Human
Immunodeficiency Virus
Infection of Monoblastoid
Cells:
Cellular Differentiation
Determines
the Pattern
of Virus
Replication
C. DAVID
PAUZA,1*
JOSEGALINDO,'AND DOUGLAS D. RICHMAN2Developmental Biology Laboratory, the Salk Institutefor Biological Studies, P.O. Box85800, SanDiego,
California
92138-9216,1
andDepartments of PathologyandMedicine, University ofCalifornia,and Veterans Administration MedicalCenter, San Diego, California 921612
Received 21 March 1988/Accepted 27 June 1988
Stringent control of human immunodeficiency virus (HIV) replication was observed in the human
monoblastoid cell line U937. A low-multiplicity infection of these cells by the LAV1 strain of HIV was
productive for 2.5 days; then virus replicationbecame restricted andnofurther evidence of virionproduction
was observed. The dramatic decrease in HIV production was due in part of reduced accumulation of
cytoplasmic viralRNA and occurred in the absence of evident cytopathic effects. In contrast, infected cells
inducedtodifferentiate by phorbolester, vitaminD3, orlymphokinesupernatant did not releasemarkers of
HIVdespite the accumulationofsignificant levelsofcytoplasmicviral RNA.HIVinfectionalteredthepattern
ofc-mycRNA accumulation in U937 cells.Expressionof thisgenechanges normallyinresponseto the stateof cellulardifferentiation; ininfected cells the level ofc-mycexpression wascorrelatedto the levels of viralRNA
accumulation and not to cellular differentiation. These results suggest that restricted replication of HIV in
monocytesmightbeanimportantmechanism of virus persistence and demonstratearelationshipbetweenHIV
replication and monocyte differentiation.
Accumulating evidence indicates that infection of
mo-nocytes by human immunodeficiency virus (HIV) is an
importantcomponentinthepathogenesisofacquired
immu-nodeficiency syndrome (AIDS). HIV-infected individuals
manifest elevated levels of circulating lysozyme (19), which
is released by activatedmonocytes (8, 18); increasedserum
lysozyme levels are correlated with progression to AIDS
(19). Monocytes derived from AIDS patients have
signifi-cantly reduced phagocytic (9) and chemotactic (35, 40)
activities, and thepercentageofthese cells thatareHLA-DR
positive is approximately halfthe level observed normally
(42). The monocytephenotypes observed in AIDS patients
suggest thatactivation and differentiation of these cells are
altered byHIVinfection.
In addition to the phenotypic alterations observed in
monocytepopulations derived from AIDS patients, evidence
exists that these and related cells aredirectly infected with
HIV. Viral RNA and antigens have been observed in brain
tissue macrophages (14, 26, 43) and in the macrophagelike follicular dendritic cells inlymph nodes (2, 3, 31). Cellswith
macrophage markers inthe skinand lungs of AIDS patients
have been shownto contain infectious virus, viral antigens,
or viral nucleic acids (6, 23, 31, 34). Peripheral blood
monocytes arepermissive forHIVin vivo(15, 23, 30), and
virus can be isolated from these same cell populations in
infected individuals (15, 23). HIV infection ofmonocytes,
and the variety of specialized cell types related to this
lineage, constitutes a significant component of virus
distri-bution inthe infected individual.
The role of monocytes in AIDS is controversial. It has
been suggested that these cells serve as the reservoir for
virus in the body (22, 25). The observation oflow ratesof
HIV replication in monocytes in vitro and the absence of
cytopathic effects on these cells has beenproposed as one
possible explanation for HIV persistence in vivo(15, 21). It
hasalso been proposed(33) thatinfectedmonocytesserveas
*Corresponding author.
theprincipalagents of HIV disseminationto helperTcells,
because thesetwo cell types necessarilycome into contact
during the course of antigen presentation and the T cell
receives activation signalsinconsequenceofthis interaction
(33). New insights into the role of monocytes in AIDS are
especially important in light of the recent discovery that
dideoxynucleosides, which constitute animportant class of
agents for the treatment ofAIDS, have reduced antiviral
activityin cells of thislineage (39).
The interaction between HIV and the host monocyte is
thoughttobecomplex, involving specialmechanisms for the
control of virus production. Accordingly, we sought to characterize thebiologyof HIVreplicationinmonocytes. In these initial studies,the humanmonoblastoid cell line U937 (41)wasusedas amodel forHIV-monocyteinteractions and
the relationship between virus production and cellular
dif-ferentiation. Weobservedthat HIVreplicationin these cells
was regulated stringently. A low-multiplicity infection of
U937 cellswas productive initially. At 2days postinfection,
we observed that cytoplasmic viral RNA, cell-free reverse
transcriptase activity,andp24gagantigenlevels diminished
rapidly withoutevidentcytopathic effects. Thus, productive
infection had become restrictedand virus release ceased. In contrast, infected U937 cells induced to differentiate by treatmentwithaphorbolester(tetradecanoylphorbolacetate
[TPA]), 1,25 dihydroxyvitaminD3,orlymphokine
superna-tants did not release markers of HIV despite significant
accumulation ofviral RNA inthe cytoplasm of TPA-treated
cultures.
Direct effectsof viralinfection on the cellular phenotype
were assessed by examining the cellular gene c-mycthatis
expressed normallyinU937 cells. Thepatternofc-mycRNA
accumulation was altered in infected cells as opposed to
uninfectedcontrols, attestingtoadirect effect ofHIVonthe
phenotype ofthese cells. Thealtered patternofc-mycRNA
accumulation in infected cells indicates that these changes
are mediated at least in part by direct effects of HIV on
cellulargene expression.
3558
0022-538X188/103558-07$02.00/0
Copyright C 1988,American Society forMicrobiology
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Cellculture andpreparation of virusstocks. U937 cells (41)
weremaintained at densities of between 1 x
105
and 5 x 105cells per ml in RPMI 1640 medium with 10% heat-inactivated
fetal bovine serum, supplemented with
penicillin-streptomy-cin-glutamine. The cells were passaged on the day of before
infection. Cell-free stocks of the
LAV1bl,
strain of HIV (1)wereprepared in the T-lymphoblastoid cell line, CEM. The
titers were adjusted to 107 tissue culture infective doses
(TCID50s)
per ml, asdetermined by syncytiumformation inaterminal dilution assaywith MT-2 cells(20).
Virus production in differentiated and undifferentiated
U937cells. Cultures of U937 cells were infected at a
multi-plicity of 0.2 TCID50 per cell. At 8 h later, the culture was
split and TPA was addedto half, at a final concentration of
1 ,ug/ml. After 1 h, cells were recovered from both cultures
bycentrifugation, washed once inphosphate-bufferedsaline
(PBS), and suspended in fresh medium to a final
concentra-tion of 5x 105cells perml. The flasks, each containing 20 ml
of culture, were then incubated at 37°C in an humidified
incubator with 5% CO2 in air and were harvested at 0, 1, 2.5,
and 5 days after TPA treatment. The cultures were
supple-mented with an equal volume of fresh medium at 2.5 and 4
dayspostinfection. The effects of 1,25 dihydroxy vitamin D3
or lymphokine supernatant treatments were assessed in a
similar manner; vitamin D3 was added to infected cell
cultures at a final concentration of
10-9
M (provided byMilan Uskokovick, Hoffmann-La Roche Inc.), and
commer-cially available delectinated supematant form
phytohemag-glutinin-activated human peripheral blood lymphocytes
(Ad-vanced Biotechnologies) was added as a 20% volume
supplement to infected U937 cells in culture.
At each time point, the cells were collected by
centrifu-gation. A 1-ml sample of the cell-free supernatant was
retained and stored at -70°C for assay of reverse
transcrip-tase activity and quantitation of p24 gag antigen (p24
en-zyme-linked immunosorbent assay, Abbott Laboratories,
Chicago, Ill.). The flasks were rinsed with 5 ml of PBS
containing 0.5 mM EDTA to recover adherent cells. The
harvested cells were washed again with cold PBS and
suspended in 2.5 ml of 15 mMNaCl-15 mM Tris
hydrochlo-ride (pH 7.5). Then, 100,ul of 20 mM vanadyl ribonucleoside
and 200 ,lI of 20% Nonidet P-40 were added. Nuclei were
removed by a2.5-min spin at 2,500 rpm in a Beckman TJ-6
centrifuge, and the supernatant was combinedwith anequal
volume of 150 mM NaCl-150 mM Tris hydrochloride (pH
7.5)-i
mM EDTA. Proteinase K (Boehringer Mannheim)was then added to a final concentration of 200 ,ug/ml.
Samples were incubated for 45 min at 56°C and then
ex-tracted with an equal volume of 1:1 phenol:chloroform
containing 0.1%hydroxyquinoline-0.5% 2-mercaptoethanol.
The aqueous phases were recovered, and RNA was
precip-itated by the addition of sodium acetate (pH 5) to a final
concentration of 0.3 M plus 2.5 volumes of ethanol (10).
Samples were stored at -20°C
overnight,
and the nucleicacid precipitates were recovered by centrifugation.
The RNA pellet was purified further to remove
contami-nating vanadyl ribonucleoside (10). The dried
pellet
wassuspended in 250 ,u of3 M sodium acetate
(pH
5), and theRNAwas recoveredby brief
centrifugation
inanEppendorf
microcentrifuge. The pellet was rinsed in 70%
ethanol,
suspended in water, and precipitated again. The
resulting
material was of sufficient purity that the concentration of
RNA could be determined from the optical
density
at 260nm. Samples of eachpreparation werealso runon agarose
gelsas an additional check on RNA contentand integrity.
The reversetranscriptaseactivity of cell-freesupernatants
wasdetermined by amodification-ofthe methodofPopovic
etal. (36). Aftera2-hincubation at37°Cin thepresenceof
5 mM
MgCl2-poly(rA)
template-oligo(dT) primer, with[32P]dTTP as label, portions ofthe reaction mixture were
spottedonDE81 paper andallowedtodry
briefly.
The paperwas washedtwiceat roomtemperature in 2x SSC (1x SSC
is0.15 MNaCland 0.015 M sodium citrate), rinsedoncein
95% ethanol, andthenexposedtofilm for3 to 5 h. Individual
sections werethencutout,andthe
incorporated
radioactiv-itywasdeterminedby Cerenkovcounting.
Assays for cellular and viralgene expression.Cytoplasmic
RNA sampleswereanalyzedtodeterminethe relative
con-tents of c-myc, HIV, and actin RNA. The assays were
performed by dotblotting ofthecytoplasmicRNA samples
onto Schleicher & Schuell, Inc., nitrocellulose filter
mem-brane and hybridization with the appropriate radiolabeled
probe.
Samples for dot blotting were made 25 mM
morpholine-propanesulfonate (pH
7.0)-5
mM sodium acetate-5 mMEDTA (lx MOPS buffer-S50% deionized formamide-3.7%
formaldehyde
(vol/vol)
in a final volume of 250 ,ul. Thesampleswere incubatedat
56°C
for20min and thenchilled briefly. Nitrocellulose membrane that had been immersed previouslyinwaterandagain
in20x SSCwasplaced
inthedot-blotting
manifold;
then60-,ul
portions
of eachsample
were placed inwells, and theliquid wasdrawnthroughthemembrane with gentle vacuum. The membrane was then
rinsed oncein2x
SSC,
baked at80°C
for2 h ina vacuumoven, and stored until
hybridization.
Radiolabeled probe for
expression
ofthec-myc gene wasgenerated
by
primer
extensionlabeling (11)
of a1.0-kilobase-pair
(kb)
PstIfragment
ofthe human c-myc gene(37). The radielabeledHIV probes were
prepared
by
using
the9.5-kbfragment
oftheplasmid pARV-2 (27),
and actin probes were derived from the 0.6-kb PstIfragment
of amurine
,-actin
cDNA clone(28).
Prehybridization
andhy-bridization of the membranes were described
previously
(32).
Analysis of proviral genomes. Pellets
containing
infectedcell nucleiwererecovered
subsequent
toNonidet P-40lysis
and
suspended
in 2.5 ml of25 mM Trishydrochloride
(pH
7.5)-12.5 mM EDTA-140 mMNaCl-0.5% sodium
dodecyl
sulfate; proteinase
K was added to afinal concentration of200
,xg/ml,
and thesamples
were incubated at56°C
for 30 min.Then,asecondportion
ofproteinase
Kwasadded,
andthe incubation continued for an additional hour before the
sample was extracted with an
equal
volume of buffered phenol solution. The aqueousphase
was collected anddialyzed
extensively against
10 mMTrishydrochloride
(pH
7.5)-i
mM EDTA. The DNAconcentration was calculated fromtheoptical
density
at260nm.Purified cellular DNA was
digested
with the restrictionendonuclease
HindIII,
and theresulting fragments
wereseparated by
agarosegel
electrophoresis.
The DNA wastransferred to
Hybond
filter membrane(Amersham
Corp.);
hybridization of the filter membrane was
performed
asdescribedfor the RNA
analysis.
RESULTS
HIVproduction isrestricted in U937 cells. The
replication
of HIV in U937 cells was evaluated
by
measuring
theaccumulated reverse
transcriptase
activity
andp24
gagon November 10, 2019 by guest
http://jvi.asm.org/
C-)
0
Lfl
z
0 2 5
(A)
0-C)
or.
b z ,
ooz C o
~ x
m c>,
z =
L .>
-500 e 2 0
LU4
_ E6
0
z DAYS
FIG. 1. LogarithmicallygrowingU937 cellswere infectedwith 0.2TCID50 per cell of LAVI (passaged in CEMcells).The culture was split, andhalf was treatedwith TPA and washedasdescribed in Materials and Methods.Infected cellswerethenplatedat adensity of 5x 105/ml. Sampleswereremovedimmediately (day0) anddaily for the remaining 5 days and assayed for reverse transcriptase activity (36). Circles denotereversetranscriptaseactivity data,and triangles designate p24antigenmeasurements;opensymbolsdepict the datafor undifferentiated U937cellcultures, and solidsymbols represent TPA-treated cell cultures. The data shown are from a
single experiment (the same culture that was used for RNA and DNAanalyses shownsubsequently) andarehighlyrepresentativeof more than15repetitions.
antigen content of infected-cell supernatants. U937 cells were infected with 0.2TCID50percell; newly synthesized
virus was released intothemedium ondays 1and 2
postin-fection. Thereafter, virus production
decreased
rapidlyandremained at a low level for the duration of the 5-day experiment (Fig. 1). The decrease in supernatant reverse
transcriptase activity was due toan effecton virus produc-tion and not to a direct inhibition ofreverse transcriptase activity. Admixing experiments, in which virus stocks of
known activity and culture supematants were combined, showed noinhibition of enzymatic activity.
Theeffects of phorbolester treatment onHIVreplication
in U937cellswereevaluatedin asimilarmanner. Cellswere
preinfected ina single flask andthensplit intoTPA-treated and untreated samples. Preinfection was required because
TPA treatment of U937 cells rendered them
resistant
toinfection (7; C.D.P., unpublished data). Infected phorbol ester-treated cells did not release detectable reverse
tran-scriptase orp24 gag antigen (Fig. 1). The results on HIV
production by normal and differentiated U937 cells were
obtained
consistently
in more than 15 independentexperi-ments.
We also assessed the accumulation ofreverse
transcrip-taseactivity inthemedium ofinfectedcells that were treated
with either1,25dihydroxy vitaminD3 ordelectinated
super-natantfromphytohemagglutinin-activated human peripheral
blood lymphocytes. Both treatments are known to induce U937 differentiation (38). In three separate analyses of the
effects of both ofthese treatments, the release of reverse
transcriptaseactivity was similar to that observed with TPA
treatment (Fig. 2). Three
distinct
agents known to induceU937 cellulardifferentiationthus were able toreduce
signif-icantly the release of HIV from infected cellcultures. The abrupt decline in virus production manifested by
undifferentiated U937 cells was not attributable to cell
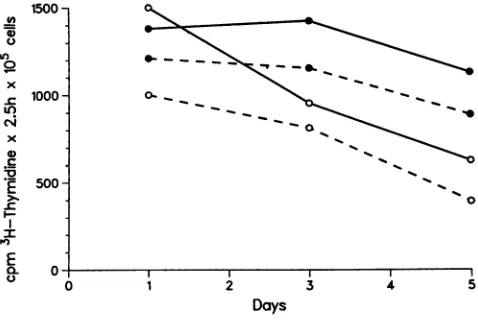
tox-icity. Nosignificantdifferences in the rates of DNA
synthe-sis were apparent between the infected and uninfected cell
culturesasdeterminedby the rates of[3H]thymidineuptake
(Fig. 3). The percentage of viable cells as determined by
0 3
DAYS
4 5 6
FIG. 2. Effects ofTPA, 1,25dihydroxy vitamin D3, and PHA-activatedlymphocytesupernatantsontheaccumulation of cell-free reverse transcriptase activity in infected U937 cultures. Infection and cellculturewere asdescribed forFig.1. Thereverse transcrip-taseactivity data arerepresented by theheight of theappropriate bars, and the range of values among threeindependentexperiments is shown by the error bars. Within each group offour bars, the leftmost open bar represents untreatedcells,thehatched bar
desig-natesTPA-treatedcells,the solid bar shows vitaminD3effects
(10-'
Mfinalconcentration), and therightmosthatched bar documents the effect ofadding activated lymphocyte supernatants at 20% final volume.
trypan blue dyeexclusionwasgreater than90% in both the
infectedand uninfected cultures. Somecell death observed
in the phorbol ester-treated cultures was attributable to
chemicaltoxicity.
Viral RNA and provirus accumulation. Levels of HIV
RNAinthe
cytoplasmic
RNAfraction
wereroughly
corre-lated to the levels of virus
production
in undifferentiated U937 cells. Accumulation ofHIV-specific cytoplasmic
RNAwas observed on days 1 and 2
postinfection;
the RNA decreased in abundanceonday
5when virusproductionwasalso atits lowest
(Fig.
4). This correlationbetweencytoplas-mic RNA levelsand virus
production
was notobserved in the differentiated cellcultures. Differentiated(TPA-treated)
U)
In
0
x
C-c
x
0)
C
E 0.r C)
1500
-1000
-500
-_0_-_
0
0 1 2 3 4 5
[image:3.612.60.291.57.197.2]Days
FIG. 3. [3H]thymidine uptake in infected and uninfected U937 cell cultures. [3H]thymidine (1 ,uCi) was added to 200 ,ul of cell culture;2.5hlater the cells were recoveredbyfiltration and washed with several volumes of 0.9% NaCl. The data are expressed as countsperminute of[3H]thymidinetaken upin 2.5 hby 105 cellsfor eachday of cell culture. Solid lines representthe uptakerates for uninfected U937cells, andthebroken linesshowthecorresponding ratesfor infected cells. Solid circles denote untreatedcultures, and opencircles mark the cultures treated for 2 h with TPA.
I1,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.319.546.79.209.2] [image:3.612.315.554.490.650.2]A
untreatedI
I
is*
.4TPA-treated
I
I1
*
-ff10.l pg2 pg 0.5 pg
B if
[0
*b..infected *-
to
-0
*-0
10 pg
2 pg 0.5 pg
10 pg *
*
2pguninfected
1 2 4 5 1 2 4 5
[image:4.612.67.296.76.307.2]days post-infection
FIG. 4. Cytoplasmic RNAdot blots showing the levels of HIV (A) and actin(B) RNA in untreated and TPA-treated U937 cells. The blotswerepreparedasdescribed inthetextandhybridized witha radiolabeled9.0-kb DNAfragment ofthe clonedgenomeof the HIV
isolateARV-2 (27)or anEcoRIfragment ofmouse actin.In panel A, only the samples frominfected cells areshown. Therewas no detectable hybridization to uninfectedcell RNA. In panel B, the
uppersectionshows theactinRNAcontentof uninfected cells and the lower panel shows the actin content of infected cells. The
amountoftotalcytoplasmicRNAperdot isshownatthe right, and thedaypostinfection is shownatthe bottom. These RNA samples
werepreparedfromthesamecell cultures represented in Fig.1.No
RNAwasloaded in the day2,uninfected TPA-treated lane, and this
accountsforthe absence ofhybridizationatthis position.
cells contained significant amounts of cytoplasmic viral
RNAeventhough viralreversetranscriptase and p24 antigen
were notreleased into the medium. The level of viral RNA
remained constant in differentiated U937 cells over the
course of the experiment. Hybridization with the mouse
,B-actingeneprobe demonstrated that equalamountsof RNA
had been applied to all positions of the filter and that the
levels of actin RNA were not affected significantly by
infection or differentiation (Fig. 4). Similar results were
obtained inthree additionalexperiments.
Therelative accumulation of HIVprovirus was assessed
by DNA blot analysis (Fig. 5). The hybridization with
samples from days 2.5 and 5 postinfection indicated a
relativelyconstant amountofproviral DNA. In this
partic-ularexperiment, the sample in lanebcontained lessDNA,as
judged by ethidium bromide staining of the gel, and this
probably accountsfor the difference in hybridization
inten-sity between lanes b andc.Therewaslittle difference in the
relative amounts of the provirus between differentiated and
undifferentiated infected cell cultures, and this result was
consistent in three separate infection experiments.
There-fore, neither the decrease in virus production during
re-stricted replication nor the absence of particle release in
differentiated cell cultures could be attributed to selective
depletion of the infected-cell population.
Expression ofthec-mycgeneininfected U937 cells.
Expres-sion of thec-mycgene isamarker ofcellular differentiation
in U937 cells (28). Accordingly, we examined the
accumu-a b c d e
5.0 kb
[image:4.612.379.500.77.136.2]Vn- 2.5 kb
FIG. 5. Southern blot analysis of HIV proviral DNA in untreated and TPA-treatedU937 cell cultures. Nuclear DNA was prepared as described in the text, digested with the restriction enzyme HindIll, andfractionated on an0.8% agarose gel. After transfer to nitrocel-lulose andhybridization with a probe from the plasmid pARV-2 (27),
the hybridization pattern of the provirus was revealed. Only an
upper section of thehybridization pattern is shown. Lanes contain the following samples: a, uninfected U937 cell DNA; b, day 2, infected, no TPA; c, day 5, infected, no TPA; d, day 2, infected, TPA-treated; and e, day 5, infected, TPA-treated. The sizes of the bandsareshownatthe right.
lated levels of cytoplasmic c-myc RNA in infected and
uninfected cells, either with or without exposure to TPA
(Fig. 6). The dot blots wereprepared by using the same RNA
samples shown in Fig. 4, thus direct, qualitative comparison
betweenrelative levels of HIV and c-myc RNA is possible.
A nearly constant level of c-myc RNA was observed in
undifferentiated cells; the relative amountdeclined rapidly
after TPA treatment. This finding is in agreement with the
results reportedpreviouslyby Mitchell et al. (28) and attests
to therelationshipbetween c-myc RNA levels and the state
of monocyte differentiation. Infection by HIV altered
signif-icantly the patternof c-myc gene expression in U937 cells.
Instead ofremaining constant in the undifferentiated cell
cultures,c-myc RNA declinedsteadilyand inparallelto the
decreasing level of HIV RNA in these infected but still
viable cells. It is important to note that the decrease in
relative abundanceofc-myc RNA shown in Fig. 6 is most
likelyanunderestimate of the actualmagnitudeof change in individual infected cells because the RNA samples in these
experiments wereprepared from cell cultures composed of
approximately equal numbers of infected and uninfected
cells as determined by indirect immunofluorescence (not
shown). Therefore, the apparent change in c-myc RNA
probably reflects a larger decline in abundance of this
untreated
TPA-treated
*s,*. *gm1o10
pg
infected
L
*
-
2
pg
**@S@.*OO2*g0.5,ug
@0
0---
e
410
Pg
uninfected
*
**
a2 pg
L .
,
0.5
pg
1 2 4
5
1 2 4 5days post-infection
FIG. 6. Relativecontentof c-mycRNAin infected and uninfec-ted U937 cells withandwithout TPAtreatment.Theupper section shows the levels of c-myc RNA in infected cells, and the lower sectionshows thelevelsinuninfected cells. The RNAsamplesare
identical to thoseused in Fig. 4, thus the ,-actin control can be
compared directlywith thec-mychybridizationpattern.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.319.561.519.672.2]species
in infected cellssuperimposed
on thenormally
constant RNA levels in uninfected cells. We have not yet
examined the
pattern
of c-myc geneexpression
in cellcultures
containing
100% infected cells because athigher
multiplicities
ofinfection the property of restrictedreplica-tion isnotobserved
(C.
D.Pauza,
unpublished).
Incontrast tothe undifferentiatedinfectedU937cells,
the abundance ofc-myc RNA remained
nearly
constant in the infectedTPA-treatedcells
throughout
the5-day
timecourse. Inthiscase,the altered
regulation
ofc-myc RNAaccumulation isespe-cially
apparent. The uninfected differentiated cell cultureswould
normally
beexpected
to contain little or no c-mycRNA after 2
days
of TPA treatment.Thus,
the relativeabundance ofc-myc RNA in theinfecteddifferentiated cells was
clearly
evident.On the basis of these
experimental observations,
weconcludethat HIV infection altered the
regulation
ofc-myc RNA accumulation.Surprisingly,
a correlation betweenHIV and c-myc RNA levelswas observed. This is different
from thepatternofc-myc
expression
observed inuninfectedcells. Altered
expression
ofthe c-myc genewasobservedinfour separate infection
experiments;
in each case, thechanges
in c-myc RNA werequalitatively
similar to thepatterns ofviral RNA accumulation. Thisresult appears to
be a
specific
effect ofviral infection becauseexpression
of the actin genewasnotaffectedby
HIVinfection.DISCUSSION
HIV
replication
in U937 cells isproductive initially
andthenbecomes restricted.
During restriction,
thelevelofviral RNA in thecytoplasm
of infected cells decreasessubstan-tially.
Expression
ofthe c-myc genealso decreasesconcom-itantly
with viral geneexpression.
DNA blotanalysis
showedthat thesameamountsof
proviral
DNAper cellarepresent
during
bothproductive
and restrictedreplication;
thisobservation attests tothefactthat the
changes
in RNAlevels indicate altered accumulation of these
species
inindividual cells andnotselection
against
asubpopulation
of infected cells.Furthermore, thymidine uptake
rates forinfected and uninfected cells are
comparable, indicating
furtherthat thealteredbehavior of infectedcellsisnotdueto
a
cytopathic
effect ofthe virus.Consequently,
thestringent
control of HIV
replication
in U937 cells is an intrinsic attribute of the interaction between the monoblastoid celland the virus.
Infected U937
cells,
induced to differentiateby phorbol
ester, release reduced amounts ofHIV. In contrast to the
restricted
replication
observed in undifferentiated U937cells,
the differentiated cellswerenonproductively
infecteddespite
accumulation ofsignificant
levels ofcytoplasmic
viralRNA. Inthiscase,
high
levels ofc-myc RNA werealsoshowntobe present
during
differentiationof infected cells incontrast to the
rapid
decline in abundance of this RNAobserved in differentiated uninfected cells. Infected U937
cells were also induced to differentiate
by
the addition of1,25
dihydroxy
vitaminD3
(4, 5)ordelectinatedsupernatantderived from PHA-activated human peripheral blood
lym-phocytes.
In all three cases, the infection was renderednonproductive
in the treatedcells.Differentiationof infectedU937cellsinitiated
by
anyofthreeindependent
compoundsresulted inan
abrupt
decline in virusproduction.The interaction between HIV and the host cell U937 is
complex.
Undifferentiatedcells,
afteraninitialburst of virusproduction
in thefirst fewdays
afterinfection,subsequentlyshow restricted virus
replication.
Incontrast, theinfectionofdifferentiated cells is nonproductive despite high levels of
viral RNA accumulation. In both cases,provirus
accumula-tion isrelativelyconstant.Alterations in the pattern of c-myc
but not actin gene expression reveal that HIV infection
exerts aspecific and selective effect on cellular gene
expres-sion. These results demonstrate that the mechanisms
con-trollingHIVreplicationin monoblastoid cellsarespecificto
the cellular differentiation state. Inaddition, virus infection
itself resulted inphenotypicchangesin the monoblastoid cell
populationwhich suggest that HIVcanalter thecapacity of
these cellsto differentiate.
Others have noted that infected U937 cells manifest
al-teredphenotypes and that their differentiation is somewhat
inhibited (7, 21). Here we present molecular evidence in
supportof this concept and introduce the findingthat virus
production is diminished in normal and differentiated U937
cells; the operative mechanisms in these two cases are
functionally distinct.
Itis of interest to compare our results with the
observa-tions of Folks et al. (12, 13)onthe behavior ofa
persistently
infected cloned cell line (designated Ul) derived from the
parent line U937. The Ul line showed minimal constitutive
expression of HIV, and treatment of these cells with
phy-tohemagglutinin-activated lymphocytes supernatants
in-creased virus production (12). In ourhands,this treatment
inevitably causes a rapid decline in virus release, which is
consistent with the observed correlation between cellular
differentiationand reduced HIVproduction. The differences
between our observations on thepattern of HIVreplication
following acute infection of the U937 cell line and those of
Folks et al. (13) on the chronically infected Ul cell line
remain to beelucidated.
HIV replication in differentiated and undifferentiated
U937 cells is very similar to the pattern of visna virus
replication in normal sheep monocytes and macrophages. In
the visna virus studies, undifferentiated monocytes were
permissive and showed low levels of virus replication.
Differentiation oftheinfectedmonocytes wasinduced by an
interferon activity (16, 24, 29) and was accompanied by
increased viralgeneexpression (16, 29). Despite the elevated
levels ofcytoplasmic viral RNA, infectious virus was not
released from the differentiated cells (16). Thus, the same
relationshipbetweendifferentiationandviralreplication that was documentedhere for HIV in human monoblastoid cells
was observed previously for another lentivirus infection of
normal sheep monocytes and macrophages. Visna virus
infection ofsheep macrophages is associated with the
for-mation ofareservoir ofvirus (24) and the establishment of a
persistent infection(16,17).Thesetwo characteristics, virus
persistence and the establishment of a virus reservoir, are also important components in HIV infection. The state of
human monocyte differentiation and its impact on virus
replicationislikely to be central to the mechanisms
control-ling these two features of HIV infection and of crucial
importance in thepathogenesis of AIDS.
ACKNOWLEDGMENTS
C.D.P. wassupported by Public Health Service grants AI-21243 and AI-05875 and Cancer Training Grant CA 09254 from the National Institutes ofHealth (to Melvin Cohn) and by grant 000557 from the American Foundation for AIDS Research. D.D.R. was supported by the VeteransAdministration grant HL-32471 and by PublicHealthService contracts HB-67019 from the National Heart, Lung,andBloodInstitute andAI-52578from the National Institute ofAllergyand InfectiousDiseases.
on November 10, 2019 by guest
http://jvi.asm.org/
LITERATURECITED
1. Alizon, M., P. Sonigo, F. Barre-Sinousi, J.-C. Chermann, P. Tiollais, L. Montagnier, and S. Wain-Hobson. 1984. Molecular cloning oflymphadenopathy-associated virus. Nature (London) 312:757-760.
2. Armstrong, J. A., R. L. Dawkins, and R. Horne. 1985. Retroviral infection of the accessory cells and the immunological paradox in AIDS. Immunol. Today 6:121-122.
3. Armstrong, J. A., and R. Horne. 1984. Follicular dendritic cells and virus-like particles in AIDS-related lymphadenopathy.
Lan-ceti:370-372.
4. Bar-Shavit, Z., S. L. Teitelbaum, P. Reitsma, A. Hall, L. E. Pegg, J. Trial, and A. J. Kahn. 1983. Induction of monocytic differentiation and bone resorption by 1,25-dihydroxyvitamin
D3. Proc. Natl. Acad. Sci. USA 80:5907-5911.
5. Bar-Shavit, Z., S. L. Teitelbaum, G. P. Stricklin, A. Z. Eisen, A. J. Kahn, and H. G. Welgus. 1985. Differentiation of a human leukemia cell line and expression of collagenase inhibitor. Proc. Natl. Acad. Sci. USA 82:5380-5384.
6. Chayt, K., M. Harper, L. Marselle, E. Lewin, R. Rose, J. Oleske, L. Epstein, and R. Gallo. 1986. Detection ofHTLV-III RNA in lungs of patients with AIDS and pulmonary involvement. J. Am. Med. Assoc. 256:2356-2359.
7. Clapham, P., R. Weiss, A. Delgleish, M. Exley, D. Whitby, and N. Hogg. 1987. Human immunodeficiency virus infection of monocytic and T-lymphocytic cells: receptor modulation and differentiation induced by phorbol ester. Virology 158:44-51. 8. Cohn, Z. A., and B. Benson. 1965. The differentiation of
mononuclear phagocytes. Morphology, cytochemistry, and bio-chemistry. Int. J. Exp. Med. 121:153-169.
9. Estevez, M. E.,I.J. Ballart, R. A. Diez, N. Planes, C. Scaglione, and L. Sen. 1986. Early defect of phagocytic cell function in
subjects at risk for acquired immunodeficiency syndrome. Scand. J. Immunol. 24:215-221.
10. Favoloro, J., R. Treisman, and R. Kamen. 1980. Transcription maps of polyoma-virus specific RNA: analysis by two-dimen-sional nuclease S1 gel mapping. Methods Enzymol. 65:718-749.
11. Feinberg, A. P., and B. Vogelstein. 1983. A technique for radio labeling DNA restriction endonuclease fragments to high spe-cific activity. Analytic Biochem. 132:6-13.
12. Folks, T., J. Justement, A. Kinter, C. Dinarello, and A. Fauci. 1987. Cytokine-induced expression of HIV-1 in a chronically infected promonocyte cell line. Science 238:800-820.
13. Folks, T., J. Justement, A. Kinter, S. Schnittman, J. Orenstein, G.Poli, and A. Fauci. 1988. Characterization of a promonocyte clone chronically infected with HIV and inducible by 13-phorbol-12-myristate acetate. J. Immunol. 140:1117-1122. 14. Gabuzda, D., D. Ho, S. de la Monte, M. Hirsch, T. Rota, and R.
Sobel. 1986. Immunohistochemical identification of HTLV-III antigens in brains of patients with AIDS. Ann. Neurol. 20:289-291.
15. Gartner, S., P. Markovits, D. M. Markovitz, M. H. Kaplan, R. C. Gallo, and M. Popovic. 1986. The role of mononuclear phagocytes in HTLV-III/LAV infection. Science 233:215-219.
16. Gendelman, H. E., 0. Narayan, S. Kennedy-Stoskopf, P. G. E. Kennedy, Z. Ghotbi, J. E. Clements, J. Sta;dley,and G. Pezeshk-pour. 1986. Tropism of sheep lentiviruses for monocytes: sus-ceptibility to infection and virus gene expression increase during maturation of monocytes to macrophages. J. Virol. 58:67-74.
17. Gendelman, H. E., 0.Narayan, S. Molineaux, J. E. Clements, and Z. Ghotbi. 1985. Slow, persistent replication of lentiviruses: role of tissue macrophages and macrophage precursors in bone marrow. Proc. Natl. Acad. Sci. USA82:7086-7090.
18. Gordon, S., J. Todd, and Z. A. Cohn. 1974. In vitro synthesis and secretion oflysozymeby mononuclear phagocytes. J. Exp. Med. 139:1228-1248.
19. Grieco, M. H., M. H. Reddy, H. G. Kothari, M. Lange, E. Buimovici-Klein, and D. William. 1984. Elevated P2-micro-globulin and lysozyme levels in patients with acquired immune
deficiency syndrome. Clin. Immunol. Immunopathol. 32:174-184.
20. Haertle, T., C. Carrera, J. S. McDougal, L. Sowers, D. D. Richman, and D. A. Carson. 1988. Metabolism and anti-HIV activity of 2'-halo-2',3'-dideoxyadenosine derivatives. J. Biol. Chem. 263:5870-5875.
21. Hammer, S., J. Gillis, J. Groopman, and R. Rose. 1986. Invitro modification of human immunodeficiency virus infection by granulocyte-macrophage colony-stimulating factor and gamma interferon. Proc. Natl. Acad. Sci. USA 83:8734-8738.
22. Ho, D., R. Pomerantz, and J. Kaplan. 1987. Pathogenesis of infectionwithhuman immunodeficiencyvirus. N.Engl.J.Med. 317:278-286.
23. Ho, D. D., T. R. Rota, and M. S. Hirsch. 1986. Infection of human monocyte/macrophages by human T-lymphotropic virus typeIII. J. Clin. Invest. 77:1712-1715.
24. Kennedy, P. G. E., 0. Narayan, Z. Ghotbi, J. Hopkins, H. E. Gendelman, and J. E. Clements. 1985. Persistent expressionof Ia antigen and viral genome in visna-maedi virus-induced in-flammatorycells. J. Exp. Med. 162:1970-1982.
25. Klatzmann, D., and J. C. Gluckman. 1986. HIVinfection: facts and hypotheses. Immunol. Today 7:291-296.
26. Koenig, S., H. E. Gendelman, J. M. Orenstein, M. C. dal Canto, G. H. Pezeshkpour, M. Yungbluth, F. Janotta, and A. Aksamit. 1986. Detection of AIDS virus in macrophages in brain tissue from AIDS patients with encephalopathy. Science 233:1089-1093.
27. Luciw, P. A., S. J. Potter, K. Steimer, D. Dina, and J. A. Levy. 1984. Molecular cloning of the AIDS-associated retrovirus. Nature(London) 312:760-763.
28. Mitchell, R. L., L. Zokas, R. D. Schreiber, and I. M. Verma. 1985. Rapid induction ofthe expression ofproto-oncogenefos during human monocytic differentiation. Cell 40:209-217. 29. Narayan, O., D. Sheffer, J. E. Clements, and G. Tennekoon.
1985.Restricted replication oflentiviruses: Visna virusesinduce aunique interferon during interaction betweenlymphocytes and macrophages. J. Exp. Med. 162:1954-1969.
30. Nicholson, J. K. A., G. D. Cross, C. S. Callaway, and J. S. McDougal. 1986. In vitro infection of human monocytes with human T lymphotropic virus type III/lymphadenopathy-associ-ated virus (HTLV-III/LAV). J. Immunol. 137:323-329. 31. Pallesen, G., J. Gerstoft, and L. Mathiesen. 1987. Stages of
LAV/HTLV-III lymphadenitis.I. Histological and immunolog-ical classification. Scand. J. Immunol. 25:83-91.
32. Pauza, C. D. 1987. Regulation of human T-lymphocyte gene expressionby interleukin-2: immediate-response genes include the proto-oncogene c-myc. Mol. Cell. Biol. 7:342-348. 33. Pauza, C. D. 1988. HIV persistence in monocytes leads to
pathogenesis andAIDS. Cell. Immunol. 112:414-424.
34. Plata, F., B. Autran, L. Martins, S.Wain-Hobson, M.Raphael, C. Mayaud, M. Denis, and J. Guillon. 1987.AIDSvirus-specific cytotoxic T lymphocytes in lung disorders. Nature (London) 328:348-351.
35. Poli, G., B. Bottazzi, R. Acero, L. Bersane, V. Rossi, M.Introna, A. Lazzarin, and A. Mantovani. 1985. Monocyte function in intravenous drug abusers withlymphadenopathy syndrome and in patients with acquiredimmunodeficiency syndrome: selective impairment of chemotaxis. Clin. Exp. Immunol. 62:136-147. 36. Popovic, M., M. G. Sarngadharan, E. Read, and R. C. Gallo.
1984. Detection, isolation, and continuous production of cyto-pathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 224:497-500.
37. Rabbitts, T. H., P. H. Hamlyn, and R. Baer. 1983. Altered nucleotide sequences of atranslocated c-mycgene in Burkitt's lymphoma. Nature (London)306:760-765.
38. Ralph, P. 1981. Continuous macrophage cell lines-their use in the study of induced and constitutive macrophage properties and cytotoxicity, p. 175-195. In M. A. Landy. Lymphokines, vol. 4. Academic Press, Inc., Orlando, Fla.
39. Richman, D., R. Kornbluth, and D. Carson. 1987. Failure of dideoxynucleosides to inhibit human immunodeficiency virus replication in cultured humanmacrophages. J. Exp. Med. 166: 1144-1149.
on November 10, 2019 by guest
http://jvi.asm.org/
40. Smith, P., K. Ohura, H. Masur, H. Lane,A.Fauci, andS.Wahl. 1984. Monocyte function in the acquired immune deficiency syndrome: defective chemotaxis. J. Clin. Invest. 74:2121-2128.
41. Sundstrom, C., and K. Nilsson. 1976. Establishmentand char-acterization ofahumanhistiocytic lymphoma cell line (U937).
Int. J. Cancer17:565-577.
42. Weiss, A., H. Hollander, and J.Stobo. 1985. Acquired immuno-deficiency syndrome: epidemiology,virology andimmunology. Annu. Rev. Med.36:545-562.
43. Wiley, C., R. Schrier, J. Nelson, P. Lampert, and M.Oldstone. 1986. Cellular localization of human immunodeficiency virus infectionwithin the brains ofacquiredimmunedeficiency
syn-dromepatients. Proc. Natl. Acad. Sci. USA83:7089-7093.