Copyright© 1990, American Society forMicrobiology
Proviral Insertions
within the int-2 Gene Can Generate
Multiple
Anomalous
Transcripts but
Leave the
Protein-Coding
Domain
Intact
CLIVE
DICKSON,'*
ROSALINDSMITH,' SHARON BROOKES,2AND GORDON PETERS2Imperial Cancer Research Fund Laboratories, Lincoln'sInnFields, London WC2A
3PX,l
andImperialCancerResearch FundLaboratories, St. Bartholomews Hospital, DominionHouse, LondonECJA 7BE,2 UnitedKingdomReceived 10 August 1989/Accepted 13 October 1989
We examined the effects ofmouse mammary tumor virus integration on the multiple RNA transcripts expressedfrom the int-2 proto-oncogene invirally inducedbreast tumors. Proviral insertion eitherupstream ordownstream of thegenecouldsimultaneously activatetranscriptionfrom threedissimilar int-2 promoters.
In some tumors,theactivating provirus lies within thetranscription unitand disruptsthe structures ofthe variousRNAs. Insertions in the 5' region of thegene hadcomplexeffects depending onthe orientationand position of the provirus relative to the three promoters and intron-exon boundaries. RNase protection experimentsidentifiedtranscriptsinitiated in the virallongterminalrepeat,at normal andcrypticsites inthe
int-2sequences,and fromcrypticpromoters inaninvertedprovirus.At the3' end, insertions occurredwithin
the untranslated trailer and provided alternative terminationsignals that substituted for one orbothofthe normal thepoly(A) additionsites.However,innoinstance,of the20tumorsanalyzedindetail,didaprovirus
perturbthepresumedopenreadingframe of thegene.These datastrongly implicatethe normalproductofthe
int-2gene, whichisrelated to the fibroblast growthfactorfamily, as acontributoryfactor invirallyinduced mammary tumors.
The int-2 proto-oncogene is now known to encode a
memberofthe fibroblast growth factor family ofproteins,
and its expression, at least in the mouse, appears to be
restricted to specificembryonic cell typesat variousstages
ofdevelopment (2, 7, 26, 27). However, the initial
identifi-cation and structural characterization of the gene resulted
from its transcriptional activation in tumors induced by
mouse mammary tumor virus (MMTV) (3, 11, 16, 17). For
example, in the BR6 strain of mice, around 70% of the
spontaneously arising mammary tumors areclonal
prolifer-ationsof cells that havesustained a proviral insertion within
about 15 kilobase pairs (kb) on either side of the int-2 gene
(16, 18, 19).
Typically,these tumors express multiple int-2 transcripts
that are superficially analogous to RNAs detected in the
embryoand in culturedembryonal carcinoma (EC) cell lines,
suchasF9 andPCC4 (10, 24, 27). In EC lines, the expression of int-2increases when the cellsdifferentiate in response to
retinoic acid and dibutyryl cyclic AMP, and different cell
lines contain different patterns of transcripts. They have
therefore provided a convenient system in which to
charac-terizethevarious species of RNA (24). By a combination of
RNaseprotection analyses and blot hybridization with spe-cificprobes, six different classes of int-2 mRNA have been
distinguished that initiate atmultiple cap sites within three
separate promoter domains and terminate at either of two
polyadenylationsites (10, 24).
The reasons behind this complexity are unclear, since all transcripts are predicted to encode the same protein (11, 24).
One possibility would be to facilitate control of
tissue-specific expression, or there could be subtle differences in
the stability or translational efficiency of the various
tran-scripts. It was therefore essential to establish whether the
RNAs seeninMMTV-induced tumors are indeed analogous
to their embryonic counterparts or whether they represent
tumor-specific variants. Moreover, a subset of mammary
*Corresponding author.
tumors expressanomalous int-2 RNAs as aconsequenceof proviral integration within the transcription unit (19). Here wehave examined thestructuresof these int-2transcriptsin
a series oftumors and established that proviral insertions
caninfluence all three promoters and alter the structures of
transcriptsat either the 5' or3' end, but innocase do they
disrupt the protein-codingcapacity ofthe gene.
MATERIALSANDMETHODS
Analysis of tumorDNAand RNA.Theprocedures used for theextraction ofhigh-molecular-weightDNA andtotal RNA
fromMMTV-inducedmammary tumorshave beendescribed
previously (3, 17), and the examples analyzed here were
included in theseries of BR6 mammary tumors described in
an earlier report (19). Poly(A)+ RNA was enriched by
chromatography onpoly(rU)-Sepharose (3). Samples(5 ,ug)
werefractionated by electrophoresis in agarose gels
contain-ing formaldehyde (9) and transferred onto nylon filters by
blottingin 20x SSC (1x SSCis 0.15 M NaClplus0.015 M
sodium citrate). Hybridizations were performed with DNA
probes thatwere labeled with 32P by either nick translation
orprimingwith randomoligonucleotides.Theconditions for
hybridizingandwashingoffiltersandautoradiography were
asdescribed previously (3). In some experiments, 20 ,ug of
F9 orPCC4cell RNAs wasincluded as a control, and RNAs
weredenatured inglyoxal rather than formaldehyde (24). Exon-specific probes. Inaddition to the int-2 genomic DNA probe, designated int-2 f (11), which is specific for exon III of the gene, three furtherprobes were prepared that contained sequences present exclusively in exons la, I, and lb (24). These corresponded to nucleotides (nt) 590 to 1219, 1342 to 1681, and 1681 to 1815, respectively, in the genomic se-quence(11).
Recombinant DNA techniques. DNA from tumors W26, E115, and D157B was digested to completion with EcoRI
and ligated into the separated arms of XgtWES.XB, or X
L47.1,asdescribedelsewhere (3, 17). After in vitro
packag-ing, approximately 5 x 105 bacteriophage were plated on
784
on November 10, 2019 by guest
http://jvi.asm.org/
ANOMALOUS int-2 RNAs AFTER PROVIRAL INSERTION
Qm U) CU
W~ 0
(0 co
V-llJ
a
la
1
lb
Ill
.t. -2 .9
U-2.7
*
I!
q " -1.8
-1.6
Is:9
,IjiI
-2.9
-2.7
-1.6
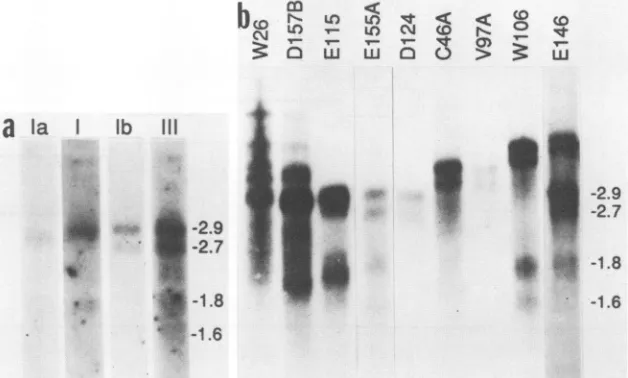
FIG. 1. int-2 RNAs in mammary tumors. Samples (5 ,ug) of poly(A)+ tumor RNA were fractionated in denaturing agarose gels, transferred tonylonmembranes, and hybridized with32P-labeledprobes specific for mouse int-2 sequences. (a) RNA from tumorE155Awas hybridized sequentiallywith probesspecific for exonsIa,I, Ib,andIII as indicated (see Materials and Methods). (b) RNAs from the indicated mammary tumors werehybridized with the int-2f probe specific for exon III. The positions of the major classes of int-2 RNA and their sizes in kilobase pairsareindicatedon the right ofeach panel.
LE392 cells and screened with both MMTV long terminal repeat(LTR) and int-2 probes. Phage containing inserts that werepositive for both were plaque purified, and the
respec-tiveEcoRIfragments were transferred into plasmid vectors
for further analysis.
DNAsequence analysis. After the clonedEcoRIfragments
werecharacterized by restriction enzyme mapping, selected
fragments that spanned the virus-host junctions were
sub-cloned into M13mp vectors. DNA sequences were
deter-mined inboth orientations by the dideoxy-chain termination
procedure. For W26, the sequence of the entire LTR was determined toverifythat noadditional viral information was present. A 6-base-pair (bp) duplication was noted at each endofthe LTR,indicatingthat thesite of integration was at
nucleotide 1089 in thegenomic sequence (11). For E115 and
D157B, only the 5'junctions wererecovered and
character-ized, but these established both the position and the
orien-tation of the respective proviruses (indicated in Fig. 2).
RNase protection. Samples (generally 2
pRg)
ofpoly(A)+ tumor RNA or 20 ,ug of total RNA from EC cells weresubjectedtoRNaseprotection analysisexactlyasdescribed
by Smith etal. (24). Theuniformly labeled antisense RNA
probes were prepared by transcribing cloned fragments of
genomicDNAwithphageSP6 RNApolymerase.Theprobes
are numbered as in the previous report (24) (with the
addition of probes 1.1, 1.2, and 1.3), and the restriction
enzymesitesthat mark the 5' and 3' endsofthe
int-2-specific
sequencesin eachprobeareindicatedin the relevantfigures.
The sizes of eachproberefertotheint-2-specific sequences
only (occasionally, enzyme sites in the
polylinker
of thetranscribed
plasmid
weremoreconvenient)
andwerecalcu-lated withoutconsideringexactcleavagepositionswithinthe
recognition sites. However, when measuring the sizes of
protected fragments, the exact ends were taken into
ac-count. Even so, we estimate that
fragment
sizes deducedexperimentallyare only accurateto +5bp.
RESULTS
Typicalandatypicalint-2 RNAs in MMTV-inducedtumors. Most tumors in which the int-2 gene is
transcriptionally
activated carry an MMTV provirus in adjacent cellular
DNA, rather than within the gene, in an orientation
appro-priate for enhancement of the normal int-2 promoters by
elements in the viral LTR (3, 18,19). These tumors typically
contain four sizes ofint-2 RNA, ofapproximately 2.9, 2.7,
1.8, and 1.6 kb, all of which are detectable with a probe for exonIIIofthe gene(Fig. 1). In the present study, the tumor
designated E155A was used to represent the typical case,
sincetheactivating MMTVprovirus is located about 10 kb
upstream ofint-2 (19). Results obtained with other typical
tumors were qualitatively indistinguishable. However, by analogy with ECcells,thefour bands detected on RNA blots were assumed to reflect six distinct types of RNA (10, 24). Thus, the2.9-kbbandisnowunderstood tocompriseRNAs that initiate at either the P1 or the P2 promoter. Tumor
RNAs ofthis sizecontained sequences derived from either
exon Ia(initiatedatP1)orexonI (initiatedatP2),while the
2.7-kbtranscriptwasnotdetectedby probes fortheseexons
(Fig. la). This latter RNA presumably initiates at a third
promoter,designatedP3(Fig.2),since itwasdetectablewith
aprobespecific forexon
Tb
(Fig. la) (24).Arationalizationof thesefindingsispresentedinFig. 2. Thesmaller 1.8- and
1.6-kb forms of int-2 RNA that are barely visible in the
reproductionofFig. la have been showntobeanalogsof the
2.9- and 2.7-kb classes, respectively, that terminate at the upstream poly(A) addition site (24). These smaller
tran-scripts are consistently underrepresented in tumors
com-paredwith EC cells.
Althoughthemajority of int-2-positivetumorsconformto
this typical pattern, our
previous
survey of spontaneoustumorsin BR6mice revealed several
examples
in whichthepatterns
ofint-2transcriptswereunusual(19).Some of theseatypicalcases are shown in
Fig.
lb. Fromrelatively
detailedmapping of the integrated
proviruses
in these tumors, itbecameclear that the anomalous
transcripts
arosethrough
physical disruption of the int-2
transcription
unit and thatthiscouldoccur ateither the 5'or3'end.
Thus,
tumorsW26,
E115, and D157B
expressed
unusual RNAs as a result ofproviral insertion in the5'
region
of the gene, while severalothers, suchasD124,
C46, V97A,
andW106,
hadproviruses
VOL.64, 1990 785
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.152.466.78.267.2]W26
D1
57B1089 1195
E115
14
1503
P1
i1
::=la
lizi:il
1175 1671
P2
P3
B Pv S Sm St N Pv
0 2500
1225
1681
-
Pvull-Sacl
Nael-BamHI
184
-~
Stul-Smal991
Stui-Pvull
9
11
12
13
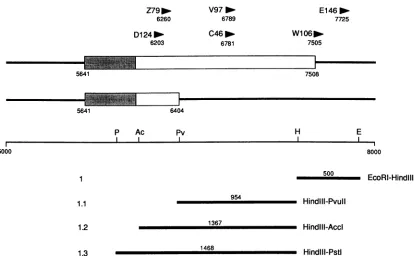
FIG. 2. Proviralinsertionsin thepromoterregion ofint-2. Intheupper part of thefigure,thestructuresofthreeclassesofint-2 RNAare
drawn to scale(relativetotherestrictionmapbelow),withexonsIa,I,andIbrepresentedasboxes and numbersreferringtocoordinates in thegenomicsequenceofMoore et al.(11).Shadedareascorrespondtoprotein-codingsequences,andthemultiplestartsites associated with promotersP1 and P2 aredepictedbythevertical lines. Thepositions andorientationsofintegrated provirusesin thenumbered mammary tumors are shown as arrowheads, with their positions in thegenomic sequenceindicated below. Therestriction map showsthe sites of cleavage for theenzymesBamHI (B), NaeI (N), PvuII (P),Sacl(S),SmaI(Sm),andStul(St)thatarerelevanttothegenerationofspecific antisense RNA probes. Not all sitesare included. The lines below themap define theextents and positions of the genomic sequences transcribed intoRNA probes, numberedas inreference 24. Thelength ofeachprobe and therestriction sitesthatdefine eachendareas
indicated.
within the presumed 3' untranslated region, leading to
ex-tendedforms of int-2 RNA (Fig. lb).
Locating proviruses by Southern blotting oftumor DNA
wasnotsufficientlyaccurate to drawfirmconclusionsabout
their effects on the structure and coding capacity of int-2 RNAs. Two approaches were therefore adopted to map these proviruses more precisely. First, recombinant DNA clones were isolated representing the junctions between
MMTVproviral DNA and int-2 genomic sequences. Inthe
second approach, tumor RNAs were subjected to RNase
protection analysis with a series of antisense probes derived
from specific regions of the genomic DNA. Since this
technique detects discontinuity between the RNA and
ge-nomicsequences, the mapping of hybrid RNAs generated by
viralinsertion canbe used to locate the site ofintegration.
Characterization ofvirus-hostjunctions. Bacteriophage K
libraries were constructed containing EcoRI-digested DNA
from tumors W26, E115, and D157B and screened with both int-2- and MMTV-specificprobes. Tumor W26 contained a
singleMMTV LTRin a 5' to 3' orientation relative to int-2,
resulting in an 11.3-kb EcoRI genomic DNA fragment as
opposed to the 10-kb normal allele (11). With E115 and
D157,only the respective5'junctionfragments were
recov-ered.Whenthesewerecharacterizedand the DNA sequence
was determined across the virus-cell boundaries, it was evident that the provirus in D157B was also in the same transcriptional orientationasthe int-2gene, but apparently
intact. In tumor E115, on the other hand, a
complete
proviruswas present inan invertedorientation. These data
also located the precise sites ofintegration in the
genomic
sequencedescribedbyMooreetal.(11)at nt1089, 1195,and
1503, respectively, in tumors W26, D157B, and E115
(Fig.
2). No common features were evident in the sequences
immediately adjacenttotheintegration sites.
The three proviruses in question lie within the promoter domains mapped in EC cells in previous studies (Fig.
2).
Thus, the LTR in tumor W26 had integrated in exon
la,
betweenP1 andP2,theprovirus in D157B lay just upstream of P2, while that in E115 split P2 and P3. We therefore attempted to reconcile these findings with the RNA struc-turespresent in therespectivetumorsby RNaseprotection.
Tumor RNAs containing exon Ia. int-2 transcripts that initiate atpromoter P1 and contain an exon Ta to Tb splice
have beenidentified in EC cells by both RNase mapping and
cDNAcloning (10, 24).However, since these transcriptsare
atrelativelylowabundance in F9 cells and are undetectable
in PCC4 cells, itwas important to establish whether
coun-terparts existed in typical and atypical mammary tumors. I
--
gfb---n .s.?.fI...
NV,
"....V.M1.1.1677 1993
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.125.494.71.388.2]ANOMALOUS int-2 RNAs AFTER PROVIRAL INSERTION
a
1840
10CrU)(
Cm LLUJ
3
I
M
*
-218
-p -1 76
138',
'S1
b
MLL
LO L()
w-
517-
398-
296-
221-I
4
(P1)
-41
Um
154-
145-91'>
_
-75
FIG. 3. RNaseprotection analysisofexonIa. Approximately2 ,ug ofpoly(A)+ tumor RNA from the indicated tumors or 20 p.goftotal RNAfrom eitherF9 orPCC4cells that had been treated withretinoic acid anddibutyrylcyclic AMP wassubjectedtoRNase protection analysisasdescribed previously(24). (a) Results obtained with the probedesignated 12 in Fig. 2. Numbersto theleft of the panel identify thepositionsandsizes(nucleotides)of theRNase-resistant products.Thesizes wereestimated relativeto aseries ofplasmidDNAfragments of known length (HinfI-digested pAT153)and astandard DNAsequence ladder, indicated as lane M. Numbers on the rightidentify the positions andsizesofthestandardmarkers. (b) Probe 13 was used todelineatethe multiple start sitesassociatedwithinitiationatpromoter
P1.The arrowheadsontherightidentifyproductscorrespondingtothe major cap sitesobserved in F9 cells. Numbers on the left identify the sizes(in nucleotides) and positions ofDNAmarkerscalibrated witha sequenceladder.
Theexpression of exon
Ta
wasfirstexamined by protectionof the 184-nucleotide antisense RNA transcribed from the
genomic DNA segment designated as probe 12 (Fig. 2).
Assumingthat the structureofthetranscript wasasdepicted
in Fig. 2(subsequently confirmedwithmultiple probe
com-binations), probe 12 should have detected discontinuity at
thesplice donor siteatthe 3' end ofexon Ta.TumorE155A,
in which the provirus lies distant from the int-2 gene,
contained RNAs that include exon la and splice at the
predicted position, yielding a protected fragment of 138 nt
(Fig. 3a). The 184-nt fragment corresponded to full
protec-tion of theprobeandmay havebeenindicative ofunspliced
forms in thepreparationof total RNA used in thesestudies;
thecontrol RNA fromrat208Fcells and F9 cell RNA didnot
yield thisfullyprotectedproduct, indicating thatconditions
were adequatefor completedigestionof the probe.
A different result was obtained with RNA from tumor
W26, in which the solo LTR lies within exon
Ta.
Twoprotected fragmentswereobservedthatwereeachabout50
ntsmaller than thoseseenin thetypicaltumorsample.Other data
(Fig.
4) confirmthatthelargerof thetwo(-135 nt)wasderivedfrom unspliced RNAand represented the distance
between the LTRandtheend oftheprobe
(calculated
tobe137 nucleotides), while the -90-nt fragment reflected the
distance between theLTRand the end ofexon
Ta
(calculated
as 91 nucleotides). It is not clear why this latterfragment
resolves intomultiple species, 1base apart,but the effect is
consistently seen with probes that span this
discontinuity
(Fig. 4aand other datanot shown). TumorE115, in which
theproviruslies downstreamof the
splice
donorsite,
didnotshow evidenceoftranscriptsthatcontainexon Ia
(data
not shown).787 VOL.64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.123.486.73.471.2]a
( U(P2)j
321
0'
(Pi)
U')
I
M
--1 244
co
CJQ-WW
C\C L
ll LL
4..
wt
*
-520
'
-396
a
-354
496
'o
(P2),J
'
-218
I.. ..
a"
a
-176
(SA)
147
(P3)
141 o188
Po(SD)
91
0 I4
-75
FIG. 4. RNase protection analysis ofexons I andIb. (a) RNase protection was performed on theindicated RNA samples with the
1,225-nucleotide PvuII-SacI probe designated9in Fig. 2. Numbers onthe left of the figure identify the positions andsizes ofrelevant RNase-resistant fragments, calculated relative to the standard markers and sequence ladder on the right (lane M). Discontinuities
characteristic of the multiplestartsin P2 and P1 (24)areindicated by the lines, and other discontinuitiesareidentifiedthatcorrespondtothe
splice donor inexonIa(SD), the spliceacceptorinexonlb (SA), and initiationatP3 (10).The 321-ntfragmentmapsthepositionofprovirus
integration intumorE115. (b) A similar analysiswasperformedwith probe11.Since this probe extends fromthe NaeIsiteatnucleotide1681,
shorteranalogsareproduced for each ofthediscontinuitiesshown in panela.Thepositions of the multiplestartsites associatedwith theP2 promoterareindicated by arrowheads, while the 496- and 188-nt fragments correspondtothesites of proviral integration intumorsD157B andE115,respectively. The arrowhead adjacenttothe -500-nt fragment in the W26 laneidentifies the novelP2initiationsitecharacteristic ofthistumor,while the larger bandinthistrackderives from unspliced RNA initiated in theLTR. Thesizesof fragments (innucleotides)
werededuced directly fromthesequence ladder (laneM).
788
LON
L-a
3S
M
S
'.
fj,.
'.
S
oill
ow
.0
^a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.75.560.51.620.2]ANOMALOUS int-2 RNAs AFTER PROVIRAL INSERTION
Although these results implied splicing from the 3' end of exon Ia, in at least a proportion of the tumor RNAs, it was
of interest to examine the 5' end of such transcripts, to
ascertain whether they initiated at the same cap sites as in F9 cells. The analyses were therefore repeated with probe 13, which extends upstream of promoter P1. InE15SA, aseries of protected fragments were observed that were indicative of multiple start sites withinP1, each fragment being equivalent to acomplete exon Ta(Fig. 3b). Most of these 5' ends were analogous to those seen in F9 cells, but there were also additional discontinuities reflected in fragments of between 280 and 450 nt. Itis not clear whether these represent novel, tumor-specific initiation sites or simply an increase in the efficiencies with which normal cap sites are employed. Neither the W26 nor E11S tumor RNAs gave signals for initiation at P1 (data not shown).
Tumor RNAs containing exonlb. Previous studies on EC cells established that int-2 transcripts that include exon Ia
are spliced such that the donor site (nucleotide 1175) is linked to the consensus acceptor site (nucleotide 1661) that marks the 5' end of exon lb (10, 24) (Fig. 2). However, we now recognize that other transcripts initiate at a unique site very close to the 5' end of exon Ib, at what was originally
interpreted as an alternative splice acceptor (nucleotide
1667). Although separated by only 6 bases, the two
discon-tinuities, attributabletoeitherspliced RNA initiated at P1 or
de novo initiation at promoter P3, can be readily
distin-guished. For example, when probe 9 (Fig. 2) was used in
RNaseprotection experimentswith tumorE1S5ARNA, two protected fragments of 147 and 141 nt were observed at
approximately equivalent intensities (Fig. 4a). This would
imply that transcription initiation at promoters P1 and P3
occurredwith roughlyequalefficiency in typical mammary
tumors, in some contrastto the situation in F9 cells where P3, reflected in the 141-nt product, was clearly dominant (24). It also contrasts with the result obtained from tumor W26 in which the converse was true (Fig. 4a). Here, the
majorRNA species initiate in the MMTV LTR, and about
70% ofthesetranscripts undergoaspliceattheendofexon
Ta(91-ntfragment in Fig. 3a). The recipient ofthis splice is
presumablythe 5' end ofexon
Tb,
asreflected in the 147-ntprotected fragment, providing an explanation for the
strengthof thissignal relative tothefragmentgenerated by
initiation atP3 (Fig. 4a).
With tumor
E115,
the signal indicative of thesplice
acceptor sitewasextremely faint(Fig. 4a). Since there was
noevidence forRNAsplicing from exonIain this tumor, it
is possible that minor transcripts initiating at a
cryptic
promoter(s) within the MMTV provirus become
spliced
tothis site. However, the signal
corresponding
to initiationatP3 was also unexpectedly weak,
despite
the fact that theMMTV
provirus
inEllSispresumably
inanidealposition
to enhancetranscription
from thispromoter.Either the leveloftranscription from P3 is inherently low in this tumor, for
reasons that are not clear, or the signals derived from P3
were masked by more abundant
transcripts
initiatedelse-where. The most likely
origin
of these RNAs would be initiation from cryptic promoters on thenegative
strand of the MMTVprovirus,but this hasnotbeenformally
proved.
Tumor RNAs containing exon I. Themajor protected
fragmentgenerated fromEllSRNAwas
approximately
321ntand corresponded to
discontinuity
atthe siteofproviral
integration.Thismappedabout 100nucleotides downstream
ofthe P2
domain,
butproviral
insertionappeared
toprevent initiation at this promoter. However, in thetypical
tumor,E1SSA, initiation within P2 could also be visualized with
probe
9, as evidencedby
aheterogeneous
set ofbands ofbetween 450 and 600nt. These are very similar in
distribu-tion to the analogous start sites observed at much lower
levels in PCC4 cells (Fig. 4a). The dataare complicated by
the fact that
probe
9 can also detect heterogeneous starts within P1(inactive
in PCC4 cells), as representedby
the characteristic series ofprotected
fragments
between 215and 265nt. These latterfragments
werenotobservedwith RNA from W26 andE115, confirming
thatP1doesnotfunction in thesetumors(Fig. 4a). However,
withW26RNA,
therewasevidence for novel initiation sitesin the upstream
region
ofP2
(fragments
of-650to750nt)
andaprotected
fragment
ofaround 91 nt was
again
observed(Fig. 4a),
representing
the distance between the MMTV LTR and thesplice
donor site in exonTa,
as inFig.
3a.Theseconclusionswere
supported
and clearerindicationsfor initiation in P2wereobtained
by
using probe
11(Fig.
4b).
This
probe
extendedfrom the NaeI siteat1681(Fig. 2)
andtherefore
yielded
shorteranalogs
of each of thefragments
generated
withprobe
9.Only
thoseoverabout 150ntin size are shown in thegel depicted
inFig.
4b.Transcription
initiation at P2 was reflected in
protected
fragments
ofbetween 308 and 443 nt, with the
major
start site(361-nt
product)
centered on nucleotide 1320.Thus,
typical
mam-mary tumors such as E1SSA expressRNAs that arealmost exact counterparts ofthose initiatedat P2inPCC4 cells. In
W26,
onthe otherhand,
a different set ofpresumptive
cap siteswereagain
evident withprobe 11,
themostprominent
of which
(-500-nt
fragment)
mapped
about 150 nucleotidesupstream of the
major
startnormally
associated with P2.Since a
fragment
of similar size was also observed withE1SSA
RNA,
albeitatmuch lowerintensity (Fig.
4b),
it maycorrespond
toanormally
rarecapsitewhose usage isgreatly
enhanced
by
thenearby
insertionofthe MMTV LTR. Thelarger product
from W26 RNA would be consistent withunspliced
RNA sinceit represents thedistancebetween the LTR(1089)
and the NaeI site(1681)
that marks the end ofprobe
11.Probe 11 also revealed discontinuities that confirmed the sitesof
proviral
integration
intumorsEllS(188-nt
fragment)
and D157B
(496-nt
fragment)
relative to the NaeI site. ForD157B,
there was insufficient tumor RNA toperform
acomprehensive analysis,
but the results shown inFig.
4bsuggested
that themajor
transcripts
in this tumor wereinitiated in the viral
LTR,
upstream ofandsubstituting
for P2.Tumor RNAs with abnormal 3' ends.
Although
proviral
insertionstendtobemore common5'tothe int-2 gene, there
are nevertheless several tumors in which
proviruses
have beenmapped
at the 3' end of the gene. In S of the 20int-2-positive
BR6 tumors, theprovirus
was inserted withinthe
transcription
unit,
in the sametranscriptional
orienta-tion.
Figure
lbshowed that thesetumors containedatypical
int-2
RNAs,
presumably
owing
toperturbation
of the normal terminationsignals
fortranscription. Indeed,
some ofthese RNAswere shownby
sandwichblotting
to behybrid
tran-scripts
comprising
both int-2 and MMTV LTR sequences (22; data notshown),
suggesting
thatthey
terminate in theviralLTR.
However,
since the int-2 gene hastwoalternativepolyadenylation
signals,
it was of interestto determine theprecise
location ofthevariousintegrated
proviruses
andto assess whether any of the insertions affected theprotein-coding
domain.Rather than construct recombinant
phage
librariesfor all thesetumors,welocated theproviruses by
RNasemapping.
Although
this method is lessprecise
thancloning
andse-VOL.64, 1990 789
on November 10, 2019 by guest
http://jvi.asm.org/
Z79 Fm 6260 D124
6203
5641
5641
V97P
6789
C46D.
6781
El46
-7725 W106 .
7505
7508
6404
H
p Ac Pv
5000
954
1367
E
8000
500
EcoRl-Hindill
HindlIl-Pvull Hindlil-Acci
1.3 1468 Hindill-Psti
FIG. 5. Proviralinsertions in the 3' untranslated regionofint-2. The3' end of the int-2geneis drawntoscale relativetotherestriction
map,withexonIIIdepictedasboxesinwhichprotein-codingsequencesareshaded. Theexonterminates at twoalternativepolyadenylation signalsatnucleotides7508 and 6404 in the genomicsequence(11).Arrowheadsatthetop of thefigure identifythepositionsandorientations
ofproviruses in the indicatedtumors.The restrictionmapshows thecleavage sites forthe enzymesAccI(Ac),EcoRI(E), Hindlll(H),PstI
(P), and PvuII (Pv) thatwereused ingenerating thenumbered antisenseprobesshown beneaththe map.Onlyrelevant sitesareshown.
quencing of virus-host junctions, we estimated that the
positions deduced from the sizes of protected fragments wereaccurate to withinabout 5 bases.The results of these
analyses aresummarized inFig. 5, anddatasupportingthe conclusions are presented in Fig. 6. Briefly, a series of antisense probes wereprepared, designated 1, 1.1, 1.2,and 1.3(24)(Fig. 5), thatwerecapable ofdetecting discontinuity between tumor-specific RNA and the genomic sequences
encompassing exon III. Wherever possible, each
disconti-nuity and therefore site of integration was mapped with at leasttwospecific probes. For example,theprovirusintumor D124waslocatedatnucleotide 6203 (+5) by using probes1.2 and1.3. Withtheformer, extendingto the AccI site
(nucle-otide 5997), a protected fragment of 206 nt was obtained (datanotshown), whilethelatter, extendingtothePstIsite
(5896), yielded a 302-nt fragment (Fig. 6a), suggesting that integration had occurredatthese distances from the respec-tive restriction sites. By similar reasoning, the provirus in tumor Z79was mapped to nucleotide 6260, approximately
359 ntfromthe PstI site(Fig. 6a). Representative data for
theother tumors and therespective probes used are
illus-trated inFig. 6b andc.
DISCUSSION
Multiple int-2 promoters respond to proviral activation.
Previousstudiesonthe expression of theint-2geneinmouse
embryos and EC cells have indicated aninherent transcrip-tionalcomplexity for which there isasyetnoclear explana-tion (7, 10, 24, 26, 27). Much of the complexity arises through the differential usage of three distinct promoter
regions in which RNA transcripts initiate at multiple sites (10, 24). However, all the transcripts characterizedthus far would be predicted to encode the same 27-kilodalton
pro-tein, now recognized as a member of the fibroblast growth
factor family (2, 4, 11). One possibility is that differential promoterusagereflectsaneedtorespondtofactors govern-inggeneexpression invery specific celltypes andathighly
restricted times duringdevelopment.
InMMTV-induced mammarytumors, expression ofint-2 is activated inacelltypeinwhichthegeneisnormallysilent, atleastasjudged by steady-statelevels of RNA(3,7).Itwas
therefore important to establish whether the transcription
patterns intumors reflected those seenin EC cells, suchas
F9 and PCC4, and whether the different int-2 promoters
responded differently to activation by MMTV. The three
promoter domains that have been distinguished, designated P1, P2, and P3 (10, 24) (Fig. 2), havenoobvious features in
common;theP1 region hasanaveragebasecomposition, P2 isveryG+C-rich, while the regionnowassociated with P3is unusually C+T-rich, analogous to some ribosomal gene
promoters(5). Northern (RNA) blotting analyses with DNA probes specific for exons Ia, I, and Ib indicated that in a
typicaltumor,suchasE155A, transcriptsaredetectable that correspondtoinitiationateachof the threepromoters(Fig. la). Thiswas subsequently confirmed by RNase protection withaseriesofantisenseprobes, aspresentedinFig. 3 and 4. Qualitatively, the RNAs initiated at each promoter are
very similar to those seen in EC cells, although there are minor variations in the distribution ofcap sites.
Quantita-tively, eachpromoter is moreactive in tumors thanin EC cells, and althoughP2 and P3aredominant,it is clear thatall three promoters function in mammary tumors. This con-trastswith the situation in F9 andPCC4cells,inwhichonly twoof thethreepromotersfunctionineach celltype:P1and P3in F9 cells, and P2 and P3 in PCC4 cells (10, 24). Since the
sequences involved in directing initiation from each
pro-1.1 1.2
OME'S'N
m
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.106.520.68.324.2]ANOMALOUS int-2 RNAs AFTER PROVIRAL INSERTION
a
PROBE1.3Z79
-Dl124'o
~XX'359
sxwow'302
cxxxx30
04 0)
o N
b
PROBE 1.1V971
C46
r
1468 _ 954isssssss 376 ssssssss 368
et CY
0>
C
PROBE
1 El46m"Wl
061m-500
<X'&%ss 360
'xx% 140/141
CD
3O
Co06
* 4 360
* 4 359 368 - *
4376
302'0
140 , 4*141
FIG. 6. RNasemapping of proviruses in exon III ofint-2.Poly(A)+ RNA from the indicated tumors was subjected toRNase mapping with probes thatcrossed the sitesofintegration. The location of the provirus (shown by the arrowhead) was deduced from the length of the RNase-resistant fragment(striped line) compared with theinput probe (solid line). Each insertion was mapped relative to restriction sites that marked theendsoftwoindependentprobes (Fig. 5). Theautoradiographs in the lower part of the figure give examples of the data obtained with probes1.3(a), 1.1 (b), and 1 (c) on the indicated tumors, with the respective probe and product depicted graphically above each panel. Thesizes of the protected fragments (innucleotides), indicated to the left and right of the panel, were calculated relative to standard markers andaDNAsequenceladder.
motermay overlap (D. Grinberg and J. Thurlow, personal
communication), itis possible that MMTV influencesgene
expressionthrough a common element.
MMTV insertions that perturb int-2 promoter usage. In
mostint-2-expressing mammary tumors, the MMTV
provi-rusliesoutside the transcription unit and the patterns of int-2
transcription are very similar if not indistinguishable from
those describedherefortumorE155A(3, 16, 19).However,
thereare also examples that expressanomalous transcripts
resulting from proviral insertions in the 5' and 3'
untrans-latedregions of the gene.Inthreeofthese,W26,E115, and D157B, theprovirushas been accurately located within the int-2 promoterregions by sequencingacross the virus-host
junctions. By combining this information with detailed
RNaseprotection analyses,wehave accountedfor themajor
int-2transcripts in thesetumors.
Inthe firstexample, W26, int-2expressionis activatedby
a single MMTV LTR in exonla. We previously concluded
that themajor site for int-2 transcription in this tumor was
likelytobe theviral promoter in the LTR, basedon primer
extensionanalysis(11; datanotshown). Herewe confirmed
this conclusion but showed that transcripts initiated in the
LTR may havetwo structures, depending on whetherthey
undergo splicing between exons la and Ib. The calculated
size of the major spliced transcript wouldbe 2.75 kb, very
similar to that estimated from Northern blotanalysis (Fig.
lb), whilethelessabundantunspliced form wouldbe around
3.2 kb. However, there is also evidence for transcription
initiation outside theLTR,mostlyfromasite in themost5'
region ofP2, yielding an approximately 3.0-kb RNA. The
reason for initiation at this unusual cap site is unclear but
mayreflect influences of the MMTV enhanceron the most
proximal cap site in the P2 promoter. A bidirectional
en-hancermightalso have been
expected
toactivatetranscrip-tion from thenearbyP1promoter, butnoinitiationatP1was
discernibleintumorW26.
Nevertheless,
it is also clear thatVOL.64, 1990 791
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.134.477.72.461.2]W26 contains minor int-2-specific transcripts for which we cannot as yet provide a full explanation, other than the possibility of unspliced orpartially spliced forms.
WithtumorD157B,therewasinsufficient RNAtoconduct
a comprehensive analysis, but Northern blots indicated a
transcription pattern similar to that in W26 (Fig.
lb).
Thiswould be consistent with transcription initiation from the viral promoter, presumably in the 3' LTR of thefull-length
MMTV provirus present in this tumor. Since the provirus liesintheupstream partof the P2domain, it has thepotential to subvert transcription from both P1 and P2 and may also
override P3 by promoter occlusion effects (1, 6).
In tumorE115, on the other hand, the ostensibly intact
provirus is in the inverse orientation and cannot program transcription from the normalviralpromoter. Nevertheless,
the RNase protection experiments we present here suggest thatthemajor transcriptinthistumormustinitiatewithin the provirus, presumably at some cryptic promoter on the
antisense strand of viral DNA. The position of the provirus wouldpreventinitiation inP2,and therewasnoevidencefor
transcription fromP1 cap sites. Thus, we conclude that the major2.8-kbRNA species consistently seenin thistumoris
initiated inthe provirus, while the less abundant2.7-kb RNA isanormal P3-derived transcript.
In the only other system in which comparable analyses have been reported, the activation of c-myc by avian and
murine leukemia viruses, there are close analogies to all threeof these scenarios,although thetranscriptionpattern is inherentlyless complex(8, 12, 20, 21, 23). Thus, proviruses havebeenfound within the firstexonorintron thatcandrive
expression from the viral promoter, enhance transcription fromthe c-myc promoters, orinduce initiation from cryptic
promoters. Although the generalities of the enhancer and promoterinsertion models stillapply (reviewed inreference 15), itis nowclear that the influences of retroviral LTRs on
proto-oncogene transcription can be manifold.
MMTV insertions thatperturb transcriptional termination.
Additional complexityin the transcription ofthemouse int-2 genearisesthroughusageoftwoalternativepolyadenylation signals, bothvariants oftheconsensus(10, 11, 24).Although
we cannot rigorously exclude some influences on RNA
stability or translational efficiency, it seems likely that this
complexity is largely irrelevant. InEC cells, up to 50% of transcripts terminate at the upstream poly(A) site (10, 24),
but in MMTV-induced mammary tumors, the majority of int-2 transcripts terminate at the downstream site, and the 1.8- and 1.6-kb RNA size classes are barely discernible on
Northern blots (Fig. 1). One possible explanation wouldbe
thatthe transcriptioncomplexes thattraverse thegenehave
different components in different cell types or in
MMTV-infected cells. Alternatively, the rate of transcription may
simply be higher in tumors than inEC cells, making it less
likelytoachieveterminationattheupstream signal. Insome
tumors, for example, E146, there is readthrough of both
polyadenylation signals, andthree sets of transcripts canbe
resolved (Fig. lb). Here the provirusis very close tothe 3'
end ofthe transcription unit (Fig. 5 and 6), allowing
termi-nation at the normal signals within the viral LTR and
extendingeach class ofRNAby the length ofthe U3region (-1.2 kb). In other tumors, a similar phenomenon occurs,
buttheproviruslieswithinthe transcription unit (Fig. 5 and
6).Depending onthelocation ofthe provirus relativetothe
two polyadenylation signals, this can result in extended
forms of the 2.9- and 2.7-kb RNAs but normal 1.8- and
1.6-kb transcripts,asfortumorsV97A,C46, and W106. The latter example is remarkable in that the integration has
occurred very close
(within
afewbases)
tothe downstreampoly(A) addition site.
Alternatively,
asin D124andZ79,
theprovirus lies upstream ofboth
signals,
resulting
inonly
onesetoftranscripts (Fig. 5 and
6).
The loss ofint-2 sequencesfrom the 3' end ofthese RNAs is
compensated
forby
the addition of viral sequences,making
their sizes very similarto the normal 2.9- and 2.7-kb species
(Fig.
lb).
Proviral insertion does not perturb the
protein-coding
do-main. Retroviral activation of cellular proto-oncogenes, whether resulting fromtransductioninto the viral genome or
insertional mutagenesis by integrated proviral DNA, can often involve structural alterations to the gene
product
(reviewed in reference 15). However, there are also
exam-ples of insertionally activated oncogenes whose
coding
po-tential remains unaltered, implying that it is thenormalgene product that contributes to tumorigenesis. Obviousexam-ples of the latter are the
int-i,
pim-J,and myc genes(12, 13,20, 23, 25), and here weestablished that int-2 belongsin this category. Despite the complexity of
transcripts
and severalexamplesofproviral insertions that disturb the
transcription
unit, all the proviruses mapped so far lie outside the open readingframethatspecifiesthe int-2protein. Atthe 5' endof
the gene, the most intrusive provirus (E115) is
approxi-mately 275 nt upstream of the presumed AUG initiation
codon. Although there is recent evidence for initiation at a
non-AUG codon, leading to amino-terminally extended formsof theprotein (P. Acland, M. Dixon, G. Peters, andC.
Dickson, submitted for publication), these would still be unaffected byinsertions upstream of P3. At 3' end, the most intrusive provirus that has been mapped to date, in tumor
D124, is -150 nucleotides downstream of the termination codon. Although we cannot rule out pointmutations orsmall alterations in thecoding sequences, we wouldconclude that the contribution of int-2 to oncogenesis is a function of elevated or inappropriate expression of the normal gene product. It has recently been shown that the product can function as a mitogen for cultured mammary epithelial cells and fibroblasts, in line with its classification as a member of the fibroblast growth factor family (4, 14). Similarly, under appropriate circumstances, the gene can induce morpholog-ical transformation of NIH 3T3 cells (R. Deed, W. Walther, C. Dickson, and G. Peters, unpublished data) and lead to epithelial hypertrophy in transgenic mice (W. J. Muller, F. S. Lee, C. Dickson, G. Peters, P. Pattengale, and P. Leder, submitted for publication). However, the normal role of int-2 is more likely to be as an inductive agent modulating the growth or differentiation of specific embryonic lineages (26, 27). The effect of MMTV integration, in
whatever
orientation and location relative to the int-2 transcription unit, is to reactivate the expression of a fetal gene in an adult tissue but not to alter the specificity of the gene product.
ACKNOWLEDGMENTS
We are grateful to Robert Moore, Mark Dixon, Marysia
Placzek,
and JanWitkowskifor contributing to some aspects of this work and to Roger Watson and Steve
Goodbourn
for helpful discussions on the manuscript.LITERATURE CITED
1. Cullen, B. R., P. T. Lomedico, and G.
Ju.
1984. Transcriptional interference in avian retroviruses-implications for the pro-moter insertion model of leukaemogenesis. Nature (London)307:241-245.
2. Dickson, C., and G. Peters. 1987. Potential oncogene product related to growth factors. Nature (London) 326:833.
3. Dickson, C., R. Smith, S. Brookes, and G. Peters. 1984.
on November 10, 2019 by guest
http://jvi.asm.org/
ANOMALOUS int-2 RNAs AFTER PROVIRAL INSERTION igenesisby mouse mammary tumor virus: proviral activation of
a cellular gene in the common integration region int-2. Cell 37:529-536.
4. Dixon, M., R. Deed, P.Acland, R. Moore, A. Whyte, G. Peters, and C. Dickson. 1989. Detection and characterization of the fibroblastgrowth factor-related oncoprotein INT-2. Mol. Cell. Biol. 9:4896-4902.
5. Dudof, K. P., and R. P. Perry. 1986. Properties of a mouse ribosomal protein promoter. Proc. Natl. Acad. Sci. USA 83: 8545-8549.
6. Emermann, M., and H. M. Temin. 1984. Genes with promoters in retrovirus vectors can be independently suppressed by an epigenetic mechanism. Cell 39:459-467.
7. Jakobovits, A., G. M. Shackleford, H. E. Varmus, and G. R. Martin. 1986. Two proto-oncogenes implicated in mammary carcinog-enesis, int-i and int-2, are independently regulated during mouse development. Proc. Natl. Acad. Sci. USA 83: 7806-7810.
8. Linial, M., and M. Groudine. 1985.Transcription of three c-myc exonsis enhanced in chicken bursallymphoma cell lines.Proc. Natl. Acad. Sci. USA 82:53-57.
9. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: a laboratory manual, p. 202. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
10. Mansour, S. L., and G. R. Martin. 1988. Fourclasses ofmRNA areexpressedfrom themouse int-2 gene, amemberoftheFGF genefamily. EMBOJ. 7:2035-2041.
11. Moore, R., G. Casey, S. Brookes, M. Dixon, G. Peters, and C. Dickson. 1986.Sequence,topographyand proteincoding poten-tial ofmouse int-2: a putative oncogene activated by mouse mammary tumourvirus. EMBO J.5:919-924.
12. Nottenburg, C., and H. E. Varmus. 1986. Features of the chickenc-mycgenethatinfluencethe structureofc-mycRNA in normal cells and bursal lymphomas. Mol. Cell. Biol. 6: 2800-2806.
13. Nusse, R., A. van Ooyen, D. Cox, Y. K. T. Fung, and H. Varmus. 1984. Mode of proviral activation of a putative mammary oncogene (int-1) on mouse chromosome 15. Nature 307:131-136.
14. Paterno, G. D., L. L. Gillespie, M. S. Dixon, J. M. W.Slack,and J. K. Heath. 1989.Mesoderm-inducing propertiesofINT-2and kFGF: twooncogene-encoded growth factorsrelated toFGF. Development106:79-83.
15. Peters, G.1989.Oncogenesatviralintegrationsites,p.23-66.In D. M. Glover and B. D. Hames (ed.), Frontiers in molecular biology: oncogenes.IRLPress, Oxford.
16. Peters, G., S. Brookes, M. Placzek, M.Schuermann, R. Micha-lides, and C. Dickson. 1989. Aputative intdomain for mouse mammary tumorvirusonmousechromosome 7isa 5'extension ofint-2. J.Virol.63:1448-1450.
17. Peters, G., S. Brookes, R.Smith, and C. Dickson. 1983. Tumor-igenesis bymousemammary tumorvirus: evidencefora com-mon region for provirus integration inmammary tumors. Cell 33:369-377.
18. Peters, G., and C. Dickson. 1987. On themechanism of carci-nogenesis bymousemammary tumor virus, p. 307-319. In D. Medina, W. Kidwell, G. Heppner, and E. Anderson (ed.), Cellular and molecular biology ofmammary cancer. Plenum Publishing Corp., NewYork.
19. Peters, G., A. E. Lee, and C. Dickson. 1986. Concerted activa-tion oftwopotentialproto-oncogenesin carcinomasinducedby mouse mammary tumourvirus. Nature(London)320:628-631. 20. Robinson, H. L., and G. C. Gagnon. 1986.Patterns ofproviral insertion and deletion in avian leukosis virus-induced lympho-mas.J. Virol. 57:28-36.
21. Shih, C.-K., M.Linial,M. M.Goodenow,and W. S.Hayward. 1984. Nucleotide sequence 5' of the chicken c-myc coding region: localization ofanoncodingexon that is absentfrommyc transcripts in most avian leukosis virus-induced lymphomas. Proc. Natl.Acad.Sci. USA81:4697-4701.
22. Selten,G., H. T.Cuypers,and A.Berns. 1985. Proviral activa-tion of the putative oncogenePim-1 in MuLV induced T-cell lymphomas. EMBOJ.4:1793-1798.
23. Selten, G.,H.T.Cuypers,M.Zijlstra,C.Melief,andA. Berns. 1984. Involvement ofc-myc in MuLV-inducedT cell lympho-masinmice:frequency andmechanisms of activation. EMBOJ. 3:3215-3222.
24. Smith, R., G. Peters, and C. Dickson. 1988. Multiple RNAs expressedfrom the int-2 gene in mouseembryonal carcinoma cell linesencode aprotein with homologytofibroblastgrowth factors. EMBOJ.7:1013-1022.
25. Van Ooyen, A., and R. Nusse. 1984. Structure and nucleotide sequence of the putative mammary oncogene int-1; provira!
insertions leave the protein-encoding domain intact. Cell 39: 233-240.
26. Wilkinson,D.G.,S.Bhatt,and A. P. McMahon. 1989.
Expres-sion patternof the FGF-related proto-oncogeneint-2 suggests multiple rolesinfetaldevelopment. Development105:131-136. 27. Wilkinson, D.G., G.Peters, C.Dickson,and A. P. McMahon. 1988.Expression of the FGF-relatedproto-oncogene int-2 dur-ing gastrulation and neurulation in the mouse. EMBO J. 7: 691-695.
VOL.64, 1990 793