JOURNAIL OFVIROLOGY, Jan.1992, p.562-566 Vol.66, No. 1
0022-538X/92/010562-05$02.00/0
Copyright © 1992,AmericanSocietyforMicrobiology
The
Unique
Sequence
of
the
Herpes
Simplex
Virus
1
L
Component
Contains
an
Additional Translated
Open
Reading
Frame
Designated
UL49.5
DAVID E. BARKERAND BERNARDROIZMAN*
Marjorie B. Kovler ViralOncologyLaboratories, University of Chicago, 910 East 58thStreet, Chicago, Illinois 60637
Received21August1991/Accepted 14 October1991
We present evidencefortheexistenceofanadditionalherpes simplexvirus 1genedesignated UL49.5. The
sequence,located betweengenesUL49andUL50,predictsahydrophobic proteinwith 91 amino acids.Attempts to delete UL49.5 were not successful. To demonstrate thatUL49.5is expressed, we made two recombinant viruses. First, weinsertedinframeanoligonucleotideencodinga 15-amino-acidepitopeknown to reactwith
a monoclonal antibody. This gene, consisting of the authentic promoter and chimeric coding domain, was
inserted intothethymidine kinase gene ofwild-type virus andininfectedcellsexpressedaproteinwhichreacted withthemonoclonal antibody. Thesecond recombinant virus containeda5'
UL49.5-thymidine
kinasefusion gene. The protein expressed by thisvirus confirmed that the first methionine codon ofUL49.5 served asthe initiating codon. The predictedaminoacid sequence ofUL49.5isconsistent with the knownpropertiesofNC-7,a smallcapsid protein whose gene has notbeenpreviously mapped. A homologofUL49.5 ispresentin the genomeofvaricella-zoster virus, locatedbetween homologsofUL49and UL50.
The genomeofherpessimplexvirus 1(HSV-1) consistsof twounique sequences, UL and
Us,
eachflanked by inverted repeats. Theinverted repeats of UL have been designated abanda'b',whereas thoseof Us have been designateda'c'and
ca(15). Earlier studieshave shown that the genome encodes
atotal of 75 open reading frames (ORFs), i.e., 12 in Us, 57 in UL, and 6 in the inverted repeat sequences (3, 9, 11). In
this communication, we report that UL encodes an
addi-tional gene which we have designated UL49.5.
Thecircumstances whichled us tothisconclusionwere as
follows. We have been systematically probing the HSV-1
genome to determine the role of the viral genes in viral replication and in recent studies demonstrated that the ORFs
UL46, UL47, UL ULS1, ULSS, and ULS6 are dispensable
with respect to replication in vitro (1, 12). To determine the role of UL49, we took advantage of recombinant virus R7101, inwhichthe BamHI D'fragment had been replaced
bya27-tk, achimericgeneconsisting of the promoter of the
cx27 gene fused to a portion of the transcribed noncoding
sequence and the coding sequence of the viral thymidine
kinase(tk) gene (1). The objective of these experiments was
todelete theinserted tk gene and the adjacent sequences in
the BamHI Ffragment situated in the R7101 virus immedi-ately adjacenttothe site of insertion of the chimeric a27-tk gene. Oneplasmidconstructed forthis purpose was derived from pRB3965 (1), into which we had previously cloned the
HSV-1(F)EcoRV Kfragment mapping at map units 0.690 to
0.727. The HSV-1 sequences in this plasmid were digested
withBamHIandBglIItodeleteBamHI-D' and the adjacent
terminal 272 bp of BamHI-F and religated to yield pRB4063. In accordance with previously described procedures for gene deletion (1, 13), rabbit skin cells were cotransfected with pRB4063 and intact R7101 DNA and the progeny virus was selected for the Tk- phenotype. These experiments were repeated several times; none of the 60+
bromodeoxy-*Correspondingauthor.
uridine-resistant isolates obtained by picking isolated
plaquescontained deletions in theBamHI F DNA
fragment.
The significance ofour results stemsfrom two consider-ations. (i)Deletion of the BamHI D' fragment from HSV-1 did not affect the ability of the resulting deletion mutants,
R7101 andR7108, to replicate, and therefore BamHI-D' does not contain sequences essential for replication in cells in culture (1). (ii) Hall et al. (7) and McGeoch et al. (11)
concluded thatUL49 initiated at map unit 0.696, i.e., down-streamfrom the 272 bp of the BamHI F fragment we had attempted to delete (Fig. 1A, lines 2, 4, and 5). One
impli-cation of our results was thatthe sequence weattemptedto
delete was contained within an element or gene that was
essential for viralreplication. This hypothesis wassupported
by studies of Hall et al. (7), who identified a small ORF and
alate
Y2
transcript which initiates inBamHI-D' atapproxi-mately map unit 0.701 (Fig. 1A, line 5). Since there wasno
evidence that the ORF wasexpressed, it was subsequently
ignored. Thetranscript has been of particular interest tous
inasmuch as its promoter was used as a prototype ofa
Y2
gene (10). Since the putative ORF was located between UL49 and ULS0, we designated it UL49.5. Because of discrepancies in the nucleotide sequences reported for the HSV-1(KOS) (7) andHSV-1(17) (11) UL49.5s, we sequenced theHSV-1(F) UL49.5 ORF cloned in plasmid pRB3965. The
HSV-1(F) sequence of UL49.5 is similar to that of
HSV-1(17); it differs from the latter in three nucleotide
replace-mentswhichresult in two conservative amino acid substitu-tions (Table 1). HSV-1(KOS) shares these two amino acids with HSV-1(F).
The UL49.5 ORF is expressed from a
Y2
transcriptwhichoriginates at map units 0.701, 110 bp to the right of the
BamHIsite, andis coterminal with the UL49 transcripts at map unit 0.677 or 0.689. Since the ORF was not previously shown to be expressed, we attempted to label the UL49.5
gene product by introducing the coding sequence for a
knownepitopeinto the coding sequence of the UL49.5 gene. We inserted into the BglII site of pRB3965 a synthetic
562
on November 10, 2019 by guest
http://jvi.asm.org/
2
--BamF UL48(TIF) UL49 BOI Sam BrnmD'
UL49.5
Y,(CTIF)
P(LU4g)
2(tL49.5) B
/
/4
,I
Bam OJL22.gH)
9 ! , _
10 I1
12 W.
Bam (UL22.gH)
0, __ - _
Xhqt -* r. _ Nco i -TT
** Y2P49.5(H943)
-,_--Alft-.|
(UL23,tK) (UL24) Bam
Sac
-_
r._
Nco/BgISac/Xho T2P iam ---f*
II/ %.
\
(UL.23,tk) B91 (UL24) Bram
_________ Bst/BgI
13 -A".. W
RV Bar
tK-UL49.5 Y2P
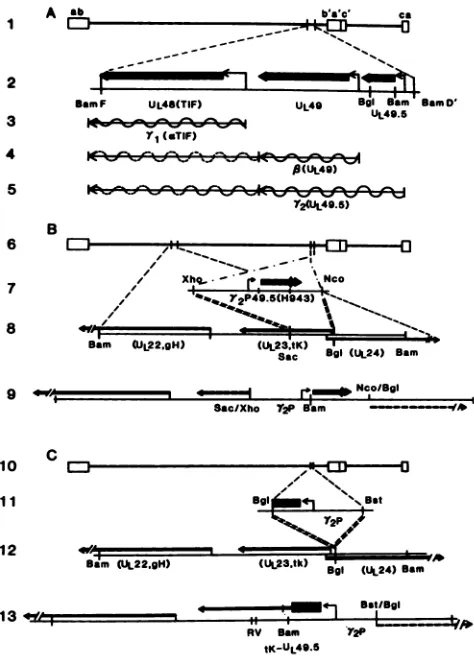
FIG. 1. Schematic representation of the genome of HSV and
HSV-1(F) (6) plasmid constructs. In panel A, line 1, the HSV-1 genome, a,b, andcarethe terminalrepetitivesequences reiterated
internally. In line 2, the positions of ORFs UL48, UL49, and UL49.5 (determined in this study) are shown. The heavy lines show the codingsequences;the thin linestothe smallarrowheadsrepresent5' transcribednontranslated RNAsequences.Lines 3, 4, and 5 show the
mRNAs ofthis partially 3'-coterminal family ofgenes (7). The lY2 transcript (line 5), which encodes UL49.5, initiates in BamHI-D', upstreamofthe UL49 gene, butcoterminates with transcripts of
UL493'ofeitherUL49orUL48(7). Panel B shows the schematics of
the construction ofrecombinant R7126. Line 7 shows the 1,300-bp XhoI-NcoIfragment ofpRB4257, which contains the late promoter and mRNAtranscriptional initiation signals from BamHI-D' and the entireUL49.5gene withthe ICP4 epitope (diamond). Line 8shows theinsertion of chimeric UL49.5 into the tk (UL23)gene.Line 9shows
thestructureofpRB4304 and the corresponding region of the R7126 recombinant. Theepitopically labeled UL49.5gene was insertedin place of the SacI-BglII sequences of the tk gene. Panel C is a
schematicrepresentation of the construction of R7127. Line 11 shows (i) the 943-bp BglII-BstEII fragment from pRB3965, whichcontains theY2 promoter and the first 80 5' codons ofUL49.5inserted into BamHI-Q (line 12)toyield thesequencearrangementofpRB4305and (ii) the corresponding domain of the R7127 recombinant. In the chimeric geneinsertedinthis recombinant, the promoter is that of UL49.5, translation initiates from the UL49.5 AUG, and the se-quencestranslatedarethoseofUL49.5 (80 codons),theremaining50 basesof thetkleader,and the tk structuralgene.
double-stranded DNA fragment made by annealing two complementary single-stranded oligonucleotides as previ-ously described (2). Thissequence encodesa15-amino-acid
epitope which occurs in infected cell protein 4 (ICP4), the
product of the a4 gene, and is recognized by monoclonal
UL49.5S of three strains ofHSV-1
Amino acid Amino acidin:
residue no. HSV-1(F) HSV-1(17) HSV-1(KOS)a
28 His Arg His
43 Gly Gly Asp
47 Glu Glu Gly
51 Ala Thr Ala
90 His His Gln
a Comparison of the HSV-1(KOS) UL49.5 sequence (7) with those of HSV-1(F)andHSV-1(17)suggested that the former contains two frameshifts. To enable comparison of the corresponding sequences of the gene, the sequence was amended by addition of two nucleotides present in both HSV-1(F) and HSV-1(17) to keep it in frame throughout the UL49.5gene.
antibody H943 (8). The HSV-1 sequences of the resulting plasmid (pRB4257) were sequenced to ensure that the in-sertedepitope wasin frame with the putative UL49.5gene. Cotransfection of pRB4257 and R7101 DNA, followed by selection for the Tk- phenotype (13), failed to yield a recombinantvirus containing theinsert. Given its small size and our failure to delete the UL49.5 gene in our earlier attempts, we hypothesized that the inserted epitope inter-fered with the function of the UL49.5 gene and that ifwe
altered asecond copy of the gene, leaving the native copy intacttoprovide its essentialfunction, wewouldmorelikely succeed in expressinganepitopically labeled product.
Plasmid
pRB4257,
described above, was digested with XhoI(map unit 0.706) and NcoI (map unit 0.697)toreleasea DNAfragment approximately 1,300 bp long which contained the _Y2 promoter and the coding sequences ofthe putative UL49.5gene,whichwasepitopically taggedattheBglII site (Fig. 1B, line 7) as described above. This fragment ,wasinserted into the BglII site of pRB3982afterfilling in of the ends with T4 DNA polymerase. pRB3982 contains the BamHI Q fragment, from which the 501-bp SacI-BglII fragment containing portions of the transcribed noncoding
sequence and thecoding sequence of the tkgene had been
replaced with a polylinker (Fig. 1B, line 8) to generate
pRB4304. Recombinant virus R7126 was generated by
cotransfection of rabbit skin cells with pRB4304 and intact HSV-1(F) DNA, and the progeny of the transfection was selected for theTk- phenotype. Figure 2 shows photographs of ethidium bromide-stained, electrophoretically separated BamHI (panelA) and EcoRV(panel B) digests ofR7126 and
autoradiograms of these digests transferredtonitrocellulose andprobed with EcoRV-K andBamHI-Qlabeled with32Pby nick translation. As shown inFig. 2A, lanes2and5, R7126
DNA contained the wild-type BamHI F, L, and D'
frag-ments, which hybridized with the EcoRV-K probe.
How-ever, the BamHI Q fragment was altered by inclusion of additional sequences which contained an internal BamHI
site. As illustrated in Fig. 1, line 9, the sequences inserted
into the BamHI Qfragment of HSV-1(F)would beexpected
tocontaintheBamHIsiteatmapposition 0.700,thatis, the site locatedimmediately upstreamofUL49.5. Aspredicted, the DNAfragments generated by cleavage ofthe chimeric
BamHIQfragmentcontainedapproximately 1,207and3,170 bpandmigrated just below BamHI-C' and-R, respectively (Fig. 2, lanes 2 and 5). Similar conclusions were reached
from analyses ofanEcoRV digest of R7126 DNA. In this instance (lanes 10 to 12), the BglII-SacI deletion in the tk generemoved the native EcoRV siteswithin the tk gene.The
insertlacked EcoRV sites, and therefore the chimeric
frag-3
4
5
6
7
8
--1--v -q
VJAW
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.68.305.73.401.2] [image:2.612.323.566.98.177.2]J. VIROL. 564 NOTES
A.
BamHI
B.
EcoRV
IL L
- CDtN --CD N
LL LL
C>
4 04y 14N
Crl)
N"
NC .0
N Cl)
14
.0
Cl ~
en IL 0
ICl
+
XL
XI oe N cn N
I
Yp- ¶- v
Nll CD) N% Cl)
m x ccX
67
1 2 3 4 5 7 d 9 10 11 12
FIG. 2. Photographs of digests ofviralDNAselectrophoretically separated in agarosegels and stained with ethidium bromide and autoradiograms of theelectrophoreticallyseparated digests obtained after hybridization withselected probes (1). Recombinant viruses R7126 andR7127weredescribed in Fig. 1. The probes,theEcoRV Kfragment(map units 0.690to0.727), and BamHI-Qwerelabeled with32Pbynick translation. Letterdesignations of the restriction fragmentsappeartotheleft. Forpanel A, viral DNAsweredigested
with BamHI (NewEngland BioLabs). In lanes 4,5, and 6,
hybrid-ization ofEcoRV-K to native BamHI-F, -L, and -D' in all three strains indicates that the native copy of UL49.5 is intact. The
electrophoretic mobilities of BamHI Q fragments of both R7126 and R7127(lane 4)werealteredby the inserts(see Fig. 1, lines 9 and 13). Inpanel B, for which viral DNAswere digested withEcoRV (1),
EcoRV-Kremained unaltered in all three strains. EcoRV-Q and -R
werefused with theinsertedsequencesin R7126 (lane 11) because of deletion of theRVsites in theSacI-BglIIsequenceof the tkgene.In R7127 (lane 12), the SacI-BglII sequence was restored, thereby
restoring the EcoRV restriction sites absent in HSV-1(F)A305. However, in this recombinant the additional sequences were
in-sertedintoEcoRV-R (Fig. 1, line 13).
mentwaspredictedtobe 4,309 bp long andtocomigrate with
EcoRV-N. In accord with this expectation, the probes
hybridized toa DNAfragmentof this size(Fig. 2B, lanes 8
and11). Note that theDNAfragmentwhichhybridized with
BamHI-Q and migrated below EcoRV-R in lanes 10 to 12
(EcoRV-S) mappedatmapunits0.291to0.299 andtherefore
shares sequenceswith aportion of the BamHI Q fragment. Analyses oflysatesof cellsinfected withtwoindependent
isolatesofR7126orwith theprogenyoftransfection which weregenotypically identicalto HSV-1(F) (designated aand
43
30
1 2 3 4 5 6 7 8 9
10
FIG. 3. Photographs of electrophoretically separated proteins
from lysates of Vero cells infected with 5 PFU of HSV-1(F) or
recombinantvirus percell,electricallytransferredtonitrocellulose sheets, and stained with monoclonal antibody H943 to ICP4(left panel)orrabbit immuneserumagainstthymidinekinase. Theright columnwas aspreviouslydescribed(14).Thecellswereharvested at18h, andelectrophoresiswasdoneon18%(lanes 1to4)or8% (lanes5to10)bisacrylamide gels.Thepositionsofmolecularweight
(103) standard proteins (Pharmacia) are shown to the left of the panels. Thecontents oflanes 1 to4 were hybridizedwith
mono-clonalantibody H943, whichreactswiththea4epitopeintroduced intoR7126. Thedetection system wasalkaline phosphatase-conju-gatedgoatanti-mouseantibody(Promega).Lanes 1and2show the
new12,000-MrantigenproducedbytheepitopicallylabeledUL49.5
gene, while lanes 3 and 4 contained proteinsfrom isolates ofthe same transfection that were genetically identical to HSV-1(F),
demonstratingthatneither transfectionnorselection is sufficientto induce theH943-detectable antigen.Thebandsatthe topofthegel
contained ICP4, whichalso reacts with the H934antibody.Lanes 5 to 10 show the profiles of electrophoretically separated proteins
reacted firstwithpolyvalent rabbit anti-thymidine kinaseserumand then with alkaline phosphatase-conjugated goat rabbit anti-body.Theanti-rabbit antibody reactedwith thewild-typeproteinin HSV-1(F)(lanes5, 7, and 9)orwiththechimericUL49.5-tkprotein
made in cells infected with R7127. The lanes were loaded with equivalentamountsof infected celllysates. The relativeintensityof the 53,000-Mr band versus that of the 41,000-Mr band in lane 6 suggests that the upstream initiationAUG ofUL49.5 is preferred
overthatof the tk codon fortranslationalinitiation. For lanes 7and 8, the infected cells were maintained in the presence of phosphonoacetate (PAA;300,ug/ml; Sigma).Forlanes 9and10, the infectedcellsweremaintained at39°C.
b) were electrophoretically separated on denaturing 18%
bisacrylamide gels, electrically transferred to a
nitrocellu-lose sheet, and reacted first with the H943 monoclonal
antibody and then with alkaline
phosphatase-conjugated
goatanti-mouseantibody (Promega)aspreviously described
(14). Theantibodyreacted withaprotein band withanMrof
approximately 12,000 (Fig. 3, lanes 1 and 2), i.e., in good
agreement with the Mroftheprotein predicted onthebasis
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.313.553.72.337.2] [image:3.612.55.294.77.412.2]by
theinsertedepitope.
The dark bandsatthe top of the gel containedICP4,
which reacted with H943 and enteredpoorly
intothegel.
The second recombinant virus was constructed to
deter-mine whether the initiation codon was the first ATG of
UL49.5
or,assuggested by
Halletal.(7),theinitiatingcodonwasthedownstreamUL49ATG. Inthisinstance,the 943-bp
fragment,
which contains the Y2 promoter and the first 80codons
Of
UL49.5 (Fig. 1C,
line 11), was removed frompRB3965 by cleavage
with BglII and BstEII at map units0.698and
0.704, respectively,
andinserted intotheBglIIsite ofthetk geneascloned inpRB173 (Fig.
1C,line12). Inthisinstance,
both thefragment
derived from UL49.5 and thepRB173-derived
vector weretreated withT4DNApolymer-ase, to which dGTP was added. As a consequence, the
enzymefilledthefirst base
complementary
tocytosinein theoverhanging
ends but removed the otherunpaired
bases toplace
theUL49.5
and tk genes in the same translationalreading
frame. In theresulting plasmid,
pRB4305 (Fig. 1C,line
13),
initiation oftranslation atthe first ATG ofUL49.5would
yield
a chimeric protein consisting of the 80 aminoacids of
UL49.5,
17 codonsrepresenting
the translatedproduct
ofthe 5' transcribednoncoding
sequence ofthe tkgene downstream from the
BglII
site,
and thecoding
se-quencesofthe tk gene
(Fig. 1C,
line13). Since deletionsandinsertions intothe 5' endoftk donotabolish the activity of
the enzyme
product,
wecotransfectedrabbit skin cellswithpRB4305
and intact tk[HSV-1(F)A305)]
DNA. The tk+progeny selected as
previously
described wasplaque
puri-fied and
designated
R7127.As shown in
Fig.
1C,
line13,
it could beexpected
thatinsertion of the
943-bp fragment
into the tk gene wouldintroduce the BamHI sitefrom map
position
0.700nearthe5' terminus ofthe
UL49.5
sequence. Thepredicted
chimericBamHI
Q fragments
generated by digestion
with thaten-zyme would contain
1,449
and3,073
bp, respectively,
ex-actly
as found(Fig.
2,
lanes 3 and6).
Restoration of theBglII-SacI
sequence frompRB4305
intoBamHI-Q
restoredto themutated tk genefrom
HSV-1(F)A305
the EcoRVsitesatmapunit 0.312. Asaconsequence,the EcoRV
Q fragment
comigrated
withthewild-type fragment (Fig.
2B,
lanes 7 and10).
Theadditional 943bp
frompRB4305
werepredicted
toincrease the size of the EcoRV R
fragment
to2,436
bp,
which would
place
itslightly
below EcoRV-P(2,691
bp)
inFig.
2,
lanes 9 and 12. Theintensity
of thehybridization
signal
ofthenewbandinlane 12isduetothefactthat allofthe 943
bp
oftheUL49.5
insertwererepresented
in themorehighly
labeled EcoRV-Kprobe,
whereas the restoredse-quences of
EcoRV-Q
werehighlighted
by
the lesshighly
labeled
BamHI-Q probe.
Figure
3,
lanes 5 and6,
shows theelectrophoretic
mobil-ities ofthe native
HSV-1(F)
andchimeric R7127thymidine
kinasesasdetected
by
the R161polyclonal
rabbitantiserummade
against
apeptide predicted by
theHSV-1(F)
tkse-quence. The cells were harvested at 18 h
postinfection,
electrophoretically separated
in8%bispolyacrylamide gels,
transferred to a nitrocellulose
sheet,
and reacted with thepolyclonal
serum,and then thethymidine
kinasepolypeptide
wasvisualized
by staining
withalkalinephosphatase-conju-gated
goat anti-rabbitantibody (Promega).
Theelectropho-retic
mobility
of theresulting
protein predicted
anMr
of53,000
(Fig. 3,
lanes 5 and6),
indicating
that translationbegins
atthefirstAUG.The chimeric
UL49.5-tk
protein
wasexpressed
fromalatepromoter
(Fig.
1)
and therefore could beexpected
to be2E
1.4
VZV ORFU5
HSV-10149.5
-24
0. 10.0 20.0 30.0 40.0 50.0 80.0 70.0 80.0 90.0
Ain AddNumbw
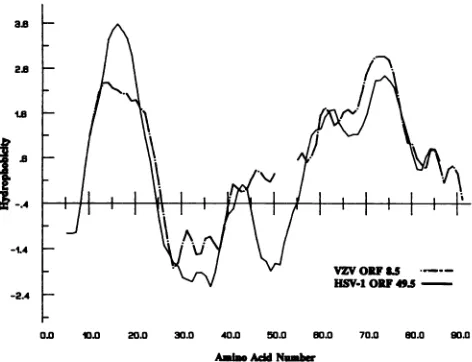
FIG. 4. Kyte and Doolittle hydropathic analysis oftheproteins
predictedto beencodedbythe HSV-1UL49.5 gene andVZVORF
8.5, whichishomologous to UL49.5. The sequences were scanned
by using a moving window ofseven residues and plotted with a three-residuesmoothing function.Toalign theoutputs,five-residue gapswereintroduced into the5' endnearthecenteroftheORF8.5 sequence.
dependenton viral DNAsynthesis. Addition of
phosphono-acetic acid, an inhibitor of viral DNA synthesis, did not
significantly inhibit production of native
p-tk
fromHSV-l(F)-infected cells butsignificantly depressed production of
UL49.5-tk
in R7127-infected cells(Fig.
3, lanes 7 and 8).Similarly,
inasmuch as ICP4 of HSV-1(F) contains a tslesion, at the nonpermissive temperature, UL49.5-tk
chi-meric
protein
synthesiswasvirtually eliminated,aswould beexpected ofalate
(Y2)
gene (Fig. 3, lanes 9 and 10).McGeoch et al. (11), in the original publication of the sequence,predicted conservatively,onthebasis of available
data,atotal of 72ORFs. Sincethen,ithas been shown that
the ORF originally designated UL26 in factconsists oftwo
3'-coterminal
transcriptional units,
UL26 andUL26.5,
eachyielding
aprotein
product (9). Another ORFexpressing
aprotein
was found in the inverted repeatsflanking
the ULsequence,between the terminala sequenceand the aOgene
(2, 3). This gene, present in two
copies
per genome, wasdesignated
1y34.5
(3).Discovery
of the UL49.5gene bringsthetotal number of known ORFsto 76.
The UL49.5gene productis expectedtobe
highly
hydro-phobic (Fig.
4). Its functions are not known.However,
thepredicted
sequenceof theUL49.5protein
is similartothat ofasmall
capsid protein designated
NC-7(4),
whose gene hasnotbeen
mapped.
NC-7 wasreported
to have an apparentmolecular
weight
ofapproximately 12,000,
at least twomethionineresidues intwodifferent
tryptic peptides,
andatleast five
arginine
residues.UL49.5
ispredicted
to contain four in-framemethionines,
the first ina five-residuetrypticpeptide,
the other three in alarge
39-amino-acid peptidederived from the
carboxyl
halfof theprotein,
and six(or
seven)
arginines.
Trypsin digestion
wouldyield
sevenpep-tides,
of which thecarboxyl
terminus(seven
aminoacids,noarginines)
and thesingle
arginine
atresidueposition
6might
notbe
detected,
but the other fivepeptides, ranging
from 3to39amino
acids,
should have been detectedby
Cohen etal.(4), who
reported
four to sixarginine-labeled
tryptic
pep-tides.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.323.559.73.254.2]566 NOTES
The UL49.5 ORF is conserved in thegenome of varicella-zoster virus (VZV). The homologs of HSV-1 ORFs UL49 andUL50areVZVORFs9 and 8. Between ORFs 8 and 9, positioned nearly identically to HSV-1 UL49.5, there is a small ORF (VZV ORF 8.5) containing 87 codons which overlaps the first 9 codons of ORF 8 (5). Incontrast, HSV-1 UL49.5 contains 91 codons and is separated from ULSO by 23 bp. The relatedness between HSV-1 UL49.5 and VZV ORF
8.5 is particularly striking if expressed in the form of a Kyte-and-Doolittle hydropathic plot (Fig. 4).ThatUL49.5is
produced fromatruelatepromoter(7, 10; R7127data in this report) is consistent with its predicted role as a virion (capsid) protein. Were the product of the UL49.5 gene a capsid protein, it would be predictedtobe essential for virus maturation andnotdispensable for replication, even in cells
inculture, asreported here.
Wethank Suzanna Rudofsky forexperttechnical assistance and LenorePereira for the invaluable gift of monoclonal antibodies.
These studies were aided bygrants from the National Cancer Institute (CA47451) and the National Institute for Allergy and Infectious Diseases (AI124009 and A11588), United States Public HealthService.
REFERENCES
1. Barker, D. E., and B. Roizman. 1990. Identification of three
genes nonessential for growth in cell culture near the right
terminus of the unique longsequencesof thelongcomponentof herpes simplex virus 1. Virology 177:684-691.
2. Chou, J., E. R. Kern, R. J. Whitley, and B. Roizman. 1990. Mapping of herpes simplex virus-1 neurovirulence to-y34.5, a
genenonessential forgrowth in culture. Science 250:1262-1266.
3. Chou, J., and B. Roizman. 1986. The terminalasequenceof the
herpes simplex virusgenome contains thepromoterofa gene
located in therepeat sequenceof the L component. J. Virol. 57:629-637.
4. Cohen, G. H., M. Ponce de Leon, H. Diggelmann, W. C. Lawrence, S. K. Vernon, and R. J. Eisenberg. 1980. Structural
analysis of the capsid polypeptides of herpes simplex virus types 1 and 2. J. Virol.34:521-531.
5. Davison, A. J., and J. E. Scott. 1986. Thecompletesequenceof varicella-zoster virus. J. Gen. Virol. 67:1759-1816.
6. Ejercito,P.M.,E. D.Kieff,andB. Roizman. 1968. Characteri-zation ofherpessimplex virus strainsdiffering in their effectson the socialbehavior of infected cells.J. Gen. Virol.2:357-364. 7. Hall, L. M., K.G. Draper, R. J.Frink, R. H.Costa,and E.K.
Wagner.1982.Herpessimplex virus mRNA speciesmapping in EcoRIfragmentI. J. Virol. 43:594-607.
8. Hubenthal-Voss,J., R. A. Houghton,L. Pereira, and B. Roiz-man. 1988. Mapping of functional and antigenic domains of the a4protein of herpessimplex virus1. J.Virol. 62:454-462. 9. Liu, F., and B. Roizman. 1991. The promoter, transcriptional
unit, andcodingsequence of herpessimplex virus 1family 35 proteinsarecontainedwithin andin framewith theUL26open readingframe. J. Virol.65:206-212.
10. Mavromara-Nazos, P., and B. Roizman. 1989. Delineation of regulatorydomainsof early(13)andlate(Y2)genesby construc-tion of chimeric genes expressed in herpes simplex virus 1 genomes. Proc. Natl. Acad. Sci.USA 86:4071-4075.
11. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M.C. Frame, D. McNab, L. J. Perry, J. E.Scott, and P. Taylor. 1988.ThecompleteDNA sequenceof thelongunique region in the genome of herpes simplex virus type 1. J. Gen. Virol. 69:1531-1574.
12. Meignier, B., R. Longnecker, and B. Roizman. 1988. In vivo behavior of genetically engineered herpes simplex viruses R7017 and R7020. I. Construction and evaluation in rodent animal models.J. Infect. Dis. 158:602-614.
13. Post, L. E., and B. Roizman.1981.Ageneralized technique for deletion ofspecificgenes inlargegenomes:agene 22ofherpes simplex virusis not essentialforgrowth. Cell25:227-232. 14. Purves, F. C., D. Spector, and B. Roizman. 1991. The herpes
simplexvirus 1protein kinase encoded by the Us3gene medi-ates posttranslational modification ofthe phosphoprotein en-codedby the UL34gene.J. Virol. 65:5757-5764.
15. Wadsworth,S.,R.J. Jacob, and B. Roizman. 1975. Anatomyof herpessimplex virusDNA.II. Size,composition, and arrange-mentof invertedterminalrepetitions. J.Virol. 15:1487-1497.
J. VIROL.