweretreated withmultiplicities
ofVSV, ranging from 10 to 100 PFU per cell,
andspin labeled after a 30-minadsorption
pe-riod. The increaseinplasma membrane bilayer rigiditycaused bytheaddition of virus is mul-tiplicitydependent,requiring50PFUpercellto
produceameasurable increase in therigidity of
theplasmamembrane, andsaturatesat
approx-imately 100 PFUpercell.
The VSV virion contains many copies of
G-protein. It is therefore possible that each VSV particle bindstoseveralG-proteinreceptorson
the cellsurface. Tomeasuretheeffects ofligand
valency on membrane fluidity, these
experi-mentswererepeated with the monovalent
deriv-ative ofG-protein, G8. Addition of1mgofG8per
ml inphosphate-buffered saline didnotcausea
measurablechange in bilayer rigidity. However, whenanti-G-protein IgG, which specifically
pre-cipitates G. (33), wasadded tocellspretreated
withG.,theESRspectraofthe C5 probe incor-porated into the membrane indicated that the plasma membrane bilayer became more rigid
J. VIROL.
(Table 1). These results and the fact that the
ratio ofvirallipid toplasmamembranelipid in theseexperimentsis verysmall indicate thatthe
observedchanges in the ESRspectrumof
spin-labeled plasma membranes are not due to
dif-fusion of spin label into the envelopes of the
virusparticles.
Preincubation of the cells with 10
,uM
colchi-cine in Eagle medium for 30 min at37°C
pre-vented bothvirus-inducedchanges in membrane
bilayer fluidity and alteration of membrane
ri-gidity due to the cross-linking of
membrane-bound G5 by IgG (1). The increase in plasma
membranebilayer rigidity could be reversed by
theaddition of colchicinetoVSV-treated cells.
Preincubation of the cells with the metabolic
inhibitorsdinitrophenol andF- (1mM
dinitro-phenol-10 mM NaFin phosphate-buffered
sa-line) didnotsignificantlyinhibit theincreasein
bilayer rigidity causedby VSV.
The results ofthe ESR experiments suggest
that binding of VSV and the cross-linking of
membrane-bound
G.
by anti-G-protein IgGcause a change in the fluidity of the BHK-21
plasma membrane lipid bilayer. The multisite
binding of a polyvalent ligand may cross-link
plasma membrane receptors, causing a change
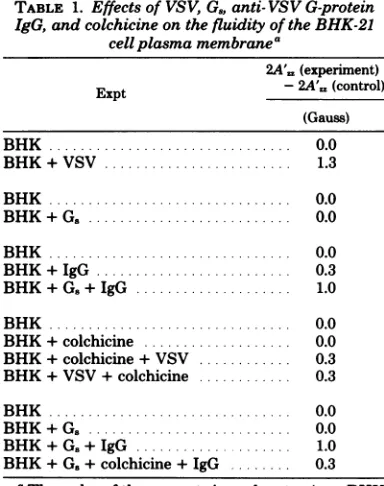
TABLE 1. Effectsof VSV,G.,anti- VSVG-protein IgG, and colchicineonthefluidity ofthe BHK-21
cellplasma membranea
2A'= (experiment)
Expt - 2A',, (control)
(Gauss) BHK... 0.0
BHK+VSV ... 1.3
BHK... 0.0
BHK + G .. ... 0.0
BHK ... 0.0
BHK+IgG ... 0.3
BHK+G.+IgG ... 1.0
BHK ... 0.0
BHK+colchicine ... 0.0 BHK+colchicine+VSV ... 0.3 BHK+VSV+colchicine ... 0.3
BHK ... 0.0
BHK+G. ... 0.0 BHK+G.+IgG ... 1.0
BHK+G.+colchicine+IgG ... 0.3
a The order of the reagents in each entry (e.g., BHK +G.+IgG) represents the order in which the various reagents were added to the cells before harvesting andspinlabeling. A changein 2A',a (experiment) -2A'=, (control) greater than 0.3 Gauss indicates that the treatment inthetableentry caused anincrease in therigidity ofthecellplasma membrane lipid bilayer.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.259.451.343.586.2]in thelateral distribution of proteinsin the plane of the plasma membrane.
VSV labeled with fluorescein isothiocyanate
wasadded to cells at370C,and the pattem of
fluorescent stainingonthe periphery ofthe
un-fixed cellswas examined at 5-min intervals after
additionof virus.Few areas oflocalized
fluores-cence wereobservedbefore20 minafter addition
offluorescein isothiocyanate-labeledVSV.
How-ever,after 20 min, thefluorescence was in the
form ofpatches around the periphery ofthecell.
Thecapping appearedtobecompleteby 40 min.
(Relatively little intracellular fluorescence was
observed40min afterinfection.) This resultand
observationsby others (3, 14)suggestthatvirus
bindingcaninduce the lateralrearrangementof
cell surface proteins.
DISCUSSION
The data presented indicate that binding of
VSV to BHK-21 cells causes ameasurable
in-crease in the rigidity of the plasmamembrane
lipidbilayerasdetected withanitroxide
deriv-ative of stearic acid. In addition, the datasuggest
that this increase in rigidity is sensitive to the
microtubule-disrupting drug colchicine.
Lyles and Landsberger (31, 32) have shown
thatthefluidity of avian and amphibian
eryth-rocyte membrane lipid bilayers increases upon
agglutination by influenza and Sendai virionsor
lectins and that this change in bilayer fluidity
requires multisite binding andcanbe inhibited
by colchicine (31, 32). The change in bilayer
fluidityobserveduponbindingofVSVto
BHK-21 cells is in the opposite direction from that
observed for nucleated erythrocyte systems.
This probably reflects the differences in lipid
and protein composition and in lipid-lipid and
lipid-protein interactions in the plasma
mem-branes and not differences in themechanisms
underlyingthe observed changesin the plasma
membranesupon adsorptionof virus.
Unfortu-nately, at present little is known about these
interactions; thus, it is not possible to give a
descriptionof the mechanismsunderlyingthese
results. The observedchangesinmembrane
bi-layer fluidityin bothsystemsdorequirebinding
ofmultivalentligands and aresensitive to
col-chicine, suggesting that a similar mechanism
maybe involved. Both thebindingofinfluenza
and Sendai viruses to avian erythrocytes and
the binding ofVSV toBHK-21 cells appear to
involve the lateral rearrangement of membrane receptors, asreflectedbytherequirementthat (i) the receptors be cross-linked and (ii) the apparentdependenceof thebinding-induced
bi-layerfluidity changeuponmicrotubule-like
sys-temswhichmay be involved inreceptor
mobility
in manymembranesystems (10, 31, 32,35).
The observation that VSV bound to the
sur-face of the BHK-21 cell membrane can form
patches and capsprovides additional evidence
for the hypothesis that virus binding involves
lateral redistribution ofcellsurface receptors. It
hasbeen postulated that membrane structural
changes suchasreceptor rearrangement are
in-volved in endocytosis(2, 36), the probable
mech-anism forVSV penetration (8, 11, 34). At
pres-ent, a distinction cannot be made between a
direct effect of receptor rearrangement on
bi-layer fluidity andsome event subsequent to
ad-sorption, such as endocytosis. However, since
relatively littleintracellular fluorescencewas
ob-served during the time periods when capping
and patching occurred, and since the
virus-in-ducedincrease in plasma membrane bilayer
ri-gidity occurred in the presence of a metabolic
inhibitor(NaF) whichinhibits endocytosis(37),
itappearsthat thealteration ofmembrane
fluid-ity observeduponaddition ofvirus is due to the
cross-linking of cell membranereceptors, rather
thanto a metabolically coupled event such as
endocytosis.
ACKNOWLEDGMENTIS
We thank EdwardGershey for assistance with the fluores-cencemicroscopy.
This work wassupported byPublicHealth Service grant AI-14040 from the National Institutes of Health and by Na-tionalScienceFoundationgrantsPCM78-09346 and PCM 79-22956.L.D.A. was a Public HealthService trainee. F.R.L. is anAndrew W.MellonFoundation Fellow.
LITERATURE CITED
1. Altstiel,LD.,and F. R.Landaberger.1977. Interac-tions of colchicine with phosphotidylcholine mem-branes. Nature(London) 269:70-72.
2. Berlin,R.D.,and J. P. Fera 1977.Changesin mem-branemicroviscosityassociatedwithphagocytosis ef-fects ofcolchicine. Proc. Natl. Acad. Sci. U.S.A. 74: 1072-1076.
3. Birdwell,C.R., and J. H. Strauss. 1974. Distribution of the receptor sites forSindbis virusonthesurface of chicken and BHK cells. J. Virol. 14:672-678. 4. Bishop,D.H.L.,P.Repik,J. F.Obijeski,N. F.Moore,
and R. R.Wagner. 1975.Restitution ofinfectivityto spikelessvesicular stomatitis virusby solubilized viral components. J. Virol.16:75-84.
5. Cartwright, B., G J. Smale, and F. Brown. 1969.
Surfacestructureof vesicular stomatitis virus. J. Gen. Virol.5:1-10.
6. Cartwright,B., P.Talbot,and F. Brown. 1970.The proteins ofbiologically active sub-units of vesicular stomatitisvirus. J.Gen. Virol.7:267-272.
7. Choppin,P.W.,and R. W.Compans.1970.Phenotypic mixing ofenvelope proteinsof theparainfluenzavirus
SV5 and vesicular stomatitisvirus. J.Virol.5:609-616. 8. Dales,S.1973.Earlyeventsincell-animalvirus
interac-tions.Bacteriol.Rev. 37:103-135.
9.D'Alisa,R.M.,D. R.Korf,and E.L Gershey.1979.T
antigenbanding on chromosomes of simian virus 40
infectedmuntjac cells.Cytogenet.Cell Genet.24:27-36. 10. Edelman,G.M. 1976.Surface modulationincell
recog-nition andcellgrowth.Science 192:218-226. 11. Fan,D.P.,andB. M.Sefton.1978.Theentryintohost
cells of sindbis virus, vesicular stomatitis virus and
on November 10, 2019 by guest
http://jvi.asm.org/
86 ALTSTIEL AND LANDSBERGER Sendai virus. Cell 15:985-992.
12. Gunther, G. R., J. L. Wang, L. Yahara,B. A.
Cun-ningham, and G.M.Edelman.1973.ConcanavalinA
derivativeswith alteredbiological activities.Proc. Natl.
Acad. Sci. U.S.A. 70:1012-1016.
13. Haywood, A. M. 1974. CharacteristicsofSendaivirus
receptorsinamodelmembrane. J. Mol.Biol.
83:427-436.
14. Helenius, A.,J.Kartenbeck,K.Simons,and E.Fries. 1980.On theentryof Semliki Forest virus intoBHK-21 cells.J. Cell Biol. 84:404-420.
15. Helenius, A.,B.Morein,E.Fries,K.Simons,P.
Ro-binson, V. Schirrmacher, C. Terhorst, and J. Strominger.1978.Human(HLA-AandHLA-B)and
murine (H-2K andH-2D)histocompatibility antigens
are cellsurfacereceptorsfor Semliki Forestvirus. Proc. Natl. Acad. Sci. U.S.A.75:3846-3850.
16. Holmes,K.V.,and P. W.Choppin.1966. On the role of
theresponseof thecellmembrane indetermining virus
virulence.Contrasting effects of the parainfluenza virus SV5in twocelltypes.J.Exp.Med.124:501-519.
17. Hubbell,W.L.,and H. M.McConnell. 1971.Molecular
motion inspin-labeledphospholipids and membranes. J.Am.Chem. Soc. 93:314-326.
18.Joseph,B.S.,and M. B. A.Oldstone. 1974. Antibody-inducedredistributionofmeasles virusantigensonthe
cell surface. J. Immunol.113:1205-1209.
19. Jost, P., L. J. Libertini, V. C. Hebert, and 0. H.
Griffith.1971.Lipid spinlabels inlecithinmultilayers.
Astudy of motionalong fattyacidchains.J. Mol. Biol.
59:77-98.
20. Kang,C.Y.,and L.Prevec. 1969. Proteins ofvesicular stomatitis virus.I.Polyacrylamide gel analysis of viral antigens.J. Virol. 3:404-413.
21. Kang,C.Y.,and L. Prevec. 1971.Proteins of vesicular
stomatitis virus.III.Intracellularsynthesis and
extra-cellularappearanceofvirus-specific proteins. Virology
46:678-690.
22. Kelley,J.M.,S.U.Emerson,and R.R. Wagner.1972.
The glycoprotein of vesicular stomatitis virus is the antigen that gives risetoandreactswithneutralizing antibody. J.Virol.10:1231-1235.
23. Lampert,P.W., B. S. Joseph, and M. B. A. Oldstone.
1975.Antibody-induced capping of measles virus
anti-gens onplasmamembranestudiedby electron
micros-copy.J. Virol.15:1248-1255.
24. Landsberger, F.R., and L.D. Altstiel. 1980. Lipid-protein interactions in envelopedviruses. Ann. N.Y.
J. VIROL.
Acad. Sci.348:419-423.
25. Landsberger, F. R., and R. W.Compans. 1976. Effect ofmembraneproteinonlipid bilayer structure: a spin-label electronspin resonance study of vesicular
stoma-titis virus.Biochemistry 15:2356-2360.
26. Landsberger, F. R., J. Lenard, J.Paxton, and R. W. Compans. 1971. Spin-label electron spin resonance study of the lipid-containing membrane of influenza virus.Proc. Natl.Acad. Sci. U.S.A. 68:2579-2583. 27.Levy, H. B., and H. A. Sober. 1960. A simple
chromato-graphic method for thepreparation of gamma globulin. Proc. Soc.Exp. Biol. Med. 103:250-252.
28. Little, S. P., and A. S. Huang. 1977. Synthesis and distribution of vesicular stomatitis virus-specific poly-peptides in the absence of progenyproduction. Virology 81:37-47.
29. Little, S. P., and A. S. Huang. 1978. Shedding of the glycoprotein from vesicular stomatitis virus-infected cells. J. Virol. 27:330-339.
30. Lonberg-Holm, K.,andL.Phillipson. 1974.Early in-teractions between animal viruses and cells. Monogr. Virol. 9:1-148.
31. Lyles, D. S., and F. R.Landsberger. 1976. Virus and lectinagglutination of erythrocytes: spin label study of membranelipid-protein interactions. Proc. Natl. Acad. Sci. U.S.A. 73:3497-3501.
32. Lyles, D. S., and F. R.Landsberger.1978.Enveloped virus- and lectin-inducedstructural change in erythro-cytelipidbilayer: dependence on species of erythrocyte andonthemultivalence of theagglutinin. Virology 88: 25-32.
33. Miller, D. K., B. I. Feuer, R. Vanderoef, and J. Len-ard. 1980. Reconstituted Gprotein-lipid vesicles from vesicular stomatitis virus and their inhibition of VSV infection. J.Cell Biol. 84:421-429.
34.Miller,D.K., and J. Lenard. 1980. Inhibition of vesicular stomatitis virus. Evidence for anintracellular, G-pro-tein-requiring step. J. Cell Biol. 84:430-437.
35. Nicholson, G. L. 1976.Transmembrane control of the receptors on normal and tumorcells. I. Cytoplasmic influence over cell surface components. Biochim. Bio-phys. Acta 457:57-108.
36.Scheckman,R., and S. J. Singer. 1976. Clustering and endocytosis of membrane receptors can be induced in matureerythrocytes of neonatal but not adult humans. Proc. Natl. Acad.Sci. U.S.A.73:4075-4079.
37. Silverstein,S.C., R. M.Steinman,andZ. A. Cohn.
1977.Endocytosis. Annu. Rev. Biochem. 46:669-722.
on November 10, 2019 by guest
http://jvi.asm.org/