0022-538X/83/010408-12$02.00/0
Copyright©1983,AmericanSocietyfor Microbiology
Expression of Adenovirus Type 12 ElA Gene in Monkey
Cells, Using
a
Simian Virus 40
Vector
KINICHIRO ODA,* HIROYUKI KATO, IZUMU SAITO,t SUMIO SUGANO, KAZUO MARUYAMA,
MIYUKIMASUDA, KAZUKO SHIROKI, ANDHIROTO SHIMOJO
DepartmentofTumor VirusResearch, InstituteofMedical Science, UniversityofTokyo,4-6-1, Shirokanedai,
Minato-ku, Tokyo 108,Japan
Received 24 June 1982/Accepted 27 September 1982
Simian virus 40(SV40) recombinants carrying the adenovirustype12ElAgene were constructed. The SV40 expression vector was constructed by removing mostof the VP1geneandaninternalpartof the interveningsequencefor late 16S
RNAand by joining the 5' and 3' splice sites intoasmallsegment.The adenovirus
type 12 ElA gene with or without its own promoter was inserted downstream
from the SV40 latepromoterand the splicing junctions. The recominant DNAwas
propagated and packaged in monkey cells by cotransfection with an early
temperature-sensitive mutant(tsA58)DNAashelper. Immunofluorescent staining
of themonkey cells infected with the resulting virus stocks showed thatupto20% of the cellsoverproduced the ElAgeneproducts in the nuclei. Two-dimensional
gel electrophoresis of the products indicated that the products werevery similar or identical to the authentic polypeptides synthesized in adenovirus type 12-infected human embryo kidney cells. The ElA mRNAwasinitiated atthe SV40 latepromoterirrespective of thepresenceof the EIApromoterand terminatedat
either the ElA orthe SV40polyadenylation signal. These hybrid mRNAs were
correctly spliced in the ElAcoding region.
The early adenovirus genes ElA and E1B, located in the leftmost11% of the viralgenome, are involved in both the transformation of sus-ceptible cells in culture andthe lytic growth of virus in permissive cells (34). The ElA gene alonecantransformthecells but isnotsufficient to completely transform them (11, 30). During lytic infection, the ElA gene is expressed first (21) and promotes the accumulation of tran-scripts from theotherearlygenes,E1B, E2, E3, and E4 (pre-early function), since the amounts ofthesetranscripts aregreatly reduced in cells infected with host range and deletion mutants defective inthe ElAgene (1, 12). Two alterna-tive mechanisms have been proposed for this accumulation ofearly transcripts, one acts at a transcriptional level (20) and the other at a posttranscriptional level (25).
The ElA gene encodes several related pro-teinsby synthesizing multiple speciesof mRNA which receive differentpatterns of RNAsplicing (8, 27, 29). In the case of adenovirus type 12
(Adl2),
a clusterof severalproteinswith molec-ular weights of 35,000 to 40,000 have been identified (29). These proteins werelocalized in thenuclei of infectedandtransformed cells and were designated as T antigen g based on theirtPresentaddress: Department of Microbiology,Facultyof
Medicine, UniversityofTokyo, Tokyo113, Japan.
granular appearance revealed by immunofluo-rescence. In spite of the interesting multiple functions ofthe ElAgene, no definitebiological function of the ElA gene products has been characterized in vitro, mainly due to a low amountoftheproductsproducedin adenovirus-infected cells.
Simian virus 40 (SV40) has been used as a
vectorforcloningandforexpression ofavariety ofeucaryotic genes in monkey cells (3, 5-7, 9, 15, 18, 24, 32), owing to its simple, well-ana-lyzed genomic structure. In the present study, SV40recombinantscarryingtheAd12 ElA gene wereconstructedtohyperproducethe ElA gene products.The SV40expression vector was con-structed by removing almost all of the coding sequence of the major capsid protein VP1 and most ofthe intervening sequence for late 16S RNAandby joiningthe 5' and 3'splicesitesinto a small segment. By usingthis vector, DNA of up to about 1,900 basepairs (bp)canbeinserted, irrespective of the presence of its own splicing junctions. The Adl2 ElA gene inserted in this vector was efficiently expressed in monkey cells.
MATERIALS ANDMETHODS
Cells and viruses. Established cell lines of African green monkey kidney, CV1 and GC7 (36), and KB cellswerecultivatedinEaglemedium with10%calfor
408
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 RECOMBINANTS CARRYING Adl2 ElA GENE 409
fetal calfserum. Adominant-selection vector,
pSV2-gpt(16, 17) was kindly provided by Paul Berg,
Stan-ford University, StanStan-ford, Calif. 3YgElAis a cell line
established after transfectionof a clonal line of Fischer
rat embryo fibroblast 3Y1-K with pSV2-gpt-EIA
DNA.pSV2-gpt-ElA wasconstructed by ligation of
the leftmost 4.5% ofthe Adl2 DNA with the large
EcoRI-BamHI fragment from pSV2-gpt by using
EcoRI and BamHI linkers. Adl2 (Huie strain) and
SV40 early temperature-sensitive mutanttsA58 were
propagated and titrated in human embryo kidney
(HEK)and CV1 cells, respectively.MutanttsA58 was
kindly providedby Peter Tegtmeyer, State University
ofNewYork at Stony Brook.
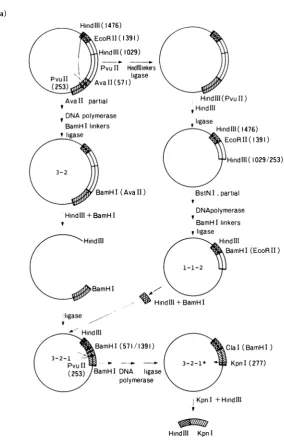
Constructionof recombinant viruses.Theprocedure
for the construction of the recombinant viruses is
diagrammed in Fig. 1.
(i) Construction of anSV40 DNA segment containing thesplice sitesfor late 16S RNA.Cloning ofthe 383-bp DNA segment containing both 5'and 3' splice sites for
SV40 late 16S RNA in pBR322 (Fig. la) was carried
outin PaulBerg's laboratoryatStanford University in
collaboration with Hiroto Okayama.
The pBR322-SV40 recombinantDNAcontaining the
SV40 latesequences from thePvuII site (253)to the
Hindlil site(1,476)(BBB numberingsystem)(34)was
partially digested with Avall, and full-length linear
DNA waspurified by extraction by the glass powder
technique (35), modified byPaterson,afteragarosegel
electrophoresis. The DNA was treated with the
Klenow fragment of Escherichia coli DNA
polymer-ase) I to convertthe cohesive termini toblunt ends,
ligated to synthetic BamHI linkers with T4 DNA
ligase, and cleaved with BamHI. TheDNAwasused
to transform E. coli HB101 afterligation. An Ampr
transformantcontaining theplasmid(no. 3-2inFig.la)
with a BamHI linker at the Avall site (571) was
selected.
The pBR322-SV40 recombinant DNA (positions
253-1,476)wascleaved withPvuII,ligatedtoHindlIl
linkers, cleaved withHindIll,and introducedintoE.
coli HB101 after ligation. The resultingplasmidDNA
was cleaved with HindIII and electrophoresed in ar,
agarosegel. The largestDNAfragmentofpBR322was
ligated to a small SV40 DNA fragment (positions
1,029-1,476) after extraction and introduced into E.
coli HB101. The plasmid DNA was then partially
digested with BstNI (which recognizes the same
se-quence as EcoRII), and full-length linear DNA was
extracted afteragarosegel electrophoresis. TheDNA
wastreated withE.coliDNApolymerase I, ligatedto
BamHIlinkers, andcleaved withBamHI. The DNA
wasintroducedintoE.coliHB101afterligation,and a
transformantcontaining theplasmid (no. 1-1-2in Fig.
la) with aBamHIlinkeratthe EcoRIIsite(1,391) was selected.
Plasmid 3-2 DNA was cleaved with HindIll and
BamHI,and thelargefragmentcontainingthepBR322
sequences wasisolated. This fragmentwasligatedto
the85-bp fragment isolated from plasmid 1-1-2 DNA
aftercleavage withHindlll and BamHI. A
transform-antcontainingplasmidno.3-2-1(Fig.la)wasselected.
The plasmidDNAwascleavedwithBamHI,treated
withE.coliDNApolymeraseI,andligatedtodestroy
theBamHI site. Atransformant containingthis
plas-mid (no. 3-2-1* in Fig. la) was selected, and the
generation ofaClaIsiteattheoriginalBamHI sitewas
confirmed. The 383-bp fragment containing both 5'
and 3' splice sitesforSV40late 16S RNA splicing was
isolated after cleavage oftheplasmidDNAwithKpnI
andHindIll.
(ii)Cloning of the Adl2 ElA gene.The Adl2SalIC
fragment (positions 0-3,497) was cloned in pBR322.
Thisrecombinant plasmid(PASC in Fig. lb)DNA was
cleaved with either HaeII and AccI orTaqI and AccI
(Fig. lb). The former cleavage generates a 1,100-bp
Adl2DNAfragment covering thecodingregion of the
ElA gene and its transcription termination signal,
whereas thelattercleavagegenerates a1,257-bp
frag-mentwhich alsocoversthepresumptiveElA
promot-ersignal. TheseDNAfragmentswereligatedtoa1:1
mixtureofHindIll and BamHI linkers after the
forma-tionof blunt ends byE. coliDNApolymerase I and
were cleaved with HindIll and BamHI. The digests
wereelectrophoresedon a 1.2% agarosegel, andthe
Adl2 DNA fragments were extracted. These DNAs
were ligated to the largeHindIII-BamHl fragment of
pBR322and introduced intoE.coliHB101. The
trans-formants containing the recombinant plasmid with
HindlIl and BamHI linkersatthe5'and3' sides of the
ElAgene wereselected.
(iii) Construction ofSV40-Adl2 EIA recombinants.
ThepBR322-SV40recombinant (no. VIII in Fig. lc) in
which whole SV40 DNA wasinserted at theBamHI
site ofpBR322 DNA was cleaved completely with
KpnIandpartially with BamHI. Afterelectrophoresis,
the larger fragment containing the whole pBR322
sequences and the SV40 early region, including the
origin, was isolated (Fig. lc). The DNA was mixed
with an equimolar ratio ofthe 383-bp fragment
con-taining the SV40late16SRNAsplicingjunctions(Fig.
la) and the Adl2 DNAfragment covering the ElA
coding sequences (Fig. lb) and was ligated with T4
DNA ligase. The DNA was introduced into E. coli
HB101, and thetransformantscontaining the plasmid
with theexpectedsequencearrangement were
select-ed(Fig. ld).Amoiety of theSV40-Adl2 recombinant
couldeasily beseparated fromthepBR322sequences
bycleavage of thepBR322-SV40-Adl2double
recom-binant (pSAdElA or pSAdElAp in Fig. ld) with
BamHI and AccI, since AccI does not cleave the
recombinant sequence but cleaves the pBR322
se-quences at two sites.
Transfection. DNAtransfectionwascarriedoutwith
DEAE-dextranasafacilitatoressentially accordingto
themethod ofSompayrac andDanna(31). Confluent
culturesofCV1 orGC7 cells (ca. 1.5 x 107 and 5 x
106, respectively) were washed twice with Dulbecco
modified Eagle medium and incubatedwith 4 to 8 ,ugof
linear recombinant DNA plus0.2 ,ug of SV40tsA58
helper DNA in Dulbecco modified Eagle medium containing 0.05 MTris-hydrochloride(pH 7.3) and 200 ,ugofDEAE-dextran per ml at37°Cin aCO2incubator
for8h.Themonolayerswerethen washedtwicewith
Dulbeccomodified Eagle medium and fed with 50 ml ofthe same medium containing5% fetal calfserum.
The cellswereincubatedateither 41°C(CV1cells)or
39.5°C(GC7 cells) for12to16days untilthe
cytopath-ic effect was well developed and then were
freeze-thawedfivetimes. Thelysatewascentrifugedat3,000 rpm for 10 min, and the supernatant was used for
infection in allsubsequentexperiments.
Spot hybridization. Intracellular viral DNAs were
extracted from theinfected cellsby the procedure of
VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/
410 ODA ET AL.
(a)
Hind111( 1476)
EcoRII(1391) HindIII(1029)
Pvu II Hindlillinkers
Pv i ligase
(253) aII (571 )
iAvaII partial IDNA polymerase
BamHIlinkers Iligase
3-2
BamHI(Avail1)
Hind 111+BamHI Hind III
BamHI
ligase
ligase
HindI1l(1476)
EcoRII( 139 )
Hind1II(1029/253)
BstNI,partial
DNApolymerase BamHI linkers
Iligase
HindllI
BamH I(EcoR II)
01-1-2
t HindIIl+BamHI
KpnI +Hind11l
HindIll KpnI
FIG. 1. Construction of SV40 recombinants carrying the Adl2ElAgene.(a)Constructionof therecombinant
plasmid containingthe 5'and 3' splice sites for SV40late16SRNA in a small DNA segment. (b)Cloning ofthe
Adl2 ElA gene. (c) Preparation of the linear pBR322 sequences containing the SV40 early region. (d)
Construction ofpBR322-SV40-Adlfdouble-recombinantDNA.TheDNAfragments preparedinpanels a, b, and
cweremixed at anequimolarratio and ligatedasdescribedin the text.Solid lines, pBR322 sequences; openbars, SV40 orAdl2sequences;0, origin ofSV40 DNAreplication;shadedregions, SV40sequencescontaining the 5'
(striped regions) and3'(crosshatched regions) splice sites forlate16S RNA (caretsymbolsrepresent the sitesof
splicing); stippled regions, Adl2 sequences containing the ElA gene; P, Adl2 ElA promoter; T, Adl2
polyadenylation signal; A,ATG; A, TAA.
Hirt (10). Aliquots (0.5 ,ug) of heat-denatured DNA were spottedonto acirculararea markedon a
Milli-pore membrane filteras previously described (14) to determine the ratio ofrecombinant to helper DNA.
Reconstructionspotsweremade by mixingthe
recom-binant DNA with salmon sperm DNA. The immobi-lized DNAs were hybridized with 32P-labeled Adl2
DNApreparedbynicktranslation(26).Theamountof
recombinant DNAwas estimatedby the intensityof
thespots in theautoradiogram.
Si
nucleasemappingofhybridmRNAs. ThehybridmRNAcontainingthe Adl2 ElAsequence was
ana-lyzed by S1 nucleasemapping (2, 28).Thecytoplasmic
RNAsprepared from the infected cellswere
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.489.109.394.68.509.2]VOL. 45, 1983
(b)
SV40 RECOMBINANTS CARRYING Adl2 ElA GENE 411
Taq 1 (338) Hae II +AccI
3amH I (Acc I )
HindIllIaamHt I HindllBaBamH I
(c) BamH 1(2516)
(d)
BamHI
pSAdEl Ap
orn
P KpnI nd IIll
ligase
BamH I
II
ligase
FIG. 1. Continued
ized with uniformly 32P-labeled Adl2 EcoRI DNA
fragment C at 52°C for 3 h. Aftertreatment with SI
nuclease, the hybrids wereprecipitated with ethanol
and subjected to either alkaline or neutral 1.4%
ag-arose gel electrophoresis. The gels were dried and
autoradiographedwith Fuji X-ray film.
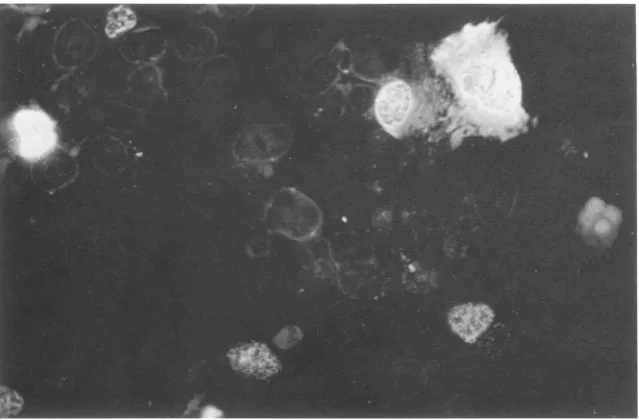
Immunofluorescent staining ofAdl2 T antigen g.
GC7 andCV1 cellson coverslipswereinfected with
the recombinantvirusstockandfixedwith acetone at
34 and 58 h after infection, respectively. Indirect
immunofluorescence wascarried outby using
anti-T-antigen-g serum from rats bearing HY cell tumors
followed by rabbit anti-rat immunoglobulin G
conju-gated withfluoresceinisothiocyanate (29).
Two-dimensional gel electrophoresis. The infected
cells werepulse-labeled with 200,uCi of
[35S]methio-nineperml for2 h in methionine-free mediumatthe
times indicated in the legend to Fig. 6. The cell
extracts were prepared by lysing the cells in lysis buffer containing 0.5% Nonidet P-40, 50 mM
Tris-hydrochloride (pH 8.0), 120mMNaCl, and150pLgof
phenylmethylsulfonyl fluoride per ml.
Two-dimen-sional electrophoresiswasperformed accordingtothe
methodof O'Farrell (23)with thefollowing
modifica-tions.Intheisoelectrofocusing gel,2%ampholine(pH
3.5 to 10) was used withoutmixing with the ampho-lines in the other pH ranges. Isoelectricfocusingwas for 12 h at 350 V. Thegelswereequilibratedand then
on November 10, 2019 by guest
http://jvi.asm.org/
applied to 15% polyacrylamide slab gels. The gels were dried andfluorographedwith Fuji X-rayfilms for 10days.
RESULTS
Construction of SV40 recombinants carrying theAdl2 EIA gene. SV40 recombinants carrying theAdl2 ElA gene were constructed and cloned inE. coli aspBR322-SV4O-Adl2 double-recom-binant plasmids (Fig. 1). From the published sequence data (33), the coding region of the Adl2 ElA gene resides between positions 502 and 1,374from the left end of the genome. The main RNA initiation site is located about 30 bp downstream fromthe TATA box at position 414. TwotypesofAdl2ElADNA wereinserted into theSV40vector;theHaeII-AccIDNAfragment (positions 495-1,595) contains a whole coding region together with the polyadenylation signal at position 1,436, and the TaqI-AccI fragment (positions 388-1,595) contains the main RNA initiation site inaddition to the sequence men-tioned above (Fig. lb). The latter fragmentwas used todeterminetheefficiencyof theutilization of the Adl2 ElA promoter in the SV40 vector. These fragments were cloned in pBR322 by using HindIll and BamHI linkers (Fig. lb).
To construct an SV40 vector which still re-tainsthelate mRNAsplice junctionsinaddition tothe initiation andtermination signals forlate mRNA, a small DNA segment in which the 5' and 3' splice sites forlate 16S RNAwerejoined by removing most ofthe intervening sequence was cloned in pBR322 (Fig. la). The DNA fragment containing the splice junctions was generated by cleaving the plasmid DNA with KpnI and HindIll.
For the construction of the pBR322-SV40-Adl2 double-recombinant plasmid,the pBR322-SV40 recombinant (no. VIII in Fig. lc) was
cleavedcompletelywithKpnIandpartiallywith BamHI; thelargeDNAfragment containingthe whole pBR322 sequences and the entire SV40 early region with thereplication origin (Fig. lc) was then mixed with an equimolar ratio of the Adl2 ElA DNA generated by cleavage with HindlIl and BamHI and the small fragment containing the splice junctions and was intro-duced intoE. coliafterligation. The
transform-antscontainingthedouble-recombinant plasmid wereselected aftercleavageof theplasmid DNA withanappropriate restriction enzyme (Fig. ld). These plasmids were designated as pSAdElA andpSAdElAp; theformercontained the Adl2 E1Acodingregionandpolyadenylation site,and the latter contained the main promoter for the ElA gene in addition to the former sequences. The precise structures of pSAdElA and pSA-dElApwereconfirmedbydigestionwith
AvaIl,
which cleaves these DNAsatmultiple sites(Fig. 2). An electrophoretic analysis ofthe products showed all of the DNAfragments larger than 500
bp
as having the expected lengths, suggesting that the recombinants were made exactlyac-cordingtotheschedule. Amoiety ofSV40-Adl2 recombinant DNA, SV-AdElA (4,470 bp) or
SV-AdElAp (4,672 bp), was obtainedby diges-tion of the recombinant plasmids with BamHI and AccI followed by gel electrophoresis. The latter enzyme, which cleaves the pBR322
se-quences at two sites but does not cleave the recombinant sequence,facilitated theseparation ofrecombinant DNA from pBR322 DNA (Fig. ld).
Propagation of SV40 recombinants carrying the Adl2 ElA gene. To increase the efficiency of infection, the recombinant DNA was packaged intovirionsby transfection of GC7 or CV1 cells with a mixture of the recombinant DNA and SV40 tsA58 DNA as helper at molar ratios of 20to40. The cells wereincubated at the nonper-missive temperature, and the virus stocks were prepared when the cytopathic effect was well developed. These stocks were used for the fol-lowing experiments. The amounts of recombi-nant and helper DNA in monkey cells infected with the virus stocks were first analyzed by ethidium bromide staining of the intracellular viral DNA after electrophoresis in an agarose
gel. However, owing to a low amount of the recombinant DNA, an exact estimation was
difficulttoperform. A quantitation of the
recom-binant DNA was, therefore, performed by spot hybridization with 32P-labeled Adl2 DNA as a
probe. Intracellular viral DNA(0.5pLg)washeat denatured and spotted onto acirculararea ona nitrocellulose filter. After hybridization, the amount of recombinant DNA wasestimated by comparing the intensity of the spots in the autoradiogram with that of reconstruction spots (Fig. 3). The results showed that the virus stocks prepared with SV-AdE1A and SV-AdE1Ap DNA in GC7 cells contained approximately 15 and 4% of recombinant genomes, respectively. This percentage was variable from preparation to preparation; however, the same difference between the amountofrecombinant genomes in SV-AdElA and that inSV-AdElApwas repeat-edly observed. The DNA sequence present only inSV-AdElAp(positions338-495)mayhave an inhibitoryeffect onthereplicationor encapsida-tion of the recombinant genome, or on both. Preparation of the virus stocks in CV1 cells alwaysresulted inareduction in the percentage ofrecombinant genomes.
Si nuclease mapping of Adl2 ElA mRNA
transcribed fromrecombinant viruses.The
struc-ture ofAdl2 ElA mRNA transcribed from the
recombinant DNA wasdeterminedbySI
nucle-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 RECOMBINANTS CARRYING Adl2 ElA GENE 413
A
Bam-lHl (2516)BarHI 1 45
(Acc
I1595)
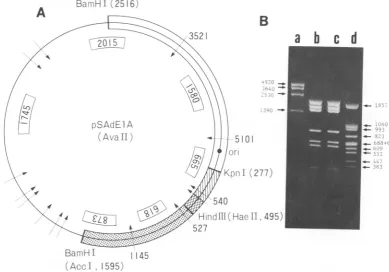
FIG. 2. Restrictionanalysis ofpBR322-SV40-Adl2double-recombinant DNA. (A) TheAvaIIrestrictionsites
inpSAdElADNA are shown by arrows. The numbers in boxes are the expected lengths of restriction fragments.
Thesamefragmentsof indicatedlengths must be generated from pSAdElAp DNA. Symbols are as described in
thelegend to Fig. 1. (B) pSAdElA and pSAdElAp DNAs digested withAvaIl andelectrophoresed on a1.4% agarosegel. Lanes: a,HindIll-digestedplasmidgABA DNA; d, BstNI-digested plasmid VIII (see Fig. 1)DNA;
b, pSAdElADNA; c,pSAdElApDNA.
ase
mapping
ofthe cytoplasmic RNAprepared
from the infected cells at the late phase of infection. The RNA was hybridized with uni-formly32P-labeled
Adl2EcoRI DNAfragment
C (0-16.5 mapunits).
AfterS1
nucleasetreat-ment, the DNA-RNA hybrid was electropho-resed in both alkaline and neutral agarose gels (Fig. 4). When the hybrid formed with SV-AdElAp RNA(the cytoplasmic RNAprepared from the SV-AdElAp-infected cells) was elec-trophoresed at alkaline pH, four segments of 730, 640, 450, and 310 basesweredetected
(Fig.
4, laneb).Theformertwosegmentscorrespond
to the
5'-coding
segments of the Al and A2mRNA species, respectively. The 5' termini of these segments correspond to the end of the insertedAdl2 DNA in therecombinant, indicat-ingthattranscription is initiated attheSV40late promoter rather than at the ElA promoter, which is located 110 nucleotides downstream from the end of the inserted Adl2 DNA. The faint band of 310 bases corresponds to the 3'-codingsegmentof theElAmRNAspecies from the 3' splice site to the ElA
polyadenylation
site,and the 450-base segmentof much strongerintensity corresponds to the region from the3' splice site totheotherendofthe inserted Adl2 DNA.The resultsindicate that mostofthe ElA transcript extends pastits own polyadenylation site and is polyadenylated at the downstream SV40 polyadenylation site. Judging from the intensity of the 730- and 640-base segments, almost equalamountsofAl and A2 mRNAwere
formed after processing of the primary
tran-script. An analysis of SV-AdElA RNA gave essentially the same results (Fig. 4, lane a). S1 nuclease treatment ofthe hybrid yielded three bands of570, 480 plus 450, and 310 bases. The former two bands correspond to the 5'-coding segment of the Al and A2 mRNA species,
re-spectively. The intensity of these bands was
strongerthan that ofbands generated from the SV-AdElAp hybrid, indicatingthat moreofthe hybridmRNAwaspresentin SV-AdElA-infect-ed cells thanin SV-AdElAp-infectedcells. The
amount was, therefore, proportional to the
amount of the recombinant virus in the virus
stocks.
S1 nuclease mapping ofthe DNA-RNA hy-brids in neutral agarosegels confirmed the
esti-VOL.45, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.489.59.449.77.349.2]2(ngi
100) 5 \/
0 41 A3
50 10
\ 20 / 44 /
O*
FIG. 3. Spot hybridization of intracellular viral
DNAfrom recombinant-virus-infected cells with32p_ labeledAdl2DNA.The virusstocks(Al, A2, A3, and A4) were prepared independently by transfection of
GC7cells withSV-AdElAandSV40 tsA58DNAs (Al
andA2)orwithSV-AdElApand tsA58 DNAs (A3 and
A4) at aweight ratio of40.Intracellularviral DNAs
were prepared from CVl cells infected with these
virus stocks. The intracellular viral DNAs (0.5
[Lg)
were spotted onto the filteras indicated and
hybrid-ized witha32P-labeled Adl2 KpnIfragment (1.7-16.0
mapunits). Reconstructionspots weremade with the
indicated amounts of SV-AdElA DNA plus salmon
spermDNA(total, 0.5 pLg).
mation described above. The hybrids formed with SV-AdElA RNA (Fig. 4, lane c) were
separated into four major bands of 1,020 (570 +
450), 930 (480 + 450),880 (570 + 310), and 790 (480+310) bp. The largertwobandscorrespond
to the Al and A2 species with their 3' termini extendedtothe end of the inserted Adl2DNA, and thesmallertwocorrespondtothe Al and A2 species polyadenylatedattheirown
polyadenyl-ation site. Similarly, the hybrids formed with SV-AdElAp RNA (Fig. 4, lane d)were
separat-ed into four major bands of1,180 (730 + 450), 1,090 (640 + 450), 1,040 (730 + 310), and 950 (640 + 310) bp. The lengths of these hybrids confirmed the initiation of transcription at the SV40 late promoter. The bands of 570 and 730 bp (Fig. 4, lanes candd) seem tobe generated by cleavage of the DNA-RNA hybrid at the splice site. The origin of the other faint bands hasnotbeendetermined.
Synthesisof Adl2 ElAgeneproductsin
recom-binant-virus-infectedcells.Theexpression of the ElA gene in the recombinant-virus-infected
monkey cellswasanalyzed by indirect
immuno-fluorescence. CV1 cells on cover slips were
infected withSV-AdElAorSV-AdElAp
recom-binant virus stocks at 41°C, and the cells were
stained with immunofluorescent antibody at 48 to60h after infection. The products (T antigen g) were detectedinthe nuclei(Fig. 5) and hada
morphology similar to that ofproducts synthe-sized in Ad12-infected HEK cells (29). The percentage of positive cells in SV-AdElA-in-fected cells varied between 10 and 20%, depend-ing on the virus stocks used, but did not exceed 20%, presumably due to a small amount of the recombinant virus in the virus stocks. The strongintensity ofimmunofluorescence, howev-er,indicates that T antigen g was overproduced in most of these positive cells. In SV-AdElAp-infected cells only 5% of the cells exhibited T antigen g, as could be expected from a smaller amountof recombinant virus in the virus stocks. The ElA gene products synthesized in the recombinant-virus-infected cells were labeled with [35S]methionine and analyzed by two-di-mensional gelelectrophoresis followed by auto-radiography. The amount produced was com-pared with that synthesized in Adl2-infected and -transformed cellsbyelectrophoresisof the cell extracts directly without immunoprecipita-tion(Fig. 6). A cluster ofpolypeptides (molecu-lar weight, 35,000 to40,000; pl, 5.0 to 5.5) was clearly visible in the extracts prepared from SV-AdElA-infected GC7 and CV1 cells(Fig. 6a and b). A much greater amount of thepeptides was detected in the infected GC7 cells than in the infected CV1cells,presumablydue to the differ-ence in the susceptibility of the cells to SV40 infection as stated above. These peptides were detectedat apositionsimilar tothat of the ElA polypeptides immunoprecipitated from the ex-tractofAdl2-infected HEKcells (Fig. 6d). No significant amount of the polypeptides was de-tected in the extracts from Ad12-infected KB cells(Fig. 6e) or from3YgE1Acellswhich were transformed with an Ad12 DNAfragment con-taining the ElA gene (Fig. 6f). No polypeptide wasdetected inuninfectedGC7 cells (Fig.6c).
DISCUSSION
The SV40vector used in thepresentstudy is
useful forcloning and forexpressionof aforeign gene of up toapproximately 1,900 bpinmonkey cells. Since the vector not only contains the initiation and termination signals for transcrip-tion butalso retains thesplicingjunctions down-stream from the late promoter, the gene to be inserteddoes not have topossess its own splic-ing junctions for expression. The Adl2 ElA gene with or without its own promoter was
insertedinto this vector toseewhether theElA
promoter is recognized in the vector. The
recombinant DNAs were propagated and pack-aged bycotransfection withSV40tsA58 DNAas
a helper in monkey cells. The ElA gene
prod-J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.489.55.241.68.245.2]VOL.45,1983~SV40RECOMBINANTS CARRYING Adl2 ElA GENE 415
a b
-130
640
510-3
450- ~
310-c d
-1180
-1090
1020- -1040
930-
- 950
790-0 (730)
(570)-310
B
-:'-- ,. / _:
[image:8.489.102.388.76.518.2]I
FIG. 4. Si nuclease mappingofAdl2 EtAmRNAfromrecombinant-virus-infected cells. (A)Cytoplasmic
RNAspreparedfromSV-AdElA(lanesaandc)-orSV-AdElAp(lanesb andd)-infectedCV cellsat56 hafter
infection. The RNAswerehybridizedwitha32P-labeled Adl2 EcoRIDNAfragmentC(0-16.5mapunits).The
hybridsweretreated withSinuclease andanalyzed byalkaline(lanesaandb)orneutral(lanescandd)agarose
gelelectrophoresis. The numbers indicate thelengthsof thefragmentsestimatedfrom thepositionsof the size
marker DNAfragments. (B)Expected lengths of the DNAfragments protected bythe RNAs.
ucts (T antigen g) were efficiently produced in
monkey cells after infection with the resulting
virus stocks,judgingfrom thestrongintensityof immunofluorescence in the nuclei. However,
theproportionofT-antigen-positive cells didnot exceed more than 20% of the cells, owingto a
small amount of the recombinant virus in the
virus stocks. Thegeneproducts werevery simi-lar or identical to those synthesized in Adl2-infected HEK cells, since they migrated to the
same spot upon two-dimensional electrophore-sis.
Si nuclease mapping of the hybrid mRNA transcribed from the recombinant genome
indi-A
I
VOL.
45,
1983.11.- i z r...
1- --1-.1 mmmmmmmmir
.k. ?
-V-I !l:
on November 10, 2019 by guest
http://jvi.asm.org/
ODA ET AL.
FIG. 5. Indirectimmunofluorescent staining ofrecombinant-virus-infected cells for T antigen g.
cated that the transcripts were initiated exclu-sively at the SV40 late promoter. The ElA promotercould not beutilized, presumably
ow-ing to its promoter activity being weaker than thatofthe SV40 late promoter under the condi-tionsused. Theexpression of thispromotermay be restricted to the early phase of infection or
may need more base sequences upstreamfrom the presumed ElA promoter, or both. In
con-trast,theElApolyadenylation
signal
was recog-nized. Althoughamajority
of thehybrid
mRNAwas terminated at the SV40
polyadenylation
site, a small butsignificant
amountofRNAwasterminatedatthe ElApolyadenylationsite. The hybrid RNA was correctly spliced in the ElA coding region, and nearly equalamounts ofAl and A2 mRNAspeciesweregenerated,aswould be the case in Adl2-infected human cells.
Translation of these hybrid mRNAs
tran-scribed from the SV-AdElArecombinant must
be efficient, since the ATG codonfor the ElA polypeptides is located
only
7bp
downstream from the 5' endoftheinsertedAdl2 ElA DNA. Thedistance between the upstream 3'splice
site and the ATG codon is therefore 40bp
and is almost thesame asthat between the 3'splicesite and the ATG codon for the VP1 (36bp) in 16SmRNA. This distance in the recombinant
SV-AdElAp was about 100 bp longer than that in SV-AdElA; however, the
efficiency
of transla-tion of the hybridmRNAs may notbeasdiffer-ent between SV-AdElAp and SV-AdElA
re-combinants, since the amount of the ElA polypeptides synthesized seemed tobe propor-tionaltothatof thehybridmRNAssynthesized.
Thehigher level of the E1A gene products in the SV-AdElA-infected cells as compared with the SV-AdElAp-infected cells may therefore be as-cribed to the higher proportion of the recombi-nantgenomein thevirus stocks. The level of the ElA gene products in the SV-AdElA-infected cells was also compared with that in KB cells infected withwild-type (WT) Adl2 and with that intransformed3YgElA cellsby two-dimension-al electrophoresis of [35S]methionine-labeled
cell extracts followed by autoradiography.
Un-der the conditions used, the dense spot of the ElAproducts was visible in the autoradiogram of the extract from SV-AdElA-infected cells, whereas no clear spot was detected in the
ex-tracts from KBcells infected with WTAdl2 or
in the extracts from transformed cells. The re-sults suggest that the level of the ElA gene products in SV-AdElA-infected cellsis at least 10-foldhigherthan that in the latter two types of cells.
The ratio of the recombinant to the helper virus in the virus stocks may be influenced by thefollowing factors. First, the sequence of the inserted DNA is critical for the replication or packagingof the recombinant DNA.SV-AdElA (4,470 bp) andSV-AdElAp(4,627bp)differ only in the sequences upstreamfrom the ElA coding sequence, whereas the amount of recombinant genome in the virus stocks was consistently lower with SV-AdElAp DNA than with SV-AdElA DNA. The base sequences presentonly in SV-AdElAp may therefore be inhibitory to replication or packaging of the recombinant DNA. It has been reported that a specific
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.489.88.408.74.284.2]SV40 RECOMBINANTS CARRYING Adl2 ElA GENE 417
~~~~~~~
e
_
-gal
FIG. 6. Two-dimensional gel electrophoresis of proteins from recombinant-virus-infected cells. Confluent
monolayersofCV1 (7 x 106)and GC7 (2x106) cells were infected with 1.0 ml of theSV-AdElA virus stock at 41
and39.5°C,respectively. The cells werelabeled with[35S]methionine for 2 h in methionine-free medium at the
times indicated below. Ascontrols, KB and HEKcells were infected withAdl2 at 10 and 50 PFU per cell,
respectively, in the presenceof 25 ,ug ofcytosine arabinoside per ml at37°Cand were similarly labeled with
[35S]methionineat 22and 12 hafterinfection, respectively. The cell extracts were prepared as described in the
text and subjectedto two-dimensional gelelectrophoresis. Each electrofocusing is from left (basic) to right
(acidic). Sincethesecond-dimension electrophoresis in panel d was carried out in a 13% gel instead of a 15% gel,
thepolypeptides migrated slightlyfaster than in theother panels. Panels: a, AdElA (GC7 cells, 36 h); b,
SV-AdElA(CV1cells, 56 h); c,uninfected GC7cells; d,Adl2 (HEK cells, 12 h) (proteins were immunoprecipitated
withanti-T-antigen-g serum);e, Adl2(KBcells,22 h); f, 3YgE1Acells.
VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.489.49.445.60.568.2]418 AL.
pBR322 DNA sequencehas aninhibitory effect onthereplication of pBR322-SV40 recombinant
DNA in monkey cells, and the recombinant DNAwhichefficiently replicated in the cells has
this sequence deleted (13). The EtA coding
sequences mayalsobe inhibitory topropagation
of therecombinant DNA sincean SV40
recom-binantcarrying the Adl2 EtBgene
(SV-AdElB-B) propagates very efficiently in monkey cells, as described in the following paper (22). Ithas
been suggested that 70 to 100% of full-length
SV40 DNAis suitable for packaging (19);
how-ever, the efficiency of packaging may differ
significantly within the lengths in thisrange.The
length of the recombinant SV-AdElB-B DNA (5,307 bp) is 101% of full length, whereas the recombinant SV-AdElA and SV-AdElAp DNAs correspondto85and 88% of full length, respectively. The upperlimit of the size is also
critical for packaging; a length 102% of full
length could notbe packaged (19).
Second, the susceptibility of monkey cells to
SV40 and the efficiency of transfection are im-portantfactorsforobtainingavirusstockwitha
higher ratio of recombinant virus, especially if the recombinant DNA is replicated and pack-aged inefficiently as compared with the helper
DNA. We consistently observed that the virus stocksprepared with GC7 cells contained higher ratios of the recombinant virus than those
pre-pared with CV1 cells. As previously described, GC7cellsare moresusceptibletoSV40 thanare
CV1cells;togetT-antigen-positive cellsinmore
than 90% of the cells by infection withSV40, a
multiplicity of 5 PFU per cell is sufficient for
GC7 cells, whereas CV1 cells require a
multi-plicity of 75 PFUpercell(36).Ahigher
efficien-cyoftransfection of the recombinantDNAwas
always observed by exposure of the cells to
DNA in medium containing 200 ,ug of DEAE-dextran permlfor 8 h thanbytheconventional method in which the cells were exposed in
mediumcontaining 500 ,ug of DEAE-dextranper
mlfor 30 min. Efficient transfection and
infec-tion may minimizethe number of infection
cy-clesrequired forthedevelopmentofa
cytopath-iceffect in the cells and avoid the decreasein the
ratio of recombinantto helpervirus.
Finally, theuseofanearlydeletionmutantof
SV40 instead of a point mutant as helper may
also facilitate the preparation of virus stocks
with higher ratios of recombinant virus. The
plaques formed after transfection of monkey cells with the recombinant plus SV40 tsA58
DNAs containedWT SV40 in aminor
propor-tion. The results suggest that the virus stocks
prepared after development of the cytopathic effect contained a small but significant amount
of WT SV40. A population of WT SV40 may
increase upon successive passages in culture.
The probability ofgeneration of WT SV40 by recombination between the recombinant and helper DNAs may be lowered if the mutant which deleted most of the SV40 early region is usedashelper.Ithas been recently reported that the helper DNApSVL5, in which only the early region between the origin and the TaqI site (about 500 bp) is retained, efficiently replicates in Cosl cells (4), giving rise to mature virions (9).
The ElA gene of adenovirus is involved in both the activation of other early genes in the lytic infection cycle and the transformation of susceptible cells. In spite of these interesting multiple functions, no biological activity of the gene products has been investigated in vitro. Efficient production ofthe
EtA
gene products in monkeycellswill facilitatetheir purificationand theanalysis for biologicalactivities in vitro.ACKNOWLEDGMENTS
Wethank P.Berg and H.Okayama for valuablesuggestions andadvice.Cloning of the SV40 DNA segment containing the splicing junctions for late 16S RNA was carried out in collabo-ration with H. Okayama when K.O. was at P. Berg's labora-tory atStanford University.
This work was supported by grants from the ministry of Education, Science, and Culture of Japan and by grants from theMitsubishi Foundation.
LITERATURECITED
1. Berk, A. J., F. Lee,T.Harrison, J. Williams, andP. A.
Sharp. 1979. A pre-early adenovirus 5 gene product regulatessynthesis of early viral messenger RNAs. Cell 17:935-944.
2. Berk,A.J., andP.Sharp. 1978.Spliced early mRNAsof simian virus 40. Proc.Natl. Acad. Sci. U.S.A.
75:1274-1278.
3. Gething, M.-J., and J. Sambrook. 1981. Cell-surface expression of influenza haemagglutinin from a cloned
DNAcopyof theRNAgene. Nature(London) 293:620-625.
4. Gluzman,Y.1981.SV40-transformed simian cells support thereplication of early SV40mutants.Cell 23:175-182.
5. Gruss, P., R. W. Ellis, T.Y. Shih, M. Konig, E. M.
Scolnick,andG.Khoury. 1981. SV40 recombinant mole-culesexpress the geneencodingp21transformingprotein
of Harvey murine sarcoma virus. Nature (London)
293:486-488.
6. Gruss, P., and G. Khoury. 1981. Expression of simian virus 40-ratpreproinsulin recombinants in monkey kidney cells:useofpreproinsulinRNAprocessingsignals.Proc.
Natl.Acad.Sci. U.S.A. 78:133-137.
7. Hamer,D.H.,M.Kaehler,and P.Leder.1980.A mouse
globin gene promoter is functionalinSV40. Cell 21:697-708.
8. Harter,M.L.,andJ.B.Lewis.1978. Adenovirustype2
early proteinssynthesizedin vitroand invivo: identifica-tionin infectedcellsofthe38,000- to 50,000-molecular-weight protein encoded by the left endof the adenovirus type2genome. J.Virol. 26:736-749.
9. Hartman, J. R., D. P. Nayak, and G. C. Fareed. 1982. Human influenza virus hemagglutinin is expressed in
monkey cellsusing simian virus40 vectors. Proc. Natl.
Acad. Sci. U.S.A. 79:233-237.
10. Hirt,B.1967.Selective extraction ofpolyomaDNAfrom infected mousecellcultures.J.Mol. Biol.26:365-369. 11. Houweling,A.,P.J.van denElsen,and A.J.van derEb.
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 RECOMBINANTS CARRYING Adl2 ElA GENE 419
1980.Partialtransformation ofprimaryratcellsby the left most 4.5% fragment of adenovirus 5 DNA. Virology 105:537-550.
12. Jones, N., and T. Shenk. 1979.Anadenovirus type 5early genefunction regulates expression of otherearly viral genes. Proc. Natl.Acad. Sci.U.S.A. 76:3665-3669. 13. Lasky, M., and M. Botchan. 1981. Inhibition ofSV40
replication in simian cells by specificpBR322 DNA
se-quences.Nature(London) 293:79-81.
14. Maruyama, K., T.Hiwasa,and K.Oda. 1981. Character-izationof flatrevertantcells isolated from simian virus 40-transformed mouse andratcellswhich containmultiple copies of viral genomes.J.Virol. 37:1028-1043. 15. Moriarty, A. M., B. H. Hoyer, J. W.-K. Shih, J.L.Gerin,
and D.H. Hamer. 1981. Expression of the hepatitis B virus surfaceantigen gene in cell culturebyusingasimian virus 40 vector. Proc. Natl. Acad. Sci. U.S.A. 78:2606-2610.
16.Mulligan, R.C., and P. Berg. 1980. Expression of a
bacterial gene in mammalian cells. Science 209:1423-1427.
17. Mulligan,R.C., andP.Berg. 1981. Selection for animal cells that express the Escherichia coli gene coding for xanthine-guanine phosphoribosyl transferase. Proc. Natl. Acad.Sci. U.S.A. 78:2072-2076.
18. Mulligan, R. C., B. H. Howard,andP.Berg. 1979.
Syn-thesisof rabbit,3-globinin culturedmonkeykidneycells following transfection withaSV40 3-globinrecombinant genome. Nature(London) 277:108-114.
19. Muzyczka, N. 1980. Construction of an SV40-derived cloningvector.Gene 11:63-77.
20. Nevins, J.R.1981.Mechanism of activation ofearly viral transcription by the adenovirus ElA geneproduct. Cell 26:213-220.
21. Nevins, J. R., H.S. Ginsberg, J.-M. Blanchard, M.C. Wilson, and J. E. Darnell, Jr. 1979. Regulation of the primary expression of the early adenovirustranscription
units. J. Virol. 32:727-733.
22. Oda, K.,Y.Fukui,I. Saito,M.Masuda,K.Shiroki,and H.Shimojo. 1983.Hyperproductionof adenovirus type 12 E1B geneproductinmonkey cells,usingasimian virus40
vector. J. Virol. 45:420-427.
23. O'Farrell, P. H. 1975. High resolution two-dimensional
electrophoresisofproteins.J.Biol. Chem. 250:4007-4021.
24. Pavlakis, G. N.,N. Hizuka,P. Gorden,P.Seeburg, and D.H. Hamer. 1981. Expression oftwo human growth
hormone genes in monkey cells infected by simian virus 40recombinants. Proc. Natl. Acad. Sci. U.S.A. 78:7398-7402.
25. Persson, H., H.-J. Monstein,G.Akusjarvi, and L.
Philip-son. 1981. Adenovirus earlygene products may control viral mRNA accumulation and translation in vivo. Cell 23:485-496.
26. Rigby, P. W. J., M. Dieckmann, C. Rhodes, and P. Berg.
1977. Labeling deoxyribonucleic acid to high specific activity in vitro by nick translation with DNA polymerase I.J. Mol. Biol. 113:237-251.
27. Ross, S. R., S. J. Flint, and A. J. Levine. 1980. Identifica-tion of the adenovirus early proteins and their genomic map position. Virology100:419-432.
28. Saito, I., J. Sato, H.Handa, K. Shiroki, and H. Shimojo. 1981.Mapping of RNAs transcribed from adenovirus type 12early and VA RNA regions.Virology 114:379-398. 29. Shiroki, K., K. Segawa, and H. Shimojo. 1980. Two tumor
antigens and their polypeptides in adenovirus type 12-infected and transformed cells. Proc. Natl. Acad. Sci. U.S.A. 77:2274-2278.
30. Shiroki, K., H.Shimojo,Y. Sawada, Y. Uemizu, and K. Fujinaga. 1979. Incomplete transformation of rat cells by a smallfragment of adenovirus 12 DNA. Virology 95:127-136.
31. Sompayrac, L. M., and K. J. Danna. 1981. Efficient infec-tion of monkey cells with DNA of simian virus 40. Proc. Natl. Acad. Sci. U.S.A. 78:7575-7578.
32. Subramani, S., R. Mulligan, and P. Berg. 1981. Expres-sion of the mouse dihydrofolate reductase complementary deoxyribonucleic acid in simian virus 40 vectors. Mol. Cell.Biol. 1:854-864.
33. Sugisaki, H., K. Sugimoto, M. Takanami, K. Shiroki, I. Saito, H. Shimojo, Y. Sawada, Y. Uemizu, S. Uesugi, and K.Fujinaga.1980.Structureand gene organization in the transforming HindIII-G fragment of Adl2. Cell 20:777-786.
34. Tooze, J. 1980. DNA tumor viruses. Molecular biology of tumorviruses, part 2. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.
35. Vogelstein, B., and D. Gillespie. 1979. Preparation and analytical purification of DNA from agarose. Proc. Natl. Acad. Sci.U.S.A. 76:615-619.
36. Yamamoto, K., N. Yamaguchi, and K. Oda. 1975. Mecha-nismof interferon-induced inhibition of early simian virus
40(SV40)functions. Virology 68:58-70.
VOL.45,1983


![FIG. 6.andAdElAthemonolayerswithtexttimesrespectively,(acidic).[35S]methionine Two-dimensional gel electrophoresis of proteins from recombinant-virus-infected cells](https://thumb-us.123doks.com/thumbv2/123dok_us/1451384.97698/10.489.49.445.60.568/andadelathemonolayerswithtexttimesrespectively-acidic-methionine-dimensional-electrophoresis-proteins-recombinant-infected.webp)