0022-538X/06/$08.00⫹0 doi:10.1128/JVI.80.10.4648–4655.2006
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
A Tryptophan-Rich Motif in the Carboxyl Terminus of the Small
Envelope Protein of Hepatitis B Virus Is Central to the
Assembly of Hepatitis Delta Virus Particles
Isabelle Komla-Soukha
1and Camille Sureau
1,2*
Laboratoire de Virologie, INTS, Paris, France 75739,1and Department of Virology and Immunology,
Southwest Foundation for Biomedical Research, San Antonio, Texas 782282
Received 29 December 2005/Accepted 20 February 2006
The small hepatitis B virus surface antigen (S-HBsAg) is capable of driving the assembly and secretion of hepatitis delta virus (HDV) particles by interacting with the HDV ribonucleoprotein (RNP). Previously, a specific domain of the S-HBsAg protein carboxyl terminus, including a tryptophan residue at position 196 (W196), was proven essential for HDV maturation (S. Jenna and C. Sureau, J. Virol. 73: 3351–3358, 1999). Mutation of W196 to phenylalanine (W196F) was permissive for HBV subviral particle (SVP) secretion but deleterious to HDV virion assembly. Here, the W196F S-HBsAg deficiency was assigned to a loss of its ability for interaction with the large HDV antigen (L-HDAg), a major component of the RNP. Because the overall S-HBsAg carboxyl terminus is particularly rich in tryptophan, an amino acid frequently involved in protein-protein interactions, site-directed mutagenesis was conducted to investigate the function of the S-HBsAg Trp-rich domain in HDV assembly. Single substitutions of tryptophan between positions 163 and 201 with alanine or phenylalanine were tolerated for SVP secretion, but those affecting W196, W199, and W201 were detrimental for HDV assembly. This was proven to result from a reduced capacity of the mutants for interaction with L-HDAg. In addition, a W196S S-HBsAg mutant, which has been described in HBV strains that arose in a few cases of lamivudine-treated HBV-infected patients, was deficient for HDV assembly as a consequence of its impaired capacity for interacting with L-HDAg. Interestingly, the fact that even the most conservative substitution of phenylalanine for tryptophan at positions 196, 199, or 201 was sufficient to ablate interaction of S-HBsAg with L-HDAg suggests that W196, W199, and W201 are located at a binding interface that is central to HDV maturation.
The hepatitis delta virus (HDV) particle consists of an outer envelope provided by the helper hepatitis B virus (HBV) and an inner ribonucleoprotein (RNP). The RNP is made of a circular single-stranded RNA genome and two related HDV-encoded proteins that bear the hepatitis delta antigen (HDAg) and are referred to as the small and large HDAg proteins (S-HDAg and L-HDAg, respectively) (9). S-HDAg is 195 amino acid residues in length, and L-HDAg is longer by 19 amino acids at its carboxyl terminus. The difference in size arises as a consequence of an RNA editing event that occurs during replication of the viral genome (31). The HDV enve-lope includes cell-derived lipids and the HBV enveenve-lope pro-teins (large, middle, and small) bearing the hepatitis B surface antigen (L-HBsAg, M-HBsAg, and S-HBsAg, respectively) (8). In the HBV replication cycle, S-HBsAg is the major com-ponent of the virion envelope, but its massive production in an infected cell leads essentially to its secretion as empty subviral particles (SVPs) through a nucleocapsid-independent mecha-nism (11). Mature HBV virions are assembled owing to the S-HBsAg activity for driving the budding of particles at the endoplasmic reticulum (ER) membrane and to the matrix function of the L-HBsAg for recruiting the HBV nucleocapsid in the budding particles (2). But as a consequence of its
pro-pensity for self-assembly, S-HBsAg alone can also package the HDV RNP, leading to the formation of particles that are structurally identical to mature HDV virions (42). However, the presence of L-HBsAg, along with S-HBsAg, in the enve-lope, is required to confer infectivity (37). The L-HDAg pro-tein is the RNP component that mediates the interaction with S-HBsAg during HDV assembly. Interestingly, when coexpressed with the HBV envelope proteins, in the absence of HDV RNA and S-HDAg, L-HDAg is packaged and secreted in the HBV SVPs (3).
The S-HBsAg sequence contains all the information neces-sary for its own secretion and that of HDV. S-HBsAg proteins are synthesized at the ER membrane as glycosylated (gp27) and nonglycosylated (p24) forms; they are thought to be trans-ported via vesicles toward the Golgi compartment and to be secreted as empty 20-nm-diameter particles after oligomeriza-tion and budding at the pre-Golgi membrane into the lumen (12, 28). At the ER membrane (see Fig. 2B), the topology of S-HBsAg has been partially elucidated: the amino terminus (residues 1 to 3) is exposed at the luminal side of the mem-brane; it is followed by a first transmembrane domain (TMD-I) between residues 4 and 28, a cytosolic loop between residues 28 and 80, and a second TMD (TMD-II) that anchors the polypeptide chain into the membrane in the opposite direction with respect to TMD-I. The region located between residues 100 and 164, also referred to as the antigenic loop (AGL), contains the major antigenic epitopes and a glycosylation site at Asn-146. The AGL is translocated into the ER lumen
* Corresponding author. Mailing address: Laboratoire de Virologie, Institut National de la Transfusion Sanguine, 6 rue Alexandre Cabanel, 75739, Paris, France. Phone: 33 1 44 49 30 56. Fax: 33 1 44 49 30 56. E-mail: csureau@ints.fr.
4648
on November 8, 2019 by guest
http://jvi.asm.org/
during synthesis, leading to its exposure at the surface of secreted particles. The topology of the carboxyl-terminal domain, between residues 164 and 226, is not precisely known, but the sequence is highly hydrophobic and pre-dicted to contain two transmembrane ␣-helical structures (TMD-III and TMD-IV) (30).
How the S-HBsAg manages to package the HDV RNP is still uncertain. But recent studies have identified discrete re-gions, or amino acid residues of S-HBsAg, which are important for HDV assembly and/or secretion and, yet, dispensable for SVP secretion. Chen et al. (6) reported that a truncation of the last 50 residues was sufficient to abolish envelopment and se-cretion of coexpressed HDAg proteins. In a study conducted by O’Malley and Lazinski (26), S-HBsAg bearing a deletion in the AGL (from residues 107 to 147) was proven defective for HDV RNP envelopment. Interestingly, this mutant remained competent for the incorporation in the SVPs of singly ex-pressed L-HDAg, suggesting that the hindrance observed in HDV maturation was probably reflecting a reduced capacity of the envelope to accommodate an RNP. However, the defi-ciency of S-HBsAg with the AGL deleted could also be ex-plained by its lack of N-linked carbohydrates, since the re-moval of the glycosylation site (Asn-146) is sufficient to impair HDV assembly (36).
In addition to the glycosylation site, we previously identified two determinants of HDV maturation: (i) a sequence encom-passing residues 24 to 28 at the cytosolic interface of TMD-I (16) and (ii) a sequence between residues 179 and 203 at the carboxyl terminus of S-HBsAg (17). In the latter determinant, mutation of a single amino acid (Trp-196) led to a greater than 50-fold reduction of the mutant’s capacity for HDV assembly without affecting SVP secretion. Surprisingly, this phenotype was observed with the most conservative substitution, W196F. This result pointed to the crucial role of the tryptophan side chain in HDV RNP interaction. However, the ability of such a mutant for interacting with L-HDAg was not directly eval-uated.
Since the carboxyl terminus of S-HBsAg (residues 163 to 226) comprises nine tryptophan residues that are conserved among all HBV genotypes, single point substitutions by alanine or phenylalanine were made to assess the function of the Trp-rich domain in HDV assembly. We report that none of the Trp substitutions had a significant effect on HBV SVP secretion, but the replacement of Trp-196, -199, and -201 with alanine or phenylalanine was detrimental to interaction of S-HBsAg with L-HDAg and, consequently, to assembly of HDV particles. The results indicate that tryptophan side chains of the short peptide sequence located between the two predicted TMDs, III and IV, are critical to HDV assembly.
MATERIALS AND METHODS
Construction of plasmids.The parental HBV recombinant plasmid (p123) contains the envelope protein-coding region of the genotype D HBV DNA (36). For construction of plasmid pCIHBS, a DNA fragment (nucleotides 129 to 837 of the ayw3 HBV genome) encoding the S-HBsAg protein was amplified by PCR
using 5⬘and 3⬘primers tagged with the XhoI and NotI restriction site sequences,
respectively. After digestion with XhoI and NotI restriction endonucleases, the DNA fragment was inserted between the XhoI and NotI restriction sites of the
pCI vector (Promega). Plasmids for the expression of⌬24-28KL, N146T, and
W196F S-HBsAg mutants have been generated by subcloning into pCI an XhoI
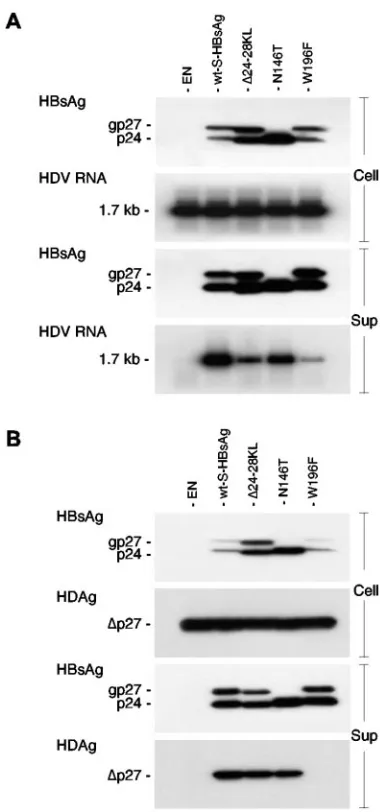
-FIG. 1. Analysis of⌬24-28KL, N146T, and W196F S-HBsAg mu-tants for their capacity for HDV assembly (A) and interaction with L-HDAg (B). Immunoblot analysis of S-HBsAg or L-HDAg proteins from culture supernatant (Sup) or cell lysate (Cell) was performed after transfection of HuH-7 cells with a mixture of 1g of HDV recombinant plasmid pSVLD3 (A) or pCIHD27 (B) and 1g of wt or mutant S-HBsAg plasmid DNA. Culture medium was harvested at day 2 (B) or days 4 to 10 (A) posttransfection, and particles were subjected to sedimentation by ultracentrifugation. Cells were harvested at day 2 posttransfection (A and B) and lysed in protein disruption buffer. Proteins from sedimented particles (equivalent to 1 ml of supernatant) or cell lysates (equivalent to 105 cells) were separated on a 15%
acrylamide gel, transferred to a PVDF membrane, and probed with anti-HBsAg or anti-HDAg antibodies (1:1,000). For Northern blot analysis of HDV RNA, particles from culture medium (Sup) harvested at days 4 to 10 posttransfection were subjected to sedimentation, and RNA was purified using the QIAmp viral RNA extraction kit (QIAGEN). Cells were harvested at day 10 posttransfection for purification of total cellular RNA using the RNeasy Mini kit (QIAGEN). RNA extracted from 1 ml of culture supernatant or 5g of total RNA was separated on an agarose gel, transferred to nylon membrane, and hybridized to a genomic strand-specific 32P-labeled HDV RNA probe. The size in
kilobases of HDV genomic RNA is indicated. The unglycosylated and glycosylated S-HBsAg (p24 and gp27, respectively) and L-HDAg (⌬ p27) are indicated at the left side of each panel. EN, HBVenv-negative plasmid.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:2.585.324.514.75.481.2]NotI DNA fragment that was amplified by PCR from the respective parental plasmids described previously (16, 17, 36).
The parental HDV recombinant plasmid (pSVLD3) contains three head-to-tail copies of the full-length HDV cDNA (19). For construction of pCIHD24, a DNA fragment (nucleotides 1598 to 954 of the genotype 1 HDV cDNA)
encod-ing the S-HDAg protein was amplified by PCR from pSVLD3 usencod-ing 5⬘and 3⬘
primers tagged with the NheI and NotI restriction site sequences, respectively. After digestion with NheI and NotI, the DNA fragment was inserted between the NheI and NotI sites of the pCI vector. Plasmid pCIHD27 was generated from pCIHD24 using the PCR mutagenesis protocol to convert the TAG stop codon of the S-HDAg gene into a TGG tryptophan codon for expression of the L-HDAg protein.
Mutagenesis was carried out using the PCR overlap extension method on the pCIHBS or pCIHD27 plasmids, with two complementary mutagenic oligonucle-otides as described elsewhere (16). The mutations were designated by the one-letter code of the wild-type (wt) amino acid followed by its position in S-HBsAg and the one-letter code for the substituted amino acid. All PCR-generated fragments were cloned in the pCI vector (Promega), and the resulting mutant plasmids were verified using the Big Dye Terminator sequencing protocol (Ap-plied Biosystems). Clones containing the desired mutations were selected and used for subsequent transfections. Plasmid pSVLD3 was used for expression of HDV RNPs, and plasmids pCIHBS, pCIHD24, and pCIHD27 were used for expression of S-HBsAg, S-HDAg, and L-HDAg, respectively.
Transfection of HuH-7 cells.HuH-7 cells were maintained in Williams’ me-dium E supplemented with 10% fetal bovine serum. For production of HDV particles, cells were transfected with a mixture of HDV recombinant plasmid pSVLD3 for the synthesis of HDV RNPs and a wt or mutant pCIHBS for the supply of the wt or mutant S-HBsAg, respectively. For coexpression of L-HDAg and S-HBsAg, cells were cotransfected with a mixture of wt or mutant pCIHD27 and wt or mutant pCIHBS plasmids.
Transfection of 8⫻105cells was carried out using the FuGENE-6 reagent
(Roche) according to the instructions of the manufacturer, with 1g of HDV
recombinant plasmid (pSVLD3 or pCIHD27) and 1 or 2g of pCIHBS or
derivatives. Culture medium was harvested on days 2, 4, 6, 8, and 10 posttrans-fection and analyzed for the presence of viral particles by immunoblotting for detection of S-HBsAg or L-HDAg proteins and by Northern blotting for detec-tion of HDV RNA. For analysis of cellular RNA or proteins, cells were harvested 9 days after cotransfection with pSVLD3 and pCIHBS or derivatives or 2 days after cotransfection with pCIHD27 and pCIHBS or their respective derivatives.
Characterization of HDV particles produced in HuH-7 cells.Culture fluids harvested on days 2 (or 3), 6, and 9 after transfection were clarified by
centrif-ugation at 5,000⫻gat 4°C for 30 min. Viral particles from the clarified medium
were subjected to sedimentation by centrifugation for 2 h at 50,000 rpm in an
SW50 rotor (Beckman) on 1 ml of a 20% sucrose cushion in 1⫻
phosphate-buffered saline, pH 7.4. After centrifugation, the particle-containing pellet was resuspended in 10 mM Tris-HCl (pH 7.4) containing 150 mM NaCl, 1 mM EDTA. Disruption buffer was added, and proteins were submitted to sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transfer to polyvinylidene difluoride (PVDF) membrane, and immunodetection with rabbit anti-HBsAg
antibodies (R247) or human anti-HDAg antibodies (36). Immunoblots were developed by chemiluminescence (ECL kit; Amersham) and exposed to Kodak films for detection of light emission.
For HDV RNA analysis, viral particles from the clarified medium were centrated by centrifugation as described above. RNA extraction from the con-centrated particles or from cells harvested at day 9 posttransfection was carried out as described previously (15). Detection of viral or cellular HDV RNA was achieved after electrophoresis through a 1.2% agarose–2.2 M formaldehyde gel,
transfer to nylon membrane (Roche), and hybridization to a32P-labeled
HDV-specific RNA probe (15).
RESULTS
The failure of W196F S-HBsAg to assist in HDV assembly
results from a lack of interaction with L-HDAg.Previously, we
showed that a deletion of residues 24 to 28 of the S-HBsAg protein (16), the removal of the glycosylation site at position 146 (36) or, more importantly, the substitution of Trp-196 with phenylalanine (17) was detrimental to the maturation of HDV particles while having no effect on SVP secretion (Fig. 1A). To investigate whether this phenomenon was due to a lack of interaction between L-HDAg and S-HBsAg, HuH-7 cells were transfected with an L-HDAg expression plasmid and a vector coding for the ⌬24-28, N146T, or W196F S-HBsAg mutant. Transfected cells and culture fluids were harvested at day 2 posttransfection and analyzed for the presence of S-HBsAg and L-HDAg by immunoblotting. Control experiments con-sisted of the transfection of HuH-7 cells with an L-HDAg expression vector in the absence (Fig. 1, EN) or presence of a wt S-HBsAg protein expression vector. As illustrated in Fig. 1B, the⌬24-28 and N146T mutants were competent for incorpo-ration and cosecretion of L-HDAg proteins in SVPs, whereas the W196F mutant appeared to have lost L-HDAg binding activity. Therefore, the defect of the ⌬24-28 and N146T mutants for HDV particle maturation that was previously reported (16, 36) was due not to a lack of interaction with L-HDAg but, rather, to an impairment of the SVP envelope to accommodate an RNP. In contrast, the deficiency of the W196F mutant in HDV maturation is clearly the result of a lack of interaction with L-HDAg.
Tryptophan residues at position 196, 199, and 201 of
S-HBsAg are instrumental in HDV assembly.Because our
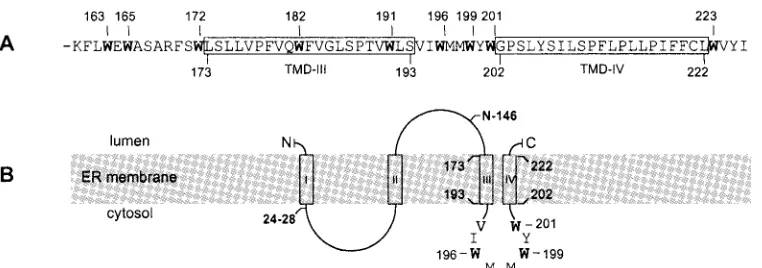
pre-FIG. 2. Schematic representation of the S-HBsAg carboxyl terminus. (A) Amino acid sequence of the carboxyl terminus of S-HBsAg. Numbers above the sequence correspond to the positions of the tryptophan residues (indicated in bold). Numbers below the sequence indicate boundaries of the predicted transmembrane domains. Open boxes represent predicted hydrophobic transmembrane domains (TMD-III and TMD-IV). (B) Schematic representation of S-HBsAg topology at the ER membrane. Open boxes represent experimentally defined (I and II) or predicted (III and IV) TMDs. Residues 24 to 28, Asn-146 (N-linked glycosylation site), and Trp-196, -199, and -201, identified as determinants of HDV maturation, are indicated in bold.
on November 8, 2019 by guest
http://jvi.asm.org/
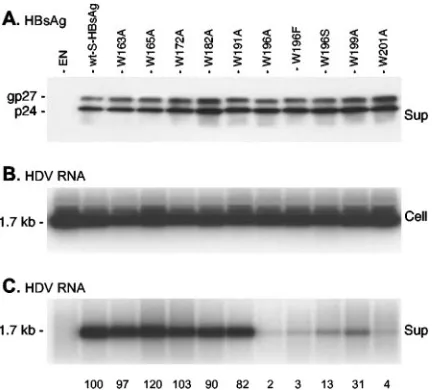
[image:3.585.105.486.67.201.2]vious study pointed to the important role of Trp-196 in HDV maturation and because the carboxyl-terminal moiety of S-HBsAg is particularly rich in tryptophan, an amino acid often engaged in protein-protein interactions, mutagenesis was con-ducted to substitute an alanine for each tryptophan between positions 163 and 201 (Fig. 2). A serine residue was also in-troduced in place of W196, because this mutant (W196S) had been described in HBV strains that occasionally emerge in lamivudine-treated HBV carriers (40). Trp-223 was not in-cluded in this analysis because we had previously shown that an S-HBsAg protein truncated at position 222 had wt charac-teristics with regard to HDV particle assembly (17). Plas-mids for expression of W163A, W165A, W172A, W182A, W191A, W196A, W199A, and W201A mutants were generated and introduced in HuH-7 cells with pSVLD3 as described elsewhere (36). Production of HDV particles was monitored by immunoblotting for the detection of S-HBsAg proteins in the culture supernatant of transfected cells and by Northern blot-ting for the detection of HDV RNA in both cell lysates and supernatants. As illustrated in Fig. 3, when S-HBsAg was not
provided (Fig. 3, EN), there was no detectable HDV RNA in the culture medium of transfected cells. Every substitution of tryptophan with alanine was permissive for SVP secretion, but only mutants W163A, W165A, W172A, W182A, and W191A had wt characteristics for maturation of HDV, as evidenced by the presence of HDV RNA in the culture fluid. In contrast, mutants bearing a substitution at position 196, 199, or 201 were deficient for HDV assembly as demonstrated by the absence, or presence in only trace amounts, of extracellular HDV RNA. As recently reported by Vietheer et al. (41), the lamivudine treatment-associated W196S mutation severely affected HDV assembly.
[image:4.585.56.272.69.264.2]To specifically address whether the deficiency of W196A, W199A, and W201A in HDV assembly resulted from a lack of interaction with L-HDAg, HuH-7 cells were cotransfected with the respective S-HBsAg mutants and a plasmid for the expres-sion of L-HDAg. As shown in Fig. 4, single substitutions of Trp-196, -199, and -201 that were detrimental to HDV assem-bly also impaired interaction with L-HDAg as evidenced by its presence in low amounts in particles derived from the culture supernatant. We concluded that tryptophan residues at posi-tion 196, 199, and 201, which are located between the two predicted TMD-III and TMD-IV of the S-HBsAg carboxyl terminus, are essential for interacting with L-HDAg while dis-pensable for secretion of HBV SVPs. The deficiency of the
FIG. 3. Effects of tryptophan-to-alanine substitutions in the C ter-minus of the S-HBsAg envelope protein on the mutants’ capacities for HDV maturation. (A) Immunoblot analysis of S-HBsAg proteins ex-tracted from culture supernatant (Sup) harvested on days 3, 6, and 9 after transfection of HuH-7 cells with a mixture of 1g of pSVLD3 plasmid DNA and 1 g of wt or mutant S-HBsAg plasmid DNA. Particles sedimented from 1 ml of culture medium were disrupted in Laemmli sample buffer. Proteins were separated on a 15% acrylamide gel, transferred to a PVDF membrane, and probed with S anti-bodies (1:1,000). (B) Northern blot analysis of intracellular HDV RNA (Cell). Cellular RNA was extracted from HuH-7 cells harvested at day 9 posttransfection. Five micrograms of total RNA was separated on an agarose gel and analyzed for the presence HDV RNA after transfer to nylon membrane and hybridization to a genomic strand-specific32
P-labeled HDV RNA probe. (C) Northern blot analysis of HDV RNA extracted from 1 ml of cell culture supernatant (Sup) harvested on days 3, 6, and 9 after transfection. RNA was analyzed by agarose gel elec-trophoresis, transfer, and hybridization to a genomic strand-specific
32P-labeled HDV RNA probe as for panel B. Numbers at the bottom
[image:4.585.312.528.72.267.2]of panel C are the percentages of HDV RNA incorporated by wt S-HBsAg, normalized to the same amount of secreted S-HBsAg. Quantitation was performed using the NIH Image software, version 1.63. The unglycosylated and glycosylated S-HBsAg (p24 and gp27, respectively) are indicated. EN, HBVenv-negative plasmid.
FIG. 4. Effects of tryptophan-to-alanine substitutions in the C ter-minus of the S-HBsAg envelope protein on the mutants’ capacities to package L-HDAg. Immunoblot analysis was performed on viral pro-teins extracted from culture supernatant (Sup) harvested at day 2 after transfection of HuH-7 cells with a mixture of 1g of L-HDAg-ex-pressing plasmid DNA and 1g of wt or mutant S-HBsAg plasmid DNA. Particles from 1 ml of culture medium were disrupted in Laemmli sample buffer. Proteins were separated on a 15% acrylamide gel, transferred to a PVDF membrane, and probed with anti-HBsAg antibodies (A) or anti-HDAg antibodies (C). (B) Immunoblot analysis of L-HDAg pro-teins extracted from cell lysates (Cell) at day 2 posttransfection. Num-bers at the bottom of panel C are the percentage of L-HDAg incor-porated by wt S-HBsAg, normalized to the same amount of secreted S-HBsAg. Quantitation was performed using the NIH Image soft-ware, version 1.63. The unglycosylated and glycosylated S-HBsAg (p24 and gp27, respectively) and L-HDAg (⌬p27) are indicated. EN, HBV env-negative plasmid.
on November 8, 2019 by guest
http://jvi.asm.org/
lamivudine resistance-associated mutant W196S clearly results from its reduced capacity to interact with L-HDAg. Further-more, as shown in Fig. 5, a triple S-HBsAg mutant carrying alanine substitutions at positions 196, 199, and 201 is fully
competent for SVP secretion but totally inactive for HDV assembly and L-HDAg interaction.
As demonstrated in this and previous studies (17), the most conservative substitution at position 196 (W196F) was not compatible with HDV maturation or L-HDAg interaction, in-dicating that the indole ring of the tryptophan residue, and not its hydrophobic or aromatic properties, was essential for inter-action with L-HDAg. To investigate if a tryptophan side chain at positions 199 and 201 is also an absolute requirement for HDV maturation or L-HDAg interaction, phenylalanine, the closest amino acid to tryptophan, was substituted for W199 and W201, and the S-HBsAg mutants were tested as described above for HDV maturation. As shown in Fig. 5, W201F and, to a lesser extent, W199F exhibited a reduced capacity for both HDV assembly and interaction with L-HDAg. These results demonstrate that HDV assembly depends most likely on an interaction between a tryptophan-rich motif on the S-HBsAg protein, comprising W196, W199, and W201, and the L-HDAg protein, assuming that a direct binding is involved.
Mutations of tryptophan and proline residues in the
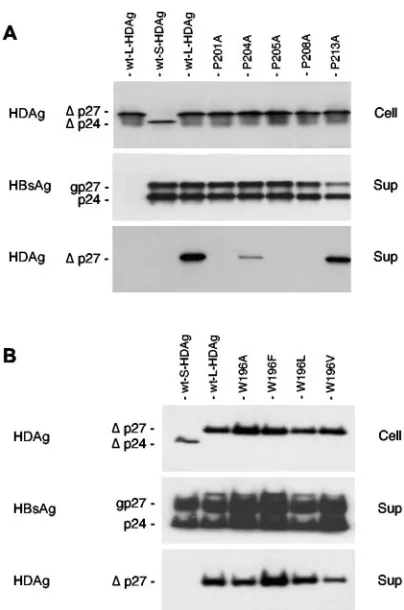
L-HDAg carboxyl terminus.With regard to L-HDAg, the
car-boxyl-terminal 19 residues are likely to constitute a packaging signal and a possible ligand to the S-HBsAg tryptophan-rich domain. This sequence has been described to include a nuclear export signal (NES) (21) and a CXXQ motif (where C is cysteine, Q is glutamine, and X is any amino acid) for farne-sylation (10). The farnesyl group covalently bound to Cys-211 is essential but not sufficient for interaction with S-HBsAg (23).
Recently, O’Malley and Lazinski (27) reported that the sub-stitution of Ala for Trp-196 of L-HDAg had no effect on packaging with S-HBsAg, whereas mutation of the proline residues at positions 201 and 205 were detrimental. Although Pro-201 and Pro-205 reside in the predicted NES, it was dem-onstrated that the lack of packaging with S-HBsAg was not due to a deficient NES in the corresponding mutants. Here we have confirmed these results (Fig. 6) by substituting alanine for proline at positions 201, 204, 205, 208, and 213 and by testing the mutants for packaging with wt S-HBsAg. Only the P213A mutant could be packaged to the wt level, whereas P201A, P205A, P208 and, to a lesser extent, P204A were deficient in S-HBsAg interaction. Experiments conducted on cell lysates and supernatants harvested on days 2 or 6 after transfection generated identical results (data not shown). In addition, by performing a separation of nuclei and cytosol from cells har-vested at day 2 posttransfection, we observed that the mutants were detected at wt level in both the nuclear and cytosolic extracts (data not shown). In agreement with the study of O’Malley and Lazinski (27), the substitution of Ala, Phe, Leu, or Val for Trp-196 in L-HDAg had no effect on the mutant’s capacity for interaction with S-HBsAg. All Trp-L-HDAg mu-tants were detected to near-wt levels in the culture medium upon coexpression with wt S-HBsAg (Fig. 6B).
DISCUSSION
Previous studies had shown that most of the small internal deletions that were introduced into the carboxyl terminus of S-HBsAg had a detrimental effect on the assembly of HDV particles (only the deletion of residues 214 to 218 was reported to exert a positive effect) (17). In the present study, we
pre-FIG. 5. Effects of tryptophan-to-phenylalanine substitutions in the C terminus of the S-HBsAg envelope protein on the mutants’ capac-ities for HDV maturation (A) or interaction with L-HDAg (B). (A) Immunoblot analysis of S-HBsAg proteins extracted from a pool of culture supernatants (Sup) harvested on days 3, 6, and 9 after transfection with a mixture of 1g of pSVLD3 plasmid DNA and 1g of wt or mutant S plasmid DNA. Particles from 1 ml of supernatant were disrupted in Laemmli sample buffer. Proteins were separated on a 15% acrylamide gel, transferred to a PVDF membrane, and probed with anti-HBsAg antibodies (1:1,000). Cellular RNA was extracted from HuH-7 cells harvested at day 9 posttransfection. Five micrograms of total RNA was separated on an agarose gel and analyzed for the presence HDV RNA after transfer to nylon membrane and hybridiza-tion to a genomic strand-specific32P-labeled HDV RNA probe (Cell).
Particles from 1 ml of culture supernatant (Sup) were analyzed for the presence of genomic HDV RNA as described above. (B) Immunoblot analysis of S-HBsAg proteins extracted from culture supernatant (Sup) harvested 3, 6, and 9 days after transfection of HuH-7 cells with a mixture of 1g of L-HDAg-expressing plasmid DNA and 1g of wt or mutant S-HBsAg plasmid DNA. Particles from 1 ml of culture medium (Sup) were disrupted in Laemmli sample buffer. Proteins were separated on a 15% acrylamide gel, transferred to a PVDF membrane, and probed with anti-HBsAg or anti-HDAg antibodies. L-HDAg pro-teins extracted from cell lysates (Cell) at day 2 posttransfection were analyzed as described above. The unglycosylated and glycosylated S-HBsAg (p24 and gp27, respectively) and L-HDAg (⌬p27) are indi-cated. EN, HBVenv-negative plasmid.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.62.265.69.394.2]cisely identified a tryptophan-rich motif comprising Trp-196, -199, and -201 as essential to interaction with L-HDAg and maturation of HDV particles.
The carboxyl-terminal end (residues 163 to 226) of S-HBsAg is highly hydrophobic and surprisingly rich in tryptophan res-idues that are strictly conserved among all orthohepadna-viruses. In contrast, only two tryptophan residues, at positions equivalent to W163 and W196 of HBsAg, are found in S-DHBsAg, the small envelope protein of theDuck hepatitis B virus(DHBV) (26). Orthohepadnaviruses such asWoodchuck hepatitis virus (WHV) and Woolly monkey hepatitis B virus
(WMHBV) can assist HDV propagation because their small envelope proteins (S-WHsAg and S-WMHBsAg, respectively) are competent for HDV RNP envelopment (1, 32, 34). In contrast, the envelope protein of theAvihepadnavirusDHBV is unable to package the HDV RNP (26). Compared to S-HBsAg or S-WHsAg, the S-DHBsAg polypeptide appears to lack the carboxyl-terminal Trp-rich domain and the region correspond-ing to the AGL between TMD-II and TMD-III (Fig. 2). When
the AGL was experimentally deleted on S-HBsAg, it led to a drastic reduction in the capacity of the mutant for HDV mat-uration (26). However, this mutant was still competent for the envelopment of the singly expressed L-HDAg protein, demon-strating the absence of an L-HDAg interaction determinant in the AGL.
The S-HBsAg carboxyl terminus is predicted to contain two TMDs (TMD-III and TMD-IV) located at positions 173 to 193 and 202 to 222, respectively (30). The Trp-196-201 cluster identified here as crucial for HDV maturation belongs to a short sequence (residues 194 to 201) that is predicted to make the junction between TMD-III and TMD-IV (Fig. 2). Accord-ing to the topological model of Persson and Argos (30), it is oriented toward the cytosolic side of the ER membrane, a position potentially adequate for interaction with L-HDAg and the RNP. However, a different topological model obtained by epitope mapping of monoclonal antibodies raised against HBV particles places the 187-207 region at the viral particle surface (7, 29). As previously discussed, the discrepancy between the two hypotheses might indicate that two topologies of the S-HBsAg carboxyl terminus coexist in the viral envelope.
The fact that Trp-196, -199, and -201 are conserved among all orthohepadnaviruses, and yet dispensable for SVP secre-tion, might indicate that their contribusecre-tion, if any, to the HBV life cycle resides in the virion assembly process or at viral entry. However, the conservation might also result from the presence of essential domains in the HBV polymerase encoded by the S-HBsAg DNA sequence in the⫺1 reading frame (38). Inter-estingly, the Trp-196 codon is included in the DNA sequence that encodes the YMDD motif of the polymerase catalytic domain (Fig. 7). This motif is crucial for the activity of the enzyme, and only in lamivudine-resistant virus is YMDD
con-FIG. 6. Effects of single amino acid substitutions in the C terminus of the L-HDAg protein on the mutants’ capacities to copackage with S-HBsAg. (A) Proline-to-alanine substitutions in L-HDAg; (B) tryp-tophan-196–to–alanine, –phenylalanine, –leucine, or –valine substitu-tions. HuH-7 cells (8⫻105) were transfected with a mixture of 1g of
[image:6.585.62.264.65.370.2]wt S-HBsAg-expressing plasmid DNA and 1g of wt S-HDAg, wt L-HDAg, or mutant L-HDAg plasmid DNA. Proteins extracted from cells (Cell) harvested at day 2 after transfection or from 1 ml of culture supernatant (Sup) harvested at day 2 posttransfection were analyzed by immunoblotting with anti-HBsAg or anti-HDAg antibodies. The unglycosylated and glycosylated S-HBsAg (p24 and gp27, respectively) and S-HDAg and L-HDAg (⌬p24 and⌬p27, respectively) are indi-cated.
FIG. 7. Sequence of the overlapping HBV polymerase and S-HBsAg genes. (A) Nucleic acid sequence coding for the 537-544 domain of wt HBV polymerase (wt-pol) and the overlapping 194-201 carboxyl-ter-minus domain of wt S-HBsAg (wt-env). (B) Nucleic acid sequence coding for a YIDD motif of lamivudine-resistant HBV polymerase and the overlapping 194-201 carboxyl-terminal domain of the correspond-ing S-HBsAg. (C) Alternative codcorrespond-ing sequences for the YMDD cata-lytic domain of wt HBV polymerase and the corresponding sequences of S-HBsAg. Note that, in order to maintain the YMDD catalytic motif of wt HBV polymerase, there is no alternative to a tryptophan residue at position 196 in S-HBsAg, Ile or Thr at position 195, Met or Thr at position 197, or Met, Thr, or Ile at position 198.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.323.517.69.232.2]verted to YVDD, YSDD, or YIDD (25, 38–40). The latter mutation creates a W196S substitution in S-HBsAg which is detrimental to HDV assembly (41). As shown in the present study, this deficiency is due to a loss of the mutant’s ability to interact with L-HDAg. Hence, one would assume that in vivo the YIDD mutation confers resistance to HDV superinfection. As depicted in Fig. 7C, the requirement for a YMDD motif in the polymerase offers no possibility, other than Trp at po-sition 196 in S-HBsAg, Ile or Thr at popo-sition 195, Met or Thr at position 197, and Met or Thr or Ile at position 198. There-fore, a few S-HBsAg residues, such as W196, might just be conserved because they share DNA coding sequence with functional domains of the polymerase, such as YMDD. It is thus tempting to speculate that HDV could recruit the HBV envelope proteins through an interaction with S-HBsAg resi-dues that are strictly conserved but nonessential to HBV, thereby lessening its interference with the helper HBV to bet-ter ensure its survival.
With regard to L-HDAg, the results of several independent studies have led to the notion that the 19 carboxyl-terminal residues, which make the difference between the L-HDAg and S-HDAg sequences, are likely to constitute a packaging signal: (i) deletions have been performed between amino acids 2 and 195 without preventing packaging with S-HBsAg (4, 5), (ii) the appending of the L-HDAg residues 196 to 214 to the carboxyl terminus of c-H-Ras (22) allowed the cosecretion of the latter with SVPs (Ras is similar in size to L-HDAg and naturally farnesylated at its carboxyl terminus, but it cannot be packaged as such by S-HBsAg), and (iii) a farnesylated S-HDAg could not be assembled with S-HBsAg, indicating that the farnesyl group, per se, is not the packaging signal (23). Surprisingly, the 19 carboxyl-terminal residues of L-HDAg are not very con-served across the different HDV genotypes, except for the CXXQ box, a tryptophan residue at position 196, and at least five proline residues diversely distributed (33). As shown here and in a previous study (17), mutation of L-HDAg Trp-196 has no effect on packaging, whereas mutation of Pro-201, -204, -205, or -208 is detrimental (27). Although the latter residues are elements of the L-HDAg NES, it was demonstrated that the lack of packaging with S-HBsAg was not due to a deficient NES in the corresponding mutants but, clearly, to a defect in S-HBsAg interaction (25).
Whether L-HDAg binds directly or not to S-HBsAg is still questionable. In favor of direct binding are the experiments conducted by Hwang and Lai (13) using a far Western blotting assay. The present study also supports this hypothesis, because it identifies a tryptophan-rich domain as the S-HBsAg deter-minant, and it confirms the results of O’Malley and Lazinski (27), which designated the carboxyl-terminal proline-rich do-main of L-HDAg as a potential ligand. Physical interaction mediated by tryptophan-rich and proline-rich motifs is a well-documented phenomenon that governs many protein-protein interactions (18, 35), but additional elements have to be con-sidered for understanding the binding potential of the Trp-196-201 cluster. First, statistical studies of sequence databases and crystal structures of integral membrane proteins show a pref-erential clustering of tryptophan residues at the membrane interface (14). It is thought to stabilize the transmembrane helix (20) and to result from the amphipathic nature of tryp-tophan that allows both hydrophobic and polar interactions
(43). Second, according to Ma et al. (24), conservation on a protein surface of Trp or, to a lesser extent, Phe and Met indicates the likelihood of their participation to a binding site. In addition, the fact that Trp-to-Phe mutations abolish inter-action with L-HDAg suggests that the Trp-rich motif belongs to, or constitutes, the L-HDAg binding site. Indeed, by replac-ing an indole rreplac-ing with a phenyl rreplac-ing, a Trp-to-Phe mutation is unlikely to affect a distal binding site because it should not create a large cavity in place of Trp in the polypeptide chain. Obviously, additional experiments need to be conducted to better understand the L-HDAg–S-HBsAg interaction. Future investigations in our laboratory will determine whether the appending of the Trp-rich domain of S-HBsAg to a heterolo-gous protein is sufficient to mediate its binding to purified L-HDAg. It would also be interesting to investigate the func-tion of the Trp-rich domain in the HBV life cycle.
ACKNOWLEDGMENTS
This work was supported by ANRS, INSERM, CNRS, and INTS. I.K.-S. was supported, in part, by the Re´gion Guadeloupe and the Conseil Ge´ne´ral de la Guadeloupe.
REFERENCES
1.Barrera, A., B. Guerra, H. Lee, and R. E. Lanford.2004. Analysis of host range phenotypes of primate hepadnaviruses by in vitro infections of
hepa-titis D virus pseudotypes. J. Virol.78:5233–5243.
2.Bruss, V.1997. A short linear sequence in the pre-S domain of the large hepatitis B virus envelope protein required for virion formation. J. Virol.
71:9350–9357.
3.Chang, F. L., P. J. Chen, S. J. Tu, C. J. Wang, and D. S. Chen.1991. The large form of hepatitis delta antigen is crucial for assembly of hepatitis delta
virus. Proc. Natl. Acad. Sci. USA88:8490–8494.
4.Chang, M. F., C. J. Chen, and S. C. Chang.1994. Mutational analysis of delta antigen: effect on assembly and replication of hepatitis delta virus. J. Virol.
68:646–653.
5.Chen, P. J., F. L. Chang, C. J. Wang, C. J. Lin, S. Y. Sung, and D. S. Chen.
1992. Functional study of hepatitis delta virus large antigen in packaging and replication inhibition: role of the amino-terminal leucine zipper. J. Virol.
66:2853–2859.
6.Chen, P. J., W. J. Lai, C. J. Wang, and D. S. Chen.1993. Hepatitis B surface antigen and large-form hepatitis delta antigen in HDV assembly: a further
study. Prog. Clin. Biol Res.382:29–34.
7.Chen, Y. C., K. Delbrook, C. Dealwis, L. Mimms, I. K. Mushahwar, and W. Mandecki.1996. Discontinuous epitopes of hepatitis B surface antigen de-rived from a filamentous phage peptide library. Proc. Natl. Acad. Sci. USA
93:1997–2001.
8.Ganem, D., and R. Schneider.2001. Hepadnaviridae: the viruses and their
replication, p. 2923–2969.InD. M. Knipe and P. M. Howley (ed.), Fields
virology. Lippincott Williams & Wilkins, Philadelphia, Pa.
9.Gerin, J. L., J. L. Casey, and R. H. Purcell.2001. Hepatitis delta virus, p.
3037–3050.InD. M. Knipe and P. M. Howley (ed.), Fields virology.
Lippin-cott Williams & Wilkins, Philadelphia, Pa.
10.Glenn, J. S., J. A. Watson, C. M. Havel, and J. M. White.1992. Identification
of a prenylation site in delta virus large antigen. Science256:1331–1333.
11.Heermann, K., and W. Gerlich.1992. Surface proteins of hepatitis B viruses.
InA. Maclachlan (ed.), Molecular biology of HBV. CRC Press, Boca Raton,
Fla.
12.Huovila, A. P., A. M. Eder, and S. D. Fuller. 1992. Hepatitis B surface
antigen assembles in a post-ER, pre-Golgi compartment. J. Cell Biol.118:
1305–1320.
13.Hwang, S. B., and M. M. Lai.1993. Isoprenylation mediates direct protein-protein interactions between hepatitis large delta antigen and hepatitis B
virus surface antigen. J. Virol.67:7659–7662.
14.Ippolito, J. A., R. S. Alexander, and D. W. Christianson.1990. Hydrogen
bond stereochemistry in protein structure and function. J. Mol. Biol.215:
457–471.
15.Jaoude, G. A., and C. Sureau.2005. Role of the antigenic loop of the hepatitis B virus envelope proteins in infectivity of hepatitis delta virus.
J. Virol.79:10460–10466.
16.Jenna, S., and C. Sureau.1998. Effect of mutations in the small envelope protein of hepatitis B virus on assembly and secretion of hepatitis delta virus.
Virology251:176–186.
17.Jenna, S., and C. Sureau.1999. Mutations in the carboxyl-terminal domain of the small hepatitis B virus envelope protein impair the assembly of
hep-atitis delta virus particles. J. Virol.73:3351–3358.
on November 8, 2019 by guest
http://jvi.asm.org/
18.Kay, B. K., M. P. Williamson, and M. Sudol.2000. The importance of being proline: the interaction of proline-rich motifs in signaling proteins with their
cognate domains. FASEB J.14:231–241.
19.Kuo, M. Y., M. Chao, and J. Taylor.1989. Initiation of replication of the human hepatitis delta virus genome from cloned DNA: role of delta antigen.
J. Virol.63:1945–1950.
20.Landolt-Marticorena, C., K. A. Williams, C. M. Deber, and R. A. Reithmeier.
1993. Non-random distribution of amino acids in the transmembrane
seg-ments of human type I single span membrane proteins. J. Mol. Biol.229:
602–608.
21.Lee, C. H., S. C. Chang, C. H. Wu, and M. F. Chang. 2001. A novel chromosome region maintenance 1-independent nuclear export signal of the large form of hepatitis delta antigen that is required for the viral assembly.
J. Biol. Chem.276:8142–8148.
22.Lee, C. Z., P. J. Chen, and D. S. Chen.1995. Large hepatitis delta antigen in packaging and replication inhibition: role of the carboxyl-terminal 19 amino
acids and amino-terminal sequences. J. Virol.69:5332–5336.
23.Lee, C. Z., P. J. Chen, M. M. Lai, and D. S. Chen.1994. Isoprenylation of large hepatitis delta antigen is necessary but not sufficient for hepatitis delta
virus assembly. Virology199:169–175.
24.Ma, B., T. Elkayam, H. Wolfson, and R. Nussinov.2003. Protein-protein interactions: structurally conserved residues distinguish between binding
sites and exposed protein surfaces. Proc. Natl. Acad. Sci. USA100:5772–
5777.
25.Niesters, H. G., R. A. De Man, S. D. Pas, E. Fries, and A. D. Osterhaus.2002. Identification of a new variant in the YMDD motif of the hepatitis B virus polymerase gene selected during lamivudine therapy. J. Med. Microbiol.
51:695–699.
26.O’Malley, B., and D. Lazinski.2002. A hepatitis B surface antigen mutant that lacks the antigenic loop region can self-assemble and interact with the
large hepatitis delta antigen. J. Virol.76:10060–10063.
27.O’Malley, B., and D. W. Lazinski.2005. Roles of carboxyl-terminal and farnesylated residues in the functions of the large hepatitis delta antigen.
J. Virol.79:1142–1153.
28.Patzer, E. J., G. R. Nakamura, C. C. Simonsen, A. D. Levinson, and R. Brands.1986. Intracellular assembly and packaging of hepatitis B surface
antigen particles occur in the endoplasmic reticulum. J. Virol.58:884–892.
29.Paulij, W. P., P. L. de Wit, C. M. Sunnen, M. H. van Roosmalen, A. Petersen-van Ettekoven, M. P. Cooreman, and R. A. Heijtink.1999. Localization of a unique hepatitis B virus epitope sheds new light on the structure of hepatitis
B virus surface antigen. J. Gen. Virol.80:2121–2126.
30.Persson, B., and P. Argos.1994. Prediction of transmembrane segments in
proteins utilising multiple sequence alignments. J. Mol. Biol.237:182–192.
31.Polson, A. G., B. L. Bass, and J. L. Casey.1996. RNA editing of hepatitis
delta virus antigenome by dsRNA-adenosine deaminase. Nature380:454–
456.
32.Ponzetto, A., P. J. Cote, H. Popper, B. H. Hoyer, W. T. London, E. C. Ford, F. Bonino, R. H. Purcell, and J. L. Gerin.1984. Transmission of the hepatitis B virus-associated delta agent to the eastern woodchuck. Proc. Natl. Acad.
Sci. USA81:2208–2212.
33.Radjef, N., E. Gordien, V. Ivaniushina, E. Gault, P. Anais, T. Drugan, J. C. Trinchet, D. Roulot, M. Tamby, M. C. Milinkovitch, and P. Deny.2004. Molecular phylogenetic analyses indicate a wide and ancient radiation of African hepatitis delta virus, suggesting a deltavirus genus of at least seven
major clades. J. Virol.78:2537–2544.
34.Ryu, W. S., M. Bayer, and J. Taylor.1992. Assembly of hepatitis delta virus
particles. J. Virol.66:2310–2315.
35.Simon, S., E. Krejci, and J. Massoulie. 1998. A four-to-one association between peptide motifs: four C-terminal domains from cholinesterase as-semble with one proline-rich attachment domain (PRAD) in the secretory
pathway. EMBO J.17:6178–6187.
36.Sureau, C., C. Fournier-Wirth, and P. Maurel.2003. Role of N glycosylation of hepatitis B virus envelope proteins in morphogenesis and infectivity of
hepatitis delta virus. J. Virol.77:5519–5523.
37.Sureau, C., B. Guerra, and R. E. Lanford.1993. Role of the large hepatitis B virus envelope protein in infectivity of the hepatitis delta virion. J. Virol.
67:366–372.
38.Torresi, J.2002. The virological and clinical significance of mutations in the overlapping envelope and polymerase genes of hepatitis B virus. J. Clin.
Virol.25:97–106.
39.Torresi, J., L. Earnest-Silveira, G. Civitico, T. E. Walters, S. R. Lewin, J. Fyfe, S. A. Locarnini, M. Manns, C. Trautwein, and T. C. Bock.2002. Restoration of replication phenotype of lamivudine-resistant hepatitis B virus mutants by compensatory changes in the “fingers” subdomain of the viral polymerase selected as a consequence of mutations in the overlapping
S gene. Virology299:88–99.
40.Torresi, J., L. Earnest-Silveira, G. Deliyannis, K. Edgtton, H. Zhuang, S. A. Locarnini, J. Fyfe, T. Sozzi, and D. C. Jackson.2002. Reduced antigenicity of the hepatitis B virus HBsAg protein arising as a consequence of sequence changes in the overlapping polymerase gene that are selected by lamivudine
therapy. Virology293:305–313.
41.Vietheer, P. T., H. J. Netter, T. Sozzi, and A. Bartholomeusz.2005. Failure of the lamivudine-resistant rtM204I hepatitis B virus mutants to efficiently
support hepatitis delta virus secretion. J. Virol.79:6570–6573.
42.Wang, C. J., P. J. Chen, J. C. Wu, D. Patel, and D. S. Chen.1991. Small-form hepatitis B surface antigen is sufficient to help in the assembly of hepatitis
delta virus-like particles. J. Virol.65:6630–6636.
43.Yau, W. M., W. C. Wimley, K. Gawrisch, and S. H. White. 1998. The
preference of tryptophan for membrane interfaces. Biochemistry37:14713–
14718.