CopyrightX 1992, American Society for Microbiology
Basis for Receptor Specificity of Nonecotropic Murine
Leukemia Virus Surface Glycoprotein
gp7OSu
DAVIDOTT* ANDALAN REIN
Laboratory ofMolecular Virologyand Carcinogenesis, ABL-Basic Research Program, NCI-Frederick CancerResearch andDevelopmentCenter,
Frederick,
Maryland 21702-1201Received 14February 1992/Accepted 21April 1992
Murineleukemia viruses (MuLVs) initiate infection of NIH 3T3 cells by binding of the viral envelope (Env) protein to a cell surface receptor. Interference assays have shown that MuLVs can be divided into four groups, eachusing a distinct receptor: ecotropic, polytropic, amphotropic, and1OAl.Inthis study, we have attempted tomapthe determinants within viral Env proteins by constructing chimeric env genes. Chimeras were made in all six pairwise combinations between Moloney MCF (a polytropic MuLV), amphotropic MuLV, and1OA1, using a conservedEcoRIsite in the middle ofthe Env coding region. The receptorspecificityof each chimera wasdetermined by using an interference assay. We found that amphotropic receptor specificity seems to map to the N-terminal portion of surface glycoprotein
gp7OSu.
The difference between amphotropic and 1OAl receptorspecificity
can beattributed to one or more ofonly
six amino aciddifferences in this region.Nearly
all othercasesshowed evidence of interaction between Env domains in thegeneration of receptor specificity. Thus, a chimera composedexclusively
of MCF and amphotropic sequenceswasfoundtoexhibit1OAl receptor specificity. None of the chimeraswereabletoinfect cellsby using the MCF receptor; however,twochimeras containing theC-terminal portion of MCFgp7OSu
could bind to this receptor, while theywereabletoinfect cells via the amphotropic receptor. This result raises the possibility that receptor binding maps to the C-terminal portion of MCFgp7OSU but requires MCF N-terminal sequences forafunctional interaction with the MCF receptor.Like all retroviruses, murine leukemia viruses (MuLVs) initiateinfectionbyaspecific interactionbetween the MuLV surfaceglycoprotein(gp7OSU)andahost cell surfaceprotein that acts as a receptor. The attached virions are subse-quently internalized into the cell, whereinfectionproceeds. Inproductively infected cells, envelope protein (Env) mol-ecules present on cellular membranes bind the host cell receptor, effectively blocking further viral infection (20). Assembling virions receive their Env coat when they bud through the hostplasmamembrane (5).
While many different MuLVs, with a wide variety of properties, have been isolated, categorizing the known MuLVs by receptor specificity yields a limited number of groups. The use ofcompetitive interference to superinfec-tionas anassayfor receptorspecificity has established only five classes of MuLVs into which all known MuLVscanbe placed (17, 18). Four of these five classes, ecotropic, poly-tropic (MCF), amphotropic, and 10A1, all utilize different receptorson mousecells. Theremaining class, the xenotro-picMuLVs, donothaveafunctional receptoronlaboratory mouse strains.
One way ofapproachingabetterunderstandingofgp7OSU function would be to determine which regions of gp7OSU participateinthespecific interaction with cell surface recep-tors. Avariety of studies have suggested that the determi-nantsfor receptor specificity lie in the N-terminal two-thirds of gp7OSU (13, 16); in fact, Heard and Danos (11) have recently shown that an Envfragmentcontaining most of this region of Friend MuLV gp7O u can bind to the ecotropic receptorin NIH 3T3 cells.
Wehavepreviously comparedEnvaminoacid sequences from each of the five classes of MuLV. We found (15) that
* Corresponding author.
four of the five classes(polytropic, xenotropic, amphotropic, and 10A1)were remarkably similar in the N-terminal two-thirds ofgp7OSU; however, two blocks ofsequence differ-ence in this group (the polytropic-related MuLV [PRM] group) were noted. The remainder of Env is highly con-served for all MuLVs.
Oneof thepolymorphic regions is located in the first 200 amino acids of the envelope polyprotein gene (env); this region ofamphotropicand1OAl
gp7oSu
containstwo inser-tions of amino acid sequences relative to the MCFgp7OSu
sequence. XenotropicMuLV lacksone of these insertions and hasauniquesequenceatthe other insertion. The second region consists ofaproline-rich sequence named the hyper-variableregion (12, 15).Besides thesetworegions,thereare scattered point differences between the PRMgp7OSu
se-quences.In this study,we have attemptedto map receptor speci-ficity in
gp70u
by constructing a series of chimeric env genes,using
MoloneyMCF(Mo-MCF), 1OA1, and ampho-tropicgp7O0u
sequences.Theanalysisof MuLVscontaining these chimericgp7Osu
gave both simple and complex re-sults.In somecases, receptorspecificitycould bemappedto asingle regionofgp7OSu.
Inother cases, receptorspecifict is determined by the interaction oftworegions ofgp70S
.Finally, some combinations seem to be capable of a fully functional interaction withonereceptor butalsoapartialor abortive interaction with one or two other receptors. The implications of these observationsareconsidered in Discus-sion.
MATERIALSANDMETHODS
DNA constructions. Chimeric MuLVs were based on Moloney MuLV (Mo-MuLV), with the placement of the chimeric env gene sequences into the virus by use of
4632
on November 9, 2019 by guest
http://jvi.asm.org/
conservedSall(atMo-MuLVnucleotide3705[19]) andClaI (Mo-MuLV nucleotide 7674 [19]) restriction sites found in Mo-MuLV, Mo-MCF, amphotropic 4070A,andlOAlMuLV
sequences. The chimeric env gene sequences were
con-structed by theuseofanEcoRI restriction site(amphotropic env nucleotide 625 [15]) that is conserved in amphotropic
4070A,Moloney-MCF,andlOAlsequences.TheEcoRI site
is located between the two major variable regions of the
gp7Os.
A Mo-MuLV infectious molecular clone in apSV2Neo-derived plasmid that encodes G418 resistance (8) had its3'pol andenvgenesequences removedbySalland ClaI restriction endonuclease digestion. PRM sequences were then inserted in this region by ligation ofSalI-EcoRI and EcoRI-ClaI PRM fragments in order to construct a
chimericenvgenearound the conserved EcoRI site. Sources ofPRMsequenceswere afull-lengthclone ofMo-MCF(1) (a kind gift fromM.Vogt, Salk Institute, SanDiego, Calif.), a
circularly permutedclone oflOAl (15), andan unpublished
molecular clone of 4070A (14). This clone has the same
receptorspecificity as does the4070A clone of
Chattopad-hyayetal. (3). We have determined itsenvgene sequence overthecodingregionforaminoacid residues 1to 190,367 to486,and 631to658(14).Over theseregions, thesequence wasidenticalatthe nucleotide leveltothat of the latter clone (15) exceptfor twonucleotide changesin thecodingregion for the Env leader peptide. Some chimeras were further
modified by exchanging lOAl and amphotropic hypervari-able regions. Two restriction endonuclease sites that flank thehypervariable regions, EcoRI (amphotropicenv
nucleo-tide625[15]) andBglI(amphotropicenvnucleotide 964 [15]) wereusedtoproduce theserecombinants.
Cells andviruses. CHO cellswere cultured, infected, and
harvested as described by Ott et al. (15). Genomes of chimeric MuLVs were introduced into CHO cells (which
lackendogenousMuLVsequences)orNIH3T3 cells by the calcium phosphate technique as described by Graham and
vanderEb(9) and selectedforstableintegration of chimeric
MuLV constructs by cultivation in G418 (GIBCO, Grand Island, N.Y.). Since chimeric MuLVplasmids also contain the neo gene from pSV2Neo (8), stable Neor expression
ensures stable chimeric MuLV integration. Viruses
pro-duced by CHO cells transfected with the chimeras were
either used to infect NIH 3T3 cells or used to produce Harvey sarcomavirus (HaSV) pseudotypes by
superinfec-tionofHaSV-transformed NIH3T3cells (17).
Interference tests.Superinfection interferenceassayswere
performedasdescribedpreviously (17).NIH3T3cells(105)
productively infected with an MuLV were exposed to
ap-proximately104 focus-formingunitsof HaSV.Thecellswere
incubated for 5to7daystoallowthesuperinfected NIH 3T3 cells to become confluent. Cells were examined for the presenceofHaSV-transformedfoci. Presenceof>103 foci is
interpreted to indicate lack of interference between the resident MuLV in the NIH3T3cells and the superinfecting HaSVpseudotype; presence of lessthan threefociinduced
bythe104 focus-forming particles onthechallenged cells is
interpreted toindicate superinfection interference between the twoviruses.
Radioimmunoprecipitation.Chimerasthatdidnotproduce infectious viruswere examined forthe production of
retro-viralparticles by assayingculturesupernatantsforpelletable capsid protein p3OCAand
gp7OSu,
using radioimmunoprecip-itation.CHO cells stably transfected withchimericconstruc-tions were labeled for 16 h with [35S]methionine. Culture
supernatants were collected, clarified through a
0.45-,um-pore-size filter, andcentrifugedinanSW27rotor(Beckman
Instruments, Palo Alto, Calif.) at 25,000 rpm for 90 min at 4°C in order to pellet viral particles. Viral pellets were resuspended in TNT buffer (20 mM Tris-Cl [pH 7.5], 0.2 M NaCl, 0.1% [vol/vol] Triton-X100). Immunoprecipitation was performed by addition of protein A-linked Sepharose (Pharmacia-LKB, Piscataway, N.J.) and either anti-p30CA or
anti-gp7oSu
antiserum(both described in reference 21) to the resuspended pellets. Afterincubation at 4°C for 2 h, the mixture wasmicrocentrifuged for 30 s. The Sepharose beads werewashed three times with ice-cold TNT before the beads werecombined with gel loading buffer (1% sodium dodecyl sulfate [SDS], 10%glycerol, 1%,B-mercaptoethanol, 0.001% bromophenol blue, 100 mM Tris-Cl [pH 8.6]), boiled for 5 min, and subjected to SDS-polyacrylamide gel electrophore-sis. Gels were treated with the fluor 2,5-diphenyloxazole in dimethyl sulfoxide and dried before exposure to X-ray film at -700C.Nucleotide sequence accession numbers. The Env se-quences of the 4070A clone of Chattopadhyay et al. and of lOAl are listed in GenBank under accession numbers M33469 and M33470, respectively. The long terminal repeat sequences of the 4070A clone ofChattopadhyay et al. (3) and of lOAl are listed under accession numbers M55597 and M55596, respectively.
RESULTS
Tomap regions of thegp70su molecule that contribute to the receptor specificity of the Env protein complex, we constructed chimeric env genes from amphotropic, Mo-MCF, and lOAl env sequences (Fig. 1). The constructions recombined these sequences at a conseived EcoRI restric-tionenzyme site which is located just after amino acid 200 of the Envpolypeptide. Thus, chimericgp7OSushad theamino half of MuLVgp7Osufrom oneMuLV fused to the carboxy half of
gp7OSU
and the majority of transmembrane protein p1SET of theother MuLV. Designations for these chimeras areabbreviated as follows: A for amphotropic 4070A, M for Mo-MCF, and 10 for lOA1, with chimeric envs denoted by simple combinations of the abbreviations. For example, A.10 has the N-terminal portion of amphotropic MuLV and theC-terminal portion oflOAl MuLV.The chimeric MuLVswere tested for receptor specificity bytesting HaSV pseudotypes forsuperinfection interference on NIH 3T3 cells previously infected with either ecotropic, MCF, amphotropic, or lOAl MuLV. The assay was also performed reciprocally: NIH 3T3 cells infected with chi-meric MuLVswere challenged with HaSV pseudotyped by an MuLV from one of the four classes that infect mouse cells.
Most chimeric MuLVs were also tested for their interfer-ence properties with each other. In all cases tested, the interference patterns observed between pairs of chimeras were consistent with the receptor specificities assigned to each chimera by interference patterns determined against ecotropic, amphotropic, Mo-MCF, and lOAlviruses.
Amphotropic MuLV and lOAl exchanges. While most classes of MuLV have straightforward interference proper-ties, lOAl receptor specificity is more complicated. Previ-ously, lOAl MuLV has been shown to interfere with both amphotropic and lOAl MuLVs (18). However, only lOA1, not amphotropic MuLV, interferes with
lOAI
superinfec-tion.These data indicate thatlOAlMuLV has dualreceptor specificity: it can apparently bind to both amphotropic and lOAl receptors.Chimeric Envs that exchanged sequences between
on November 9, 2019 by guest
http://jvi.asm.org/
Carboxy-Terminus
A | 10 | M
A AMPHO
E
10_a3_ ~10A1
E Nobinding MCF
FIG. 2. Three-by-threematrixshowingthereceptorspecificities ofthe sixchimeric
gp70s's.
Ntermini ofamphotropic (AMPHO), lOA1,and MCFgp70Susarelistedattheleft,andthe Cterminiarelisted above the matrix. The receptorspecificities of the combina-tions are noted inside the boxes. Wild-type combinations are in whiteboxes; combinations for which receptorspecificity is deter-minedcombinatoriallyarestippled; combinationsfor which recep-torspecificitymaps to theamino half ofgp7OSuareshown in black.
M.A-10
FIG. 1. Maps of chimeric 7OSUs. Stippled regions denote highly variable regions of gp7O as described by Ott et al. (15). Restriction enzyme sites used for the constructions within gp7OSU are also shown. The different sequences are denoted as follows: openbox foramphotropic; black box for Mo-MCF; and hatched box for lOAl.
photropicand1OAl
gp70sus
wereassayedfortheirreceptorspecificities (Table 1). In interference assays, A.10 MuLV interfered with superinfection by the amphotropic
pseudo-type of HaSV [HaSV(ampho)], while both amphotropic MuLV and1OAlinterfered withHaSV(A. 10) superinfection. Thus, theinterference properties ofA.10were identical to
those ofamphotropicMuLV. Our results with chimeras are
summarized inFig. 2.
Incontrast, infection bythe10.A chimera rendered NIH 3T3 cells resistant to superinfection by both HaSV(lOA1) and HaSV(ampho) pseudotypes, while only 1OAl-infected
cellswereresistanttoHaSV(10.A) superinfection (Table 1). Thus, the 10.A chimera had the interference properties of 1OAl MuLV(Fig. 2). Therefore, in thesetwocases, the N termini determine the receptor specificityof thesechimeric gp7osUs.
Amphotropic MuLV and Mo-MCF exchanges.The ampho-tropicandMo-MCFrecombinationsproducedchimeras with intriguing properties (Table 1; Fig. 2). Prior infection of NIH 3T3 cells with theA.M chimerarendered them resistantto
superinfection by HaSV(ampho). Conversely, amphotropic MuLV interfered withsuperinfection by HaSV(A.M). Thus, A.Mclearlyhasamphotropicreceptorspecificity. However, theA.M virus alsodisplayed nonreciprocalinterference with Mo-MCF;thatis,itinterfered withMo-MCFsuperinfection, although itwas ableto efficiently superinfect cells infected with Mo-MCF.These resultssuggestthat thechimeric A.M gp7OSu is able to bind the MCF receptor as well as the
amphotropic receptor but can use only the amphotropic
receptorforentryinto the cell(Fig. 2).
The M.A chimera also hadan unexpected property: this chimera had the same interference properties as did 1OAl
(Table 1; Fig. 2). Thus, HaSV(M.A)wasabletoinfect all cell lines except those infected with 1OAl MuLV, while cells infected with M.A MuLV were resistant to either HaSV(lOA1) orHaSV(ampho)butnotHaSV(MCF).
[image:3.612.333.532.72.147.2]Mo-MCF and 1OAl exchanges. The Mo-MCF and 1OAl recombinations also gave unexpected results. CHO cells transfected with the M.10construct did not producevirus particles capableofinfectingmouse,CHO, mink,orratcell
TABLE 1. Interferencetestingof MuLVscontaining chimericenvelopes
Foci presentafterinfection with HaSVpseudotyped
by':
NIH3T3 cells
infected with: Moloney Mo-MCF Amphotropic lOAl 1O.A A.1O A.M M.A 1O.M
(4070A)
Novirus + + + + + + + + +
Moloney - + + + + + + + +
Mo-MCF + - + + + + + + +
Amphotropic (4070A) + + - + + - - +
lOAl + + - -
-1O.A + + - - - ND
A.10 + + - + + - - + ND
A.M + - - + + - - + ND
M.A + + - - - ND
1O.M + - - - ND ND ND ND
-M.lOb + + + + + + + + ND
+,foci present; -, fociabsent;ND, notdone.
b NIH3T3 cells transfected with plasmid M.10 and selected with G418.
Amphotropic
Insertions ,+i°.Se
IS
F
iF
////////.IzZZ
P99997
gp7OSU
A.10
1O.A
M.A
A.M
1O.M
M.1O
M.1-A
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.100.247.75.340.2] [image:3.612.59.553.564.705.2]CHO M10 Mo(1OA1)
I Ir II 1
C) CL c') CL CO) CL
K L CDm CL> m
CDvl
200-
97-_ $
68
-43
-
[image:4.612.120.244.82.265.2]26-_
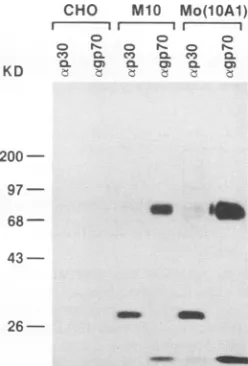
FIG. 3. Immunoprecipitation of p30CA and gp70SU. Cell lines usedareCHO,CHO transfected withM.10,andCHO infectedwith
Mo(lOA1), a chimeric Mo-MuLV that has a 1OAlenv gene (14). Antisera used for immunoprecipitation are indicated above the
lanes.
lines(datanotshown). Further,NIH 3T3 cellsstably trans-fected with the M.10 MuLVconstructwerepermissive for all of the natural or chimeric viruses tested (Table 1). By
virtue of the transfection and the G418 selectionprocedure, all of the challenged cells should have contained the M.10 construct(asnoted in Materials andMethods).The absence of any superinfection interference by these cells and the
absence of infectious virus suggest that the M.10 Envwas
functionally inactive.
The presence of both gag and env gene products in supernatantsofCHO cells transfected with M.10was
exam-ined to determine whether Env is expressed and is associ-ated with virus particles. [35S]methionine-labeled virions
were analyzed by radioimmunoprecipitation. As shown in Fig. 3, CHO cells transfected with the M.10 construct
producedvirions that containedgg70SU,asevidencedbythe
presence of bothpelletable p30 and gp7OSU. Since M.10
seems to produce virions containing correctly processed
Envproteins,the defect in the virusseemsto result froma
functional defect in the chimericgp7O.
The defectiveness of theM.10chimeric
gp70su
issurpris-ing since, asshownabove,theM.A chimeric
gp7Osu
isfully functional. This difference mustbe due to sequencediffer-ences in thecarboxy halvesof1OAl gp7OSUand
amphotro-pic
gp7OSu.
Weattemptedtoidentifytheregion responsible for the defectiveness of M.10 by making exchanges within thecarboxyhalf ofgp7Osu.
Byuseof conserved EcoRI and BglI sites, the hypervariable region and its immediate 3' flanking sequences of env were alternately exchangedbe-tween1OAlandamphotropic4070AtoproduceM.A-10and M.10-A (Fig. 1). Neither of these chimeras gave rise to infectiousviruses, althoughcells containingthemproduced
virusparticlesdetectableby immunoprecipitationwith
anti-p30CAand
anti-gp70Su
antisera(data notshown). Asimilarsetof constructions between 1OAl andamphotropic MuLV thatyielded 10.A-10andA.10-Awerefullyfunctional (data
not shown), showing that the A-10 and 10-A chimeric EcoRI-BglI C-terminal regions are not intrinsically defec-tive. Weconclude that the MCFN-terminal region cannot
combine with either the A-10or 10-A C-terminal region to formafunctionalgp7OSU.
The 10.M chimera exhibited a complex interference pat-tern (Table 1; Fig. 2). The MuLV containing the 10.M chimeric gp70SU had amphotropic receptor specificity, as 10.M interferedwith HaSV(ampho)and cells infected with amphotropic MuLVinterferedwith HaSV(10.M) superinfec-tion. However, in addition to this functional amphotropic receptorspecificity, 10.M interfered with Ha(Mo-MCF) and Ha(10A1). Taken together, these results indicate that the 10.M chimera has one functional receptor specificity, i.e., amphotropic; in addition, itcanbind toboth the MCF and 1OAl receptors but cannot use them to initiate infection.
DISCUSSION
Inthis study, wehaveanalyzed the receptorspecificities ofaseries ofchimeric
gp7Oss.
Thechimerasweredesigned to separate two variable regions of gp7OSU in order to determine which, if either, of these regions determines receptorspecificity.Ourresults, summarizedinFig. 2, revealed the existence ofahighly complex series of interactionsbetweendifferent domains ofgp7O. Thus,inonlytwoofthesix chimerascould receptorspecificity be attributed simply to a single region of the molecule. The properties of the chimeras and their implications for theorigin of the different receptor specific-ities arediscussed below.
Twochimeraswerefoundtobe abletoinfect cells viaone receptorbut to also interfere with infection at one or more additional receptors. One way toexplain thisobservation is to postulate a second function for gp7OSu, in addition to receptorbinding. Accordingtothis hypothesis, these chime-ras would be unable to carry out this second function after binding the receptor in question. This idea is analogous to thatusedtoexplainsimilar results with TM protein mutants by Delwart andPanganiban (4)andGranowitz et al. (10): in the case of these mutants, it seems likely thatgp70canbind to the receptor but that the mutant TM is defective. An alternatehypothesisassumesthat ininterference with super-infection, the receptor is prevented from reaching the cell surface asaresultofanencounterwith a newly synthesized Envmolecule within the cell. According to this hypothesis, one orbothof these molecules is in an immature state at the time of this encounter. The chimeric Env would then be capable of interacting with the receptor intracellularly, al-though the mature chimeric Env on a virion would be incapable of binding the corresponding maturereceptor at the cell surface.
Determinants for amphotropic receptor specificity. Our analysis shows clearly that most of the chimeras have amphotropic receptor specificity, since three of the five functional chimeras(A.10,A.M, and10.M)can useonly the amphotropicreceptortoinfect NIH 3T3 cells(Fig. 2). Both chimeric Envs containing the amino half of amphotropic
gp7OSu
had amphotropicreceptorspecificity.Therefore the amphotropic receptor specificity apparently maps to se-quences within the amino half ofgp7OSU.
Thisregion con-tainstwovariableloopsof sequence whichareeither absent or different in the xenotropic and MCF-polytropicgp7OSu
sequences (15); further studies are required to define the participationof these sequences in receptorspecificity.Both A.M and 10.M have additional attributes which will be discussed later.Determinants for
1OAl
receptor specificity. The interfer-ence properties of1OAl (18) suggest that its Env is able toon November 9, 2019 by guest
http://jvi.asm.org/
gp7OSU
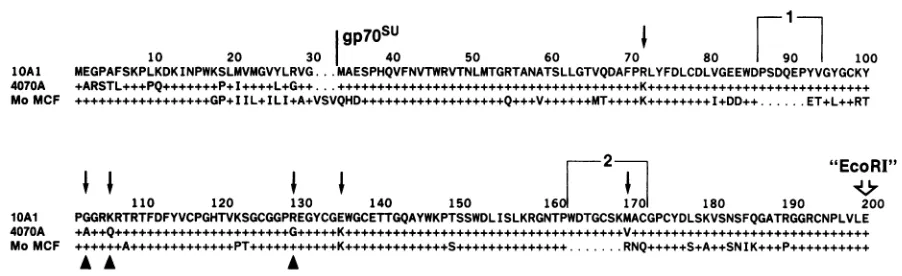
10 20 30 40 50 60 i
MEGPAFSKPLKDKINPWKSLMVMGVYLRVG...MAESPHQVFNVTWRVTNLMTGRTANATSLLGTVQDAF
+ARSTL+++PQ+++++++P+I++++L..G++...
- - - -s- -f- - - --t.D T TI T I T- %CVI n- - -BMn--s-- ----f-s---A --Is---STUr
-+++++++IL+ILI+A+VbVUrM.1L+1LR1 VV.U...Q. .V...+++ I....K...I+DD++
1
70 80 90 100
FPRLYFDLCDLVGEEWDPSDQEPYVGYGCKY
ET+L++RT
lOAl 4070A Mo MCF
2 "EcoRI"
110 120 130 140 150 160 170 180 190 200
PGGRKRTRTFDFYVCPGHTVKSGCGGPREGYCGEWGCETTGQAYWKPTSSWDLISLKRGNTPWDTGCSKMACGPCYDLSKVSNSFQGATRGGRCNPLVLE
,A--^fi.. . .sfi|X. . . . .. Lfiks.t... ...
+4
RNO
.O++++S+A++SNIK.P+.+.+.+...
FIG. 4. Proteinsequencecomparison of theamino-halves ofgp7OSus.1OAl (15), amphotropic 4070A (15), and Mo-MCF (1)sequences are
aligned. Amphotropic and Mo-MCFsequenceidentitieswith 1OAlaredenotedby+,whilesequencedifferencesaredisplayed by single-letter
amino acid code.Periods indicatespacesinsertedformaximizing alignment. Thestartofmaturegp7oSUisindicated withabar,and thetwo
amphotropic insertions areindicated by numbered brackets. Arrowshighlightsequencedifferencesbetween1OAl andamphotropic 4070A MuLV; triangles indicate thesequencedifferences from4070A that aresharedby 1OAl and Mo-MCF.
recognize boththe uniqueIOAlreceptorandthe amphotro-picreceptor. Inourpresentanalysis,wehave notidentified
anychimeraswhich interact with the 1OAlreceptorbutnot
theamphotropicreceptor.However, the resultsshowedthat 1OAl receptor specificity can apparently originate in two
distinct ways.
First, in the chimeras between amphotropic and 1OAl
sequences, 1OAl receptor specificity is clearly determined by the N-terminal portion of 1OAl
gp7oSu
in the 10.A chimera (Fig. 2). Our previous sequence analysis (15) has shown1OAlMuLVtobeaninvivo recombinantcontainingmostly amphotropicenv sequences exceptfor a
hypervari-ableregion (in the C-terminal region of 1OAl gp7O )
con-sisting of endogenous MCF-related polytropic env se-quences. This observation led us to propose that 1OAl
receptor specificity may arise by a combinatorial
mecha-nism, i.e., as a result of interactions betweentwo different regions, neither ofwhich is fromavirus with1OAlreceptor
specificity (15). It is clear that this is notthe case, as 10.A displays 1OAl receptor specificitywhereas A.10 does not.
Rather, in comparing 1OAl and amphotropic sequences of themature
gp7OSu
moleculeuptotheEcoRI restriction site (aminoacid200)usedtojointhechimeras,wefindsix amino acid differences (Fig. 4). Since 10.A recognizes the 1OAlreceptorwhereas amphotropic MuLV(A.A) does not, one or more of these six amino acid differences is clearly responsible for1OAlreceptorspecificityinthiscombination. Wearecurrently examining thecontributions of these amino
acidsto 1OAlreceptorspecificity bysite-directed
mutagen-esis.
While thismapping of 1OAl receptorspecificity israther straightforward, another chimera that displays this same
specificityismore difficult to interpret. The M.A combina-tion is made up of Mo-MCF N-terminal and amphotropic
C-terminal sequences; neither of these MuLVs can usethe
1OAl receptor, yetwhen these components arejoined
to-gether, the resultingchimera displays 1OAl receptor speci-ficity(Fig. 2).Toourknowledge,thisis the first clearcaseof acombinatorial interactionbetweentwoportions of gp7OSU thatproducesathird, distinct receptorspecificity.
It is interesting to compare these results with previous
observations on avian retroviruses. Like MuLVs, these
virusesarepolymorphic withrespect toreceptorspecificity, and their gp85Su molecules have discrete regions of se-quencevariation(2). Chimeras between different avian
sub-groupswereextensivelyinvestigated byDorneretal.(6, 7).
They found that certain
gp85"U
chimeras exhibited the receptorspecificity of both parental viruses. This additivereceptorspecificityshould bedistinguishedfrom the combi-natorialorinteractivereceptorspecificity thatweobserved
with M.AMuLV.
What is responsible for the 1OAl receptor specificity of M.A? As described above, M.A and 10.A clearly exhibit 1OAlreceptorspecificity whereas amphotropicMuLV(A.A) doesnot. One inference fromthese datamightbe thatboth MCF and 1OAl N termini can determine 1OAl receptor
specificity. Therefore, thepresenceofcommonamino acids
in MCF and 1OAl N termini that aredifferent from thosein
theamphotropicN-terminus sequenceareprovocative
can-didates for 1OAl receptorspecificity. Inthis 170-amino-acid region of mature
gp7OSu,
there are three amino acids incommon between 1OAl and Mo-MCF that are not shared
with amphotropic MuLV (Fig. 4). Thus, it is possible that
one or moreofthese threeamino acid differences determines
1OAl receptor specificity whencombined with the ampho-tropic, but notwith the MCF, carboxyhalf ofgp7OSU. The mutagenesis experiments outlined above shouldclarifythis possibilityalso.
DeterminantsforMCF receptorspecificity.Themost
com-plexinteractions that weobservedwere those whichmight determine MCFreceptor specificity. Consideration of three facts seems relevant to an understanding of these
interac-tions.
First,it is notablethatnochimerawasabletoinfect cells by usingtheMCFreceptor.Thisfindingimpliesthat bothN andC termini of the MCFgp7OSUcontributetoitsspecificity for theMCFreceptor, i.e., thatMCFreceptorspecificityis generated byinteraction between these domains.
Second, both of the chimeras containing the MCF C terminus,i.e.,A.M and10.M,areapparentlyabletobind the MCF receptor, although they are unable to use it for
infection. This observation suggeststhat theMCF C
termi-nuscontains information for MCFreceptorbinding. Third, itwasnoted above that thepropertiesof the 10.A
chimera suggest that specificity for the 1OAl receptor is carried in the1OAlNterminus.However,the10.M chimera, while it is able tobindthe 1OAl receptor, cannotuse it for infection. This difference between 10.A and 10.Msuggests thepossibilitythat theMCF C terminus contains sequences
that can mask or suppress functional 1OAl receptor speci-ficityin 10.M.
As notedabove, the1OAl receptorspecificityofthe M.A
lOAl
4070A
Mo MCF -- -- - ---uT. r%%
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.82.533.75.213.2]chimera (resembling that of the 10.Achimera) suggests that the MCF N terminus may actuallyparticipate in binding to
the 1OAl receptor. However, in this case, why. does the MCFEnv (M.M) itselfnot possess10AI receptor specific-ity?Onepossibility is thatthe MCF C terminus masks1OAl specificity in this case, as postulated above to explain the properties of10.M.
Defectiveness oftheM.10 chimera. The only chimera that didnot form afunctional env gene was the M.10 combina-tion. M.10 MuLV cannot infect mouse, CHO, rat, or mink cells. Since the M.10 Env is incorporated into virions (Fig. 3), the M.10 envelope is presumably on the plasma mem-brane of the transfected CHO cells bearing this chimeric MuLV. This M.10
gp7O5s
is apparently nonfunctional with respect to receptor binding as well as receptor utilization, since NIH 3T3 cells transfected with M.10 show no super-infection interference. Thus, M.10 appears to be defective in gp70SU-mediated receptor binding. (Of course, the M.10 chimera also contains two other gene fusions: between Mo-MuLVand Mo-MCF at the Sall site in the coding region forreversetranscriptase, andbetween1OAl and Mo-MuLVattheClaI site in the TM gene. However, these fusions yield fully functional products and are not responsible for the defectiveness of M.10, since they are also present in the infectious genomes M.A and A.10, respectively.)
It was surprising that the M.10 chimera was defective whereas the M.A chimera was not. This difference must reflect a difference between the 1OAl and amphotropic carboxy halves. We attempted to localize the 1OAl se-quencesresponsible for the defectiveness of M.10 by recip-rocally exchanging portions of amphotropic and
IOAl
car-boxy halves (Fig. 1); however, neither of the resulting chimeras (M.10-A and M.A-10) was functional (data not shown). Thus, onepossibility is that both the hypervariable regionandC-terminal region of 1OAl Env contain sequences which are independently incompatible with the Mo-MCF N-terminal sequences. However, while incompatibilitybe-tween these sequences is one explanation, there is another
possible
mechanism for the lack of M.10 gp7OSU function. From our chimeric results, it seems possible that the Mo-MCF amino half, unlike the 1OAl or amphotropic amino half, does notcontain determinants for receptor binding. In that case, itmightbethat the 1OAl C-terminal region (unlike the amphotropic C-terminal region) does not have enough information to interact with the N-terminal determinants of MCFgp7OSu
andbind a receptor.Conclusions. Our analysis of receptor specificity using chimeric envs has revealed a complex pattern of receptor
specificities,
with some unexpected results (Fig. 2). Theycan be briefly summarized as follows. The amphotropic amino half of
gp7Ou
is alwaysassociated with amphotropic receptor specificity and appears to be responsible for this receptor specificity. The 1OAl amino half is also correlated with 1OAl receptor specificity; however, when it is com-bined with the Mo-MCF carboxy half, the resulting 10.M chimera canonlybind to, not infect via, the 1OAl receptor. The Mo-MCFamino half ofgp7OSu wasnotclearly associ-ated with a receptor specificity. The carboxy halves ofgp7Oss
did not have a clear association with a receptorspecificity
exceptfor the Mo-MCF carboxy half ofgp7OsU.
Thisregionof Mo-MCF isconsistently associated with MCF receptorbinding, though it requires the corresponding Mo-MCF N-terminalportion for full function in infection. From these observations, it appears that receptor specificity can be formed in twoways: by determinants in the N-terminalportion
ofgp7OSU
orbycombinatorial interactionsbetweentwo regions. Further studies are in progress to better de-scribe PRM receptorspecificity.
After this workwas submitted for publication, Battini et al. (la) published areport describingproperties ofa similar setof chimeras. Theresults of the twostudies are generally consistent except thatthe chimera corresponding to our M.A chimera was defective in the experiments of Battini et al., whereasM.A was fully infectiousin our hands. Onepossible explanation for this discrepancy is that different MCF iso-lates were used as parentsofthe chimeras constructed in the two laboratories.
ACKNOWLEDGMENTS
We thank Marguerite Vogt for the Mo-MCF clone; Donald Blair, James Cunningham, Hung Fan, and Nancy Rice for helpful discus-sions and comments on the manuscript; and Carol Shawver for preparation of the manuscript.
This research was sponsored by the National Cancer Institute under contract N01-CO-74101 with ABL.
REFERENCES
la.Battini, J.-L., J. M. Heard, and0. Danos. 1992. Receptor choice determinants in the envelope glycoproteins of amphotropic, xenotropic, and polytropic murine leukemia viruses. J. Virol. 66:1468-1475.
1. Bosselman, R. A., F. van Straaten, C. Van Beveren,
I.
M. Verma, and M. Vogt. 1982. Analysis of the env gene of molec-ularly cloned and biologically active Moloney mink cell focus-forming proviral DNA. J. Virol. 44:19-31.2. Bova, C. A., J. P. Manfredi, and R. Swanstrom. 1986. env genes of avian retroviruses: nucleotide sequence and molecular re-combinants define host range determinants. Virology 152:343-354.
3. Chattopadhyay, S. K., A. I. Oliff, D. L. Linemeyer, M. R. Lander, and D. R. Lowy. 1981. Genomes of murine leukemia viruses isolated from wild mice. J. Virol. 39:777-791.
4. Delwart, E. L., and A. T. Panganiban. 1989. Role of reticuloen-dothelosis virus envelope glycoprotein in superinfection inter-ference. J. Virol. 63:273-280.
5. Dickson, C., R. Eisenman, H. Fan, E. Hunter, and N. Teich. 1982. Protein biosynthesis and assembly, p. 513-648. In R. A. Weiss, N. M. Teich, H. E. Varmus, and J. M. Coffin (ed.), Molecular biology of tumor viruses: RNA tumor viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. 6. Dorner, A. J., and J. M. Coffin. 1986. Determinants for receptor
interaction and cell killing on the avian retrovirus glycoprotein gp85. Cell 45:365-374.
7. Dorner, A. J., J. P. Stoye, and J. M. Coffin. 1985. Molecular basis of host range variation in avian retroviruses. J. Virol. 53:32-39.
8. Gorelick, R. J., L. E. Henderson, J. P. Hanser, and A. Rein. 1988. Point mutants of Moloney murine leukemia virus that fail to package viral RNA: evidence for specific RNA recognition by a "zinc-finger-like" protein sequence. Proc. Natl. Acad. Sci. USA85:8420-8424.
9. Graham, F. L., and P. J. van der Eb. 1973. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 52:456-471.
10. Granowitz, C., J. Colicelli, and S. P. Goff. 1991. Analysis of mutations in the envelope gene of Moloney murine leukemia virus: separation of infectivity from superinfection resistance. Virology 183:545-554.
11. Heard, J. M., and0. Danos. 1991. An amino-terminal fragment of the Friend murine leukemia virus envelope glycoprotein binds the ecotropic receptor. J. Virol. 65:4026-4032.
12. Koch, W., G. Hunsmann, and R. Friedrich. 1983. Nucleotide sequence of the envelope gene of Friend murine leukemia virus. J. Virol. 45:1-9.
13. Koch, W., W. Zimmerman, A. Oliff, and R. Friedrich. 1984. Molecular analysis of the envelope gene and long terminal repeat of Friend mink cell focus-inducing virus: implications for
on November 9, 2019 by guest
http://jvi.asm.org/
the functions of these sequences.J. Virol. 49:828-840. 14. Ott, D. (NCI-Frederick CancerResearch and Development
Cen-ter).Unpublished data.
15. Ott, D., R. Friedrich, and A. Rein. 1990. Sequence analysis of amphotropic and 1OAl murine leukemia viruses: close relation-shiptomink cellfocus-inducing viruses. J. Virol. 64:757-766. 16. Pinter, A., and W. J. Honnen. 1984. Characterization of
struc-tural and immunological properties of specific domains of Friend ecotropic and dualtropic murine leukemia virus gp70s. J. Virol.49:452-458.
17. Rein, A. 1982. Interference grouping of murine leukemia
vi-ruses: adistinctreceptor for the MCF-recombinant viruseson
mousecells. Virology 120:251-257.
18. Rein,A., andA. Schultz. 1984. Different recombinant murine
leukemia virusesuse different cell surfacereceptors.Virology 136:144-152.
19. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981. NucleotidesequenceofMoloney murine leukemia virus. Nature (London) 293:543-548.
20. Steck, F. T., andH.Rubin. 1966. The mechanism of interference between an avian leukosis virus and Rous sarcomavirus. II.
Earlysteps of infection by RSV under conditions of
interfer-ence.Virology 29:642-653.
21. Versteegen, R. J., and S. Oroszlan. 1980. Effect of chemical
modification and fragmentation on antigenic determinants of
internal protein p30 and surface glycoprotein gp7O oftype C retroviruses. J. Virol. 33:983-992.