Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Norovirus Capture with Histo-Blood Group Antigens Reveals Novel
Virus-Ligand Interactions
Patrick R. Harrington,
1† Jan Vinje´,
2Christine L. Moe,
3and Ralph S. Baric
1,4*
Department of Microbiology and Immunology, School of Medicine,1Department of Environmental Sciences and
Engineering,2and Department of Epidemiology, School of Public Health,4University of North Carolina
at Chapel Hill, Chapel Hill, North Carolina 27599, and Department of International Health, Rollins School of Public Health, Emory University, Atlanta, Georgia 303223
Received 9 September 2003/Accepted 18 November 2003
Noroviruses are genetically diverse, uncultivable, positive-sense RNA viruses and are the most common cause of epidemic acute gastroenteritis in humans in the United States. Recent studies of norovirus attachment in vitro by using recombinant virus-like particles (VLPs) suggest that various norovirus strains exhibit different patterns of attachment to ABH histo-blood group antigens, which are carbohydrate epitopes present in high concentrations on mucosal cell surfaces of the gut. However, attachment of live norovirus strains to histo-blood group antigens has not been investigated to date. Utilizing a newly designed magnetic bead-virus capture method, we characterized histo-blood group antigen attachment properties of various norovirus strains obtained from clinical stool specimens to compare the attachment properties of wild-type virus and VLPs and to further map norovirus attachment. Consistent with previous reports using VLPs, various strains of noroviruses exhibited different patterns of attachment to histo- blood group antigens. Norwalk virus bound specifically to H type 1, H type 3, and Leb. Two genogroup II noroviruses, one representing the Toronto
genotype and the other from a novel genotype, bound specifically to Leb. A Desert Shield-like strain did not
attach to H types 1, 2, or 3, H type 1 and 3 precursors, Lea, or Leb. Surprisingly, wild-type Snow Mountain virus
(SMV) attached specifically to H type 3, which contradicted previous findings with SMV VLPs. On further investigation, we found that stool components promote this attachment, providing the first known observation that one or more components of human feces could promote and enhance norovirus attachment to histo-blood group antigens.
Noroviruses are a group of genetically diverse RNA viruses in the familyCaliciviridaethat account for⬎90% of outbreaks of acute nonbacterial gastroenteritis in humans in the United States (18, 22). Noroviruses cause an estimated 23 million infections every year nationwide, and although serious compli-cations are not common, infection can result in hospitalization and/or death (40, 43). Noroviruses are transmitted via inges-tion of feces-contaminated water and food, exposure to aero-solized feces and vomitus, exposure to contaminated surfaces, and direct person- to-person contact, with disease symptoms including nausea, vomiting, and diarrhea. Outbreaks typically occur in areas of close human contact such as schools, day care centers, medical care facilities, cruise ships, and the military (2, 10, 22).
Because a tissue culture model for human norovirus infec-tion is not available, recombinant molecular approaches have been necessary to dissect their basic biological properties. The genome of Norwalk virus (NV), the prototype norovirus strain, has been cloned, sequenced, and characterized biochemically (35). The genome is a single-stranded, positive- sense RNA of about 7.5 kb and is organized into three open reading frames (ORFs). ORF1 encodes the nonstructural proteins, which
in-clude putative polymerase, protease, and helicase motifs. ORF2 encodes a major capsid protein, and ORF3 encodes a minor structural protein (21, 34).
Noroviruses are extremely genetically and antigenically di-verse. Based on sequence homologies in the polymerase and capsid genes, the major human strains can be genetically di-vided into two main genogroups and at least 15 different ge-notypes (51). In addition, at least three other norovirus geno-groups have been identified, which include strains detected in calves (genogroup III), humans (genogroup IV), and mice (genogroup V) (1, 17, 37, 38, 48). The major capsid proteins differ by as much as 30% in amino acid sequence between isolates within a given genogroup and by more than 50% be-tween isolates in different genogroups (23).
The ORF2-encoded major capsid protein self-assembles into virus-like particles (VLPs) when expressed in high con-centrations from recombinant baculoviruses in insect cells (34) or from Venezuelan equine encephalitis virus replicon vectors in mammalian cells (5, 27, 29). Norovirus VLPs are morpho-logically and antigenically indistinguishable from wild-type vi-rus and have provided the foundation for studies of assembly, attachment, and immunobiology. Moreover, they have been indispensable for the development of diagnostic immunoassays and are also the basis for multiple candidate vaccine strategies (4, 5, 25, 27, 29, 36).
Despite the lack of a tissue culture system or an animal model that accurately depicts human norovirus disease, signif-icant advances in norovirus attachment and pathogenesis
re-* Corresponding author. Mailing address: Department of Epidemi-ology, School of Public Health, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599-7435. Phone: (919) 966-3895. Fax: (919) 966-2089. E-mail: rbaric@sph.unc.edu.
† Present address: Lineberger Cancer Center, University of North Carolina at Chapel Hill, Chapel Hill, NC 27599-7295.
3035
on November 8, 2019 by guest
http://jvi.asm.org/
search have been achieved, including the identification of par-ticular ABH histo-blood group antigens as putative ligands for the attachment of noroviruses to mucosal cell surfaces (27, 33, 42; reviewed in reference 28). These carbohydrate epitopes are present in mucosal secretions and on many tissues throughout the human body. The complexity of the ABH histo-blood group antigens is reflected in the large number of type chain branches and modifying enzymes, as well as the broad distri-bution of different antigens based on blood type, genetic poly-morphisms, and tissue specificity (41). At least five type chains of the ABH histo-blood group antigens have been identified, which are produced and distributed in various tissue compart-ments in a type- specific manner (47). The type 1 and 3 chain molecules are the predominant ABH histo- blood group anti-gens on cell surfaces of the gut, which are presumed to be the site(s) of norovirus infection. The FUT2 enzyme, an ␣ 1,2-fucosyltransferase, is responsible for the production of type 1 and 3 chain ABH histo-blood group antigens. Approximately 20% of Europeans and Africans express a defective form of the FUT2 enzyme and therefore do not produce type 1 or 3 chain histo-blood group antigens on cell surfaces of the gut or in mucosal secretions; this is known as the “nonsecretor” or se-cretor-negative phenotype.
The type 1 and 3 H antigens are produced by the FUT2 enzyme in secretor-positive individuals from H precursor mol-ecules. The A or B enzymes produced in blood type A or B individuals, respectively (or both in blood type AB individuals), further modify the H antigens along the particular type chain to produce type-specific A or B antigens. Similarly, the type 2 chain A or B antigens are produced on red blood cells, and the presence or absence of one or more of these molecules is used to determine an individual’s blood type. In the type 1 chain, the FUT3 Lewis enzyme, an␣1,4-fucosyltransferase, modifies the H type 1 precursor to produce Lea or modifies H type 1 to
produce Leb. Approximately 10% of Europeans lack a
func-tional FUT3gene product, representative of the Lewis-nega-tive phenotype, and therefore will not produce Leaor Lebon
mucosal surfaces (41). Secretor-positive, Lewis-positive indi-viduals will produce Leband relatively less Leaon cell surfaces
of the gut and in mucosal secretions, whereas secretor-nega-tive, Lewis-positive individuals will produce only Lea.
Recombinant NV VLPs were first shown to attach to H types 1 and 3 epitopes on the surfaces of gastroduodenal epi-thelial cells from secretor-positive individuals (42). In addition, we demonstrated that Norwalk VLPs attach specifically to H type 1, H type 3, and Lebcarbohydrates, even in the absence of
other cellular components (27). The significance of these in vitro findings was recently demonstrated in vivo by Lindesmith et al. using a human challenge model for Norwalk virus infec-tion (39). This study demonstrated that secretor-negative hu-man volunteers, who do not produce H type 1, H type 3, and/or Lebon mucosal cell surfaces, were resistant to live NV
chal-lenge. Taken together, the results of these studies suggest that H type 1, H type 3, and Lebare attachment molecules
neces-sary for NV infectivity in the gut.
We have also observed that VLPs of other norovirus strains exhibit different properties of attachment to ABH histo-blood group antigens in vitro (27), suggesting that different norovirus genotypes may utilize different mechanisms for attachment and entry in vivo and that various other host factors may play
a role in susceptibility to different norovirus strains. Because VLP reagents are not available for many norovirus strains, the overall extent to which noroviruses utilize ABH histo-blood group antigens for attachment is unclear and cannot be thor-oughly evaluated using previously described biochemical meth-ods. Also, no study to date has examined and compared the ability of wild-type virus and recombinant VLPs to attach to specific ABH histo-blood group antigens.
In this study, we developed a method to purify noroviruses from clinical specimens and at the same time characterize their histo-blood group antigen attachment properties. Consistent with our previous observations, various norovirus strains ex-hibited distinct histo-blood group antigen binding patterns. In addition, we observed that attachment of one particular noro-virus strain to H type 3 occurs only in the presence of stool contents, suggesting a potential mechanism of norovirus at-tachment enhancement by a component of the extracellular environment. The methods and results described in this study may ultimately be useful in mapping the ABH histo-blood group antigen attachment specificities of all Norovirus strains, resulting in the establishment of novel hypotheses regarding norovirus pathogenesis and host susceptibility.
MATERIALS AND METHODS
Norovirus specimens.Norovirus clinical stool specimens were obtained from individuals challenged with live NV or Snow Mountain virus (SMV) or from various norovirus outbreak samples. The norovirus outbreaks occurred over at least a 5-year period, and the viruses were archived and previously demonstrated to represent various norovirus genotypes (52; C. Moe, unpublished data; J. Vinje´, unpublished data). Various strains from the outbreak specimens were identified and retyped in a blind fashion by reverse transcription-PCR (RT-PCR) followed by DNA sequencing of the RT-PCR products. When necessary to produce a
liquid suspension,⬃20% stool/phosphate-buffered saline (PBS) sample solutions
were made and clarified by low-speed centrifugation. Viral RNA was extracted from stool samples using Qiamp viral RNA minikits (Qiagen, Valencia, Calif.). A broadly reactive RT-PCR amplification targeting a 327-bp region of the poly-merase gene was performed using a One-Step RT-PCR kit (Qiagen) and mod-ified JV12/JV13 primers (52). RT-PCR-positive products were sequenced and aligned to previously published sequences to identify specimens that represent distinct norovirus genotypes. Human challenge specimens and outbreak samples representing different norovirus genotypes were then subjected to titer determi-nation by end-point dilution RT-PCR using the same primers described above, except for Norwalk virus, where strain-specific primers were used (15).
Viral capture and purification method.A total of 50l of streptavidin-coated magnetic bead suspensions (M270; Dynal Biotech, Oslo, Norway) were rinsed three times with 1 ml of sterile PBS in a 1.5-ml microcentrifuge tube. The beads were then preblocked with 5% milk in PBS (5% BLOTTO) for 1 h at ambient temperature on an end-over-end mixer. Synthetic, biotinylated histo-blood group antigens were obtained from Glycotech (Rockville, Md.) and resuspended in
PBS at 1 mg/ml. A 50-l volume of the synthetic carbohydrates was added to the
preblocked magnetic beads, and the mixture was incubated at ambient temper-ature for 2 h on an end-over-end mixer. The beads were then rinsed three times with PBS-Tween (0.05% Tween) and blocked for 1 h at ambient temperature and then overnight at 4°C in 5% BLOTTO containing 0.25% Tween, while contin-ually kept on an end-over-end mixer. The beads were then rinsed three times with PBS-Tween and resuspended in 5% BLOTTO. A norovirus-containing stool
specimen (1 to 10l) with 105PCR-detectable units of virus was then added to
the bead suspensions, and the mixture was incubated at ambient temperature for 4 h on an end-over-end mixer. The beads were then washed five times with PBS-Tween and three times with sterile PBS and subsequently resuspended in 50
l of sterile, RNase-free PBS. The washed beads were then 10-fold serially
diluted three times and stored at⫺80°C until tested by RT-PCR.
RT-PCR detection of norovirus attachment.RT-PCR detection of viral at-tachment was conducted in a blind fashion, with no prior knowledge of the norovirus strains or histo- blood group carbohydrates being tested for attach-ment. First, encapsidated viral RNA was heat released from the bead
suspen-sions by incubating 10l of each bead dilution at 99°C for 5 min. Then 2.5l of
on November 8, 2019 by guest
http://jvi.asm.org/
the heat-released RNA, as well as RNA extracted and purified from the original stool sample as a positive control, was subjected to RT-PCR using the same primers and conditions for determining the titers of the samples. RT-PCR products were examined on an ethidium bromide-stained 2% agarose gel.
VLP attachment assays.Two microwell-based assays were designed to detect and quantify norovirus VLP attachment to histo-blood group antigens. In the
first assay, synthetic biotinylated carbohydrates (2.5g/ml in Tris-buffered saline,
100 l per well) were added to streptavidin-precoated microwells (Thermo
Labsystems, Basingstoke, United Kingdom) and the mixtures were incubated for
2 h at 37°C. The microwells were then blocked with 300l of 5% BLOTTO
overnight at 4°C. Sucrose gradient-purified norovirus VLPs (5, 27) were then added to the carbohydrate-coated microwells at various concentrations and in the presence or absence of various stool extracts (as indicated in Results). The wells were incubated at 37°C for 4 h and then washed five times with PBS-Tween.
To detect attachment, 100l of pooled polyclonal antisera from SMV, NV, and
Hawaii virus (HV)-infected human volunteers was added per well at a 1:100 dilution in 5% BLOTTO and the wells were incubated at 37°C for 2h. The plates were then washed five times with PBS-Tween, a goat anti-human
immunoglob-ulin G (IgG)-alkaline phosphatase conjugate (1:1,000 in 5% BLOTTO, at 100l/
well) (Sigma, St. Louis, Mo.) was added, and the mixture was incubated for 1 h
at 37°C. After a final washing step,p-nitrophenyl phosphatase substrate (pNPP)
(Sigma-FAST tablets; Sigma) was added at 150l/well and the plates were
in-cubated for 25 to 30 min at ambient temperature. Developed plates were then
read at an optical density of 405 nm (OD405) using a microplate reader (Molecular
Devices, Sunnyvale, Calif.). Wells with no VLPs added were used as plate blanks. A second VLP attachment assay was used to detect the attachment of carbo-hydrates to VLP-coated microwells, similar to previously described methods (27).
Briefly, 2.5g of the appropriate VLP reagent per ml (100l/well in Tris-buffered
saline) was added to high-binding microwells (Costar, Corning, N.Y.) and the plates were incubated for 4 h at ambient temperature and then blocked with 5% BLOTTO overnight at 4°C. Synthetic biotinylated histo-blood group antigens were then added at various concentrations in the presence or absence of stool extracts (as indicated in Results) in 5% BLOTTO, and the plates were incubated at 37°C for 4 h. The plates were then washed five times with PBS-Tween, streptavidin-alkaline phosphatase conjugate (1:500 dilution in 5% BLOTTO, at
100l/well) (Sigma) was added, and the mixture was incubated at 37°C for 1 h.
Finally, the plates were washed and developed with pNPP and the OD405was
determined. Wells with no carbohydrate added were used as plate blanks.
Stu-dent’sttest (unpaired, two-tailed, unequal variance) was used to determine the
statistical significance of stool extract-enhanced attachment of SMV VLPs.
RESULTS
Identification and genotyping of clinical norovirus speci-mens.To identify high-titer clinical specimens to be used for this study, we first screened and determined the titers of no-rovirus stool specimens obtained from human challenge stud-ies or outbreak samples for the presence of noroviruses by RT-PCR using both broadly reactive and NV-specific primer pairs. We then sequenced RT-PCR products from norovirus-positive samples and aligned them with previously published norovirus sequences to determine the genotype of the viruses. Finally, we determined the titers of norovirus stool specimens representing various genotypes by end-point RT-PCR using the same primer sets used to identify the norovirus strains. Stool samples with norovirus titers of 107 to 108
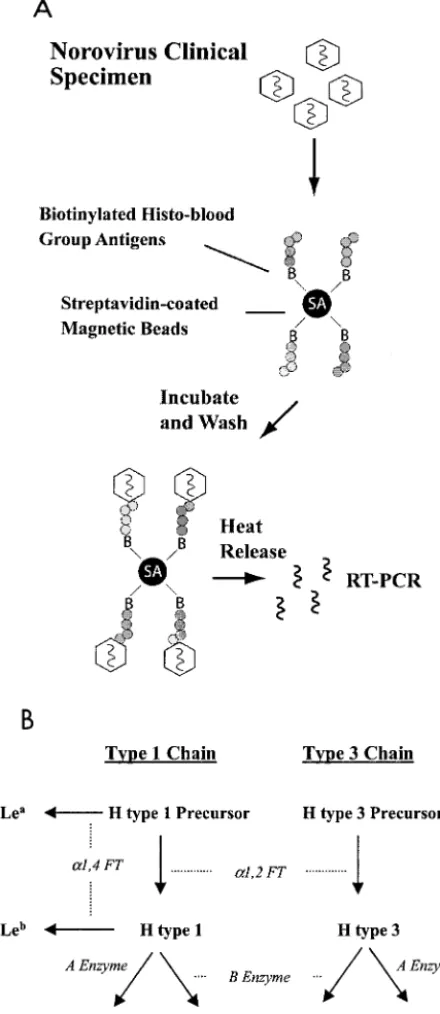
[image:3.603.53.273.73.580.2]RT-PCR-detectable units were identified and used for attachment char-acterization assays. For a genetic classification of the strains used in this study and a general summary of attachment data, see Fig. 10.
FIG. 1. Magnetic bead method and carbohydrates used in this study. (A) A magnetic bead-based method was designed to character-ize histo-blood group antigen attachment specificities of wild-type no-roviruses obtained from clinical stool specimens. Streptavidin-pre-coated magnetic beads were Streptavidin-pre-coated with specific synthetic, biotinylated histo-blood group antigens and then blocked with 5% milk–PBS con-taining 0.25% Tween 20 to inhibit nonspecific binding. Stool extracts containing equal RT-PCR detectable levels of particular norovirus strains were then incubated with the magnetic beads. After extensive washing to remove unbound virus, the magnetic beads were 10-fold serially diluted and subjected to heat release to extract the RNA genome. The virus-bead suspensions were diluted for two reasons: (i) to generate a semiquantitative result of viral attachment and (ii) to dilute any possible inhibitors of the RT-PCR amplification. Finally, RT-PCR was performed to assay for the presence of viral RNA to detect the attachment of the virus to the specific histo-blood group
antigens. (B) Type 1 and 3 chain histo-blood group antigens used in this study are shown in bold type. Enzymatic activities required to pro-duce the various carbohydrates are shown in italics. H type 2 antigen, which is not present in high concentrations on the superficial surfaces of the gut (47), was also used in this study as a tissue-specific control.
on November 8, 2019 by guest
http://jvi.asm.org/
Norovirus capture and histo-blood group antigen attach-ment characterization.We designed and optimized a magnetic bead-based method to capture and characterize the histo-blood group antigen attachment properties of noroviruses in clinical stool specimens (Fig. 1A). Using this assay, we evalu-ated the ability of noroviruses representing five different ge-notypes to attach to synthetic carbohydrates representative of the various histo-blood group antigens that would be present on gut mucosal cell surfaces of individuals of different secretor and Lewis phenotypes (Fig. 1B). The histo-blood group anti-gen attachment properties of NV, a anti-genogroup I (GI) strain and the prototype norovirus (GI.1 genotype) have been previ-ously characterized using VLP reagents and a variety of bio-chemical and histological methods (27, 33, 42). Consistent with these studies, NV obtained from a human challenge stool spec-imen (39) bound specifically to H type 1, Leb, and, to a
rela-tively lesser extent, H type 3 (Fig. 2), suggesting that NV VLPs and wild-type virus are biologically indistinguishable in the case of attachment to these histo-blood group antigens.
Also consistent with our previous studies using VLP
re-agents (27), different norovirus strains exhibited a variety of histo-blood group antigen attachment phenotypes (Fig. 2). A second GI strain, M23, representative of the Desert Shield/ Stavanger-virus genotype (GI.3) did not attach to any of the histo-blood group antigens tested. This observation was firmed by repeating the experiment with a 10-fold-higher con-centration of virus (data not shown). Two GII strains, M15 and M116, bound specifically only to Leb. The M15 virus is
repre-sentative of the Toronto (GII.3) virus genotype, while M116 represents a novel norovirus genotype (52).
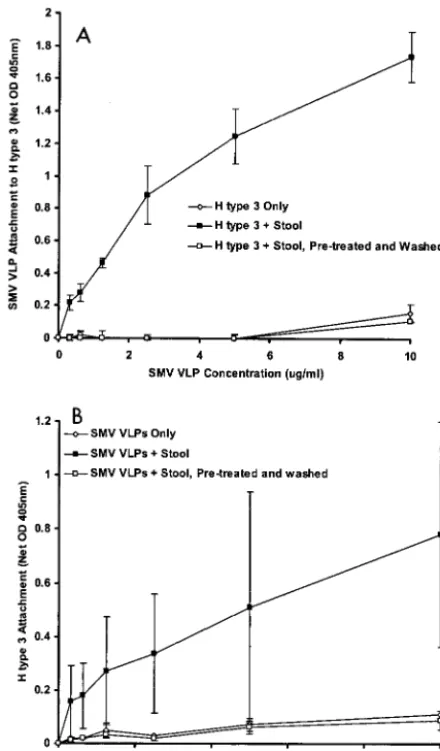
Finally, SMV, which is a prototype GII strain (GII.2), was obtained from a human challenge stool specimen and tested for attachment to the various histo-blood group antigens. Pre-vious studies from our laboratory demonstrated that SMV VLPs did not attach to any of the synthetic carbohydrates used in this study (27). However, we observed that SMV from the clinical stool specimen did in fact attach specifically to H type 3 (Fig. 2).
[image:4.603.106.482.66.443.2]SMV attachment characterization.We further investigated the SMV-H type 3 attachment properties to identify the cause
FIG. 2. Histo-blood group antigen attachment specificities of noroviruses obtained from clinical specimens. RT-PCR-positive products indicate attachment of norovirus strains to the specific synthetic histo-blood group antigens shown. RNA samples extracted from the same clinical stool specimens subjected to the attachment assay were used as positive controls (lane⫹), and sterile water was used for negative controls (lane⫺). “No Carbohydrate” represents mock-coated magnetic beads as a negative control for nonspecific viral binding.
on November 8, 2019 by guest
http://jvi.asm.org/
of the discrepancy between the SMV VLP and clinical virus attachment observations. First, we tested if SMV VLPs will compete with the virus for attachment to H type 3. Using the magnetic bead method described above, the SMV stool spec-imen was added to H type 3-coated magnetic beads in either the absence or presence of excess SMV VLPs (10g, sucrose pure) or to beads pretreated with the same amount of SMV VLPs and then washed with PBS before addition of the SMV stool specimen. Interestingly, SMV VLPs outcompeted the virus to attach to H type 3 (Fig. 3). However, the SMV VLPs could be washed from the H type 3-coated beads before addi-tion of the SMV stool specimen, restoring the ability of the virus to attach to H type 3 (Fig. 3).
We then used a series of microwell-based assays to further characterize SMV VLP attachment to H type 3. Using a bio-chemical microwell method, which previously established that SMV VLPs do not attach to H type 3 (27), we first character-ized the attachment of three additional batches of sucrose-purified SMV VLPs. Consistent with the previous results, syn-thetic, biotinylated H type 3 did not attach to microwells coated with any of the three different batches of SMV VLPs (data not shown).
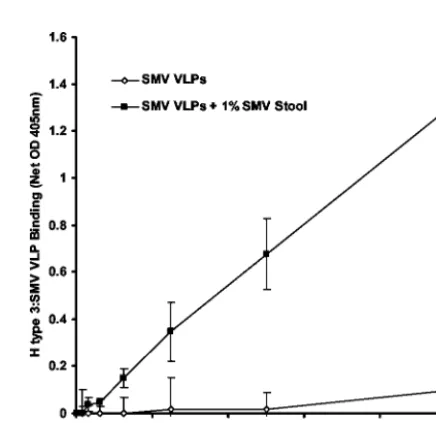
Using another method to detect VLP attachment to carbo-hydrate-coated microwells, similar to the method previously described by Hutson et. al. (33), SMV VLPs again did not attach to H type 3-coated microwells (Fig. 4). However, when the SMV stool specimen was added to sample buffer along with the VLPs, attachment to H type 3 was detected (Fig. 4). The binding signal appeared in a linear fashion in relation to VLP concentration, suggesting that the SMV VLPs and not the SMV virions in stool attached to H type 3.
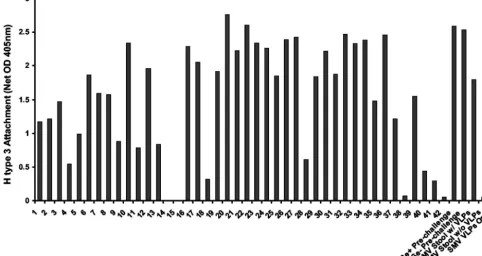
Stool contents promote SMV attachment to H type 3.To determine if SMV VLP attachment is dependent on the pres-ence of SMV virions or on another component of the extra-cellular environment in the clinical sample, we investigated the ability of other stool extracts to enhance SMV VLP attachment to H type 3. Forty-two archived clinical stool specimens from various norovirus outbreaks, representing at least 10 different norovirus genotypes, were screened for their ability to enhance SMV VLP attachment to H type 3. Remarkably, nearly all of the extracts promoted SMV VLP attachment to some extent (Fig. 5). The mean attachment (net OD405⫾standard
devia-tion) of SMV VLPs to H type 3 in the presence of the stool samples was 1.57⫾0.84, whereas attachment of three different batches of SMV VLPs to H type 3 in the absence of stool samples was 0.08⫾0.01. The difference in the ability of SMV VLPs to attach to H type 3 in the presence and absence of stool
extracts is significant (P⬍0.001; Student’sttest). No correla-tion was found between attachment enhancement levels and the presence, absence, genotype, or titer of noroviruses in the outbreak specimens (data not shown). In fact, even prechal-lenge, virus-free stool extracts from human volunteers pro-moted SMV attachment regardless of the individuals’ secretor phenotype (Fig. 5). Therefore, stool contents alone are most likely promoting SMV attachment to H type 3, suggesting that a component of the extracellular environment in normal hu-man feces might play a role in SMV attachment in vivo.
We further investigated the attachment enhancement mech-anism to characterize its specificity for SMV and H type 3. The ability of the SMV stool sample to enhance SMV VLP attach-ment was specific for H type 3, since attachattach-ment to H type 1, the H type 3 precursor, or mock-coated microwells was not detected even in the presence of the stool specimen (Fig. 6). In fact, neither wild-type SMV nor SMV VLPs attached to any of the carbohydrates used in this study other than H type 3, regardless of the presence or absence of stool extracts (Fig. 2 and data not shown).
The attachment enhancement mechanism was not necessar-ily specific for SMV. The ability of NV VLPs to attach to H type 1, H type 3, or Lebwas not enhanced by the presence of
[image:5.603.48.281.69.116.2]stool contents (Fig. 7A). In addition, no enhancement effect on NV VLP attachment with other histo-blood group antigens was evident (data not shown). However, we previously dem-onstrated that HV VLPs do not attach to any of the histo-blood group antigens used in this study (27). Although the
FIG. 3. Attachment of wild-type SMV to H type 3 in the presence and absence of VLPs. SMV from a clinical stool specimen was added to H type 3-coated magnetic beads and assayed for attachment by RT-PCR as described in the legend to Fig. 1. The effects of adding excess sucrose-purified SMV VLPs (10g) to the attachment reaction mixture (lanes⫹SMV VLPs) or preincubating and then washing the VLPs prior to adding the SMV clinical stool specimen (lanes⫹SMV VLPs, washed) were investigated.
FIG. 4. Attachment of SMV VLPs to H type 3. The ability of SMV VLPs to attach to H type 3 in the presence or absence of the SMV clinical stool extract was investigated by using a microwell-based at-tachment assay. Wells were coated with H type 3 and blocked over-night, and the indicated protein concentrations of SMV VLPs (with or without 1% [vol/vol] stool extract in the sample buffer) were added. The wells were then washed, and VLP attachment was quantified by using human polyclonal anti-norovirus antiserum, goat anti-human IgG-alkaline phosphatase conjugate, and pNPP substrate. Net OD 405nm represents the OD405values corrected for blank OD405values
(no VLPs added). Mean values from triplicate wells are shown, and error bars represent the standard deviation.
on November 8, 2019 by guest
http://jvi.asm.org/
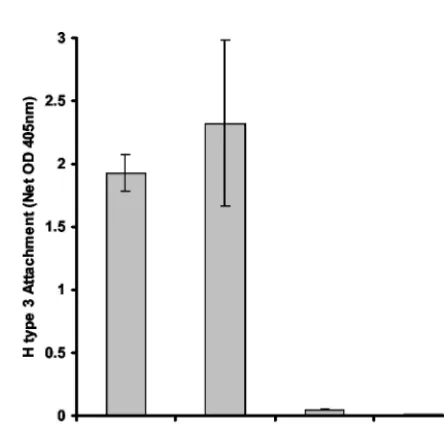
[image:5.603.311.529.403.614.2]effect was not as robust as for the SMV VLPs, addition of stool extracts modestly promoted HV VLP attachment specifically to Leb(Fig. 7B).
We further characterized the attachment enhancement mechanism to begin to dissect the components of stool that might play a role. Enzyme-linked immunosorbent assays of the stool extracts were performed to determine if antibody might play a role in SMV attachment enhancement. Of the 42 dif-ferent outbreak specimens tested, only a small fraction had detectable levels of norovirus-specific IgA, IgM, or IgG, with no correlation between the presence of detectable norovirus-specific antibody and the ability of the stool extracts to enhance SMV attachment (data not shown). In fact, denaturing the stool samples by boiling did not impair their ability to enhance SMV attachment to H type 3 (Fig. 8), suggesting that neither antibody, enzymes, nor other protein components in stool are responsible for SMV attachment enhancement.
We then determined if pretreatment of the SMV VLPs or H type 3 with the stool extracts was sufficient to detect the at-tachment of SMV VLPs to H type 3. Before the addition of SMV VLPs, H type 3-coated microwells were pretreated with stool extracts for 2 h and washed to remove unbound stool components. Similarly, SMV VLP-coated microwells were pre-treated with stool components and washed before addition of synthetic H type 3. In both cases, SMV VLP attachment to H type 3 was evident only when all three components—stool extract, SMV VLPs, and carbohydrate—were present at the same time (Fig. 9). Pretreatment of the SMV VLPs or H type 3 with stool extracts was not sufficient to promote attachment.
DISCUSSION
Several recent studies by multiple investigators have impli-cated the histo-blood group antigens as putative receptors for NV (27, 31, 33, 42). The importance of these molecules in vivo
[image:6.603.49.532.75.331.2]FIG. 5. Effect of nonspecific stool extracts on SMV VLP attachment to H type 3. The effect of various archived stool extracts on SMV VLP attachment (10 g/ml) to H type 3 was investigated by using the same microwell-based assay described in the legend to Fig. 4. Lanes Se⫹ Pre-challenge and Se⫺Pre-challenge contain virus-free stool extracts from secretor-positive and secretor-negative human volunteers, respectively, obtained prior to norovirus challenge.
FIG. 6. SMV VLP attachment to other histo-blood group antigens in the presence of stool extract. Attachment of SMV VLPs in the presence of stool extract was investigated using the same microwell-based assay described in the legends to Fig. 4 and 5. A pool of no. 22, no. 27 (numbers are shown in Fig. 5), and SMV stool extracts was used at a final concentration of 1% in the sample buffer when added with the VLPs. Mean OD405values from duplicate wells are shown, and
error bars represent the standard deviation.
on November 8, 2019 by guest
http://jvi.asm.org/
has been established using a human challenge model for NV infection in which secretor-negative individuals were resistant to live NV challenge (39). We have also previously observed three distinct patterns of binding, both to specific histo-blood group antigens and to histo-blood group antigens in saliva, for three different norovirus strains (27). These initial observations were supported by another study that revealed three potential additional binding patterns to saliva samples (31), although saliva-binding studies alone cannot distinguish which specific
molecules are involved in attachment. Taken together, these observations suggest that the ABH histo-blood group antigens are important factors in norovirus pathogenesis and that var-ious degrees of susceptibility may exist for different norovirus strains based on one’s blood type, secretor phenotype, and/or Lewis phenotype.
To our knowledge, all studies prior to this utilized only VLP reagents to characterize norovirus attachment in vitro (27, 31, 33, 42, 50, 53). In this study, we designed a method to charac-terize the attachment of wild-type noroviruses obtained from clinical specimens with specific histo-blood group antigens to (i) determine if different attachment properties between VLPs and wild-type virus exist, (ii) reduce the need to produce ad-ditional VLP reagents to investigate the attachment properties of various norovirus strains, and (iii) develop an approach to concentrate and purify wild-type noroviruses from clinical or environmental samples. We found that different norovirus strains exhibited different histo-blood group antigen attach-ment patterns, consistent with previous studies using VLPs (27). However, in the case of SMV, wild-type virus bound to H type 3 whereas our previous study with SMV VLPs demon-strated no attachment to H type 3 (27).
[image:7.603.54.273.71.471.2]Surprisingly, adding stool components to the attachment reaction mixtures promoted the specific attachment of SMV VLPs to H type 3, suggesting that the contradictory observa-tions of SMV attachment are not due to phenotypic differences in wild-type virus and VLPs. In fact, excess amounts of SMV VLPs clearly outcompeted SMV virions for attachment to H type 3-coated magnetic beads, suggesting that both the virus and VLPs attach to H type 3 in a similar manner. This phe-nomenon was not specific for SMV-positive stool specimens since stool extracts from unrelated stool samples also efficiently
FIG. 7. Effect of stool components on NV and HV VLP attach-ment to histo-blood group antigens. (A) The effect of stool on NV VLP attachment to H type 1, H type 3, and Lebwas investigated using the
same microwell-based assay described in the legends to Fig. 4 and 5. Note that different “1⫻” VLP concentrations of 0.2g/ml for H type 1 and 0.4 g/ml for H type 3 and Leb were used to account for
previously determined differences in attachment avidity of the different carbohydrates with NV VLPs (27). (B) Effect of stool extract on HV VLP attachment. Data shown are based on using 10g of HV VLP per ml. The dashed line represents an arbitrary background cutoff equiv-alent to twice the average background OD405. In all assays, a pool of
no. 22, no. 27 (numbers from fig. 5), and SMV stool extracts was used at a final concentration of 1% in the sample buffer when added with the indicated VLPs.
FIG. 8. Effect of denaturing stool extracts on SMV VLP attach-ment to H type 3. Stool extracts (no. 22, no. 27 [numbers from fig. 5], and SMV) were denatured by boiling for 10 min before being added to the sample buffer with SMV VLPs (10g of VLPs per ml plus 1% stool extract). Mean OD405values for SMV VLP attachment in the
presence of each of the three different extracts are shown, and error bars represent the standard deviation.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:7.603.310.532.447.660.2]promoted the attachment of SMV VLPs to H type 3 in mi-crowell-based assays. Because noroviruses are typically trans-mitted by the fecal-oral route, the findings that both SMV and HV VLPs attached to histo-blood group antigens only in the presence of fecal extracts suggest that one or more stool com-ponents, as elements of the extracellular environment, might play a role in norovirus attachment in vivo. We therefore speculate that identification of this attachment enhancement
factor could potentially lead to a target for novel, broadly reactive norovirus treatments and/or vaccines.
It is well established that the infectivity of many viral patho-gens is enhanced by components of the extracellular environ-ment. An extracellular proteolytic event that probably occurs in the intestine enhances the infectivity of several other enteric viruses, such as rotaviruses and reoviruses, probably at the level of attachment and entry (3, 6–8, 12, 13). Also, the infec-tivity of certain other viral pathogens, such as Dengue virus, is enhanced by the presence of virus-specific antibody (26, 44, 49).
It is unclear which particular component of stool might be promoting or enhancing norovirus attachment to histo-blood group antigens. Our results suggest that antibody, enzymes, and other protein molecules are not playing any role since denaturing the stool extracts did not impair their ability to enhance SMV VLP attachment to H type 3. In addition, ex-tracts obtained from secretor-negative and secretor-positive individuals enhanced SMV attachment equally, suggesting that histo-blood group antigens secreted from the gut are not in-volved in promoting SMV attachment to H type 3. It is also puzzling that the stool extracts must be present with both the SMV VLPs and H type 3 to detect attachment, which suggests that the component is unlikely to be a ligand that first attaches to the virus and then links it to the carbohydrate, or vice versa. The acidity of the stool extracts is probably not playing a role, since the pH of the sample buffers used in the attachment assays does not change with the addition of stool extracts (data not shown). Divalent cations in stool extracts do not appear to play a role, since addition of MgCl2and CaCl2or EDTA to the
sample buffers does not remarkably alter SMV VLP attach-ment properties (unpublished data). Other possible factors not ruled out in these studies include various salts, lipids, peptides, and proteins resistant to heat denaturing and other carbohy-drate molecules that could be present as components of stool. Future experiments designed to fractionate the extracts by high-performance liquid chromatography or other methods will be conducted to attempt to precisely identify the factor(s) involved in promoting or enhancing norovirus attachment.
A similar observation of fecal or intestinal contents playing a role in viral infectivity has been made with an enteric calici-virus of pigs. Porcine enteric calicicalici-virus Cowden (PEC) is the only human or animal enteric calicivirus known to replicate in cell culture (19). Interestingly, the virus is cultivable only when porcine intestinal contents are added to the medium (19, 46). Unlike SMV and HV, however, porcine intestinal contents probably do not affect the propagation of PEC in tissue culture at the level of viral attachment. Rather, a component of the intestinal contents is probably acting on cell signaling machin-ery to promote PEC replication (11). Nevertheless, the fact that fecal or intestinal contents affect the biology of multiple caliciviruses warrants further investigation of the identity and the role of environmental components in calicivirus attach-ment, infectivity, replication, and pathogenesis. Identifying and optimizing the use of such components could be essential in the development of a tissue culture or animal model system for cultivating human noroviruses, as in the case of PEC.
The magnetic bead-based technique for characterizing no-rovirus attachment described in this study not only provided a method to map histo-blood group antigen attachment
proper-FIG. 9. Effect of pretreating VLPs or carbohydrate with stool ex-tracts followed by washing on SMV VLP attachment to H type 3. (A) SMV VLPs were serially diluted and added to H type 3-coated micro-wells in either the absence or presence of 1% stool extracts or to coated wells that were pretreated for 2 h with sample buffer containing 1% stool extracts followed by washing before the addition of the VLPs. VLP attachment was quantified as described in the legends to Fig. 4 and 5. (B) Biotinylated H type 3 carbohydrates were serially diluted and added to SMV VLP-coated microwells in either the presence or absence of 1% stool extracts or to coated wells that were pretreated for 2 h with sample buffer containing 1% stool extracts followed by wash-ing before the addition of addwash-ing biotinylated H type 3. Attachment of biotinylated H type 3 to SMV VLPs was quantified by adding strepta-vidin-alkaline phosphatase conjugate and pNPP substrate. Net OD 405nm indicates OD405values corrected for plate blank values (no H
type 3 added). In both experiments, mean OD405values for attachment
in the presence or absence of three different stool extracts (no. 22, no. 27 [numbers from Fig. 5], or SMV) are shown, and error bars represent the standard deviation.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:8.603.54.274.69.444.2]ties but also was central in the observation that stool compo-nents enhance SMV attachment. Therefore, we propose that additional experiments in mapping the attachment properties of different norovirus strains should include methods to di-rectly study viruses in stool, used either alone or in combina-tion with previously described VLP-based methods. Address-ing norovirus attachment questions usAddress-ing VLP reagents alone might result in the overlooking of molecules that are recog-nized for attachment only by wild-type virus in its more natural environment. This point is clearly illustrated in the fact that our previous studies with SMV VLPs failed to identify H type 3 as a possible attachment molecule (27). In addition, the methods described in this paper will be useful in identifying other norovirus strains that attach to histo-blood group anti-gens only in the presence of stool components or in identifying additional environmental components that influence norovirus attachment.
On a more practical level, these methods could also be valu-able for concentrating, purifying, or diagnosing noroviruses in clinical or environmental samples to aid in norovirus molecular biology, epidemiology, and environmental science studies. In addition, with a more complete picture of norovirus–histo-blood group antigen interactions, the methods described in this paper might be useful for detecting and quantifying infectious virus. Unlike conventional RT-PCR diagnostics and previously described antibody-based norovirus capture methods (20), capture with histo-blood group antigen-coated magnetic beads selects for intact virions that are capable of attaching to these ligands, potentially providing a more accurate depiction of infectious viruses in the samples.
Future experiments will be aimed at mapping the histo-blood group antigen attachment patterns of noroviruses rep-resenting all of the major genogroups and genotypes. However, several limitations of the magnetic bead-based method exist, and will have to be addressed before all the different norovirus histo-blood group antigen binding patterns can be elucidated. One assay limitation is that high titers of the norovirus strains are required to detect attachment to the carbohydrate-coated beads. Although we can detect NV attachment to H type 1 over three 10-fold serial dilutions of the beads, addition of 1/10 of the clinical sample resulted in no detection of NV to H type 1, H type 3, or Leb, regardless of the bead dilution (data not
shown). Therefore, only norovirus clinical samples that have a titer of at least 105 RT-PCR-detectable units per ml of stool
suspension could be used in this assay in its present format, which screens out most norovirus-positive outbreak specimens. For this reason, although equal amounts of RT-PCR-detect-able virus were added in the experiments, we cannot com-pletely rule out the possibility of obtaining false-negative re-sults for attachment. Our laboratory is currently investigating several potential solutions to this problem, including concen-trating viruses using the magnetic-bead approach, reducing the total volume used for the assay, using new-generation strepta-vidin-coated beads, increasing the overall sensitivity of the RT-PCR amplification by using nested or real-time RT-PCR methods, and designing a flow cytometry, antibody-based method to detect viral attachment to carbohydrate-coated beads. Another potential problem is the presence of RT- PCR inhibitors in the reaction mixtures that remain after heat re-lease of the RNA genome, especially in undiluted virus-bead
suspensions. Although it would add several additional steps to the method, chemical extraction of the RNA genomes may increase the overall sensitivity of the assay.
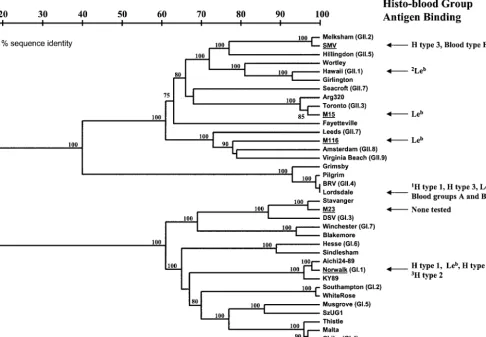
Despite these limitations, several novel observations of no-rovirus attachment have been revealed in this study. A sum-mary of the norovirus attachment data compiled with results from previous studies is shown in Fig. 10. No obvious histo-blood group antigen binding pattern based on genetic identity seems to exist, although none of the sequences analyzed be-tween strains tested in this study were more than 75% identi-cal. It is of interest that only the histo-blood group antigens that would be present in secretor-positive individuals were involved in attachment of the noroviruses analyzed in this study. We therefore speculate that secretor-negative individu-als might be genetically resistant to a variety of norovirus strains, not just NV. However, most adult humans have serum antibody against noroviruses regardless of their secretor status (14, 16, 39, 45; C. Moe et al., unpublished data), suggesting that some norovirus strains might be able to infect secretor-negative individuals. In support of these observations, VLPs of one particular norovirus stain (VA207) bound to saliva from secretor-negative individuals (31), although the VLPs also bound modestly to saliva from secretor-positive individuals. Importantly, it remains unclear whether noroviruses utilize histo-blood group antigens as primary receptor molecules, co-receptors, or attachment-enhancing molecules or if all norovi-rus strains even attach to histo-blood group antigens.
It is also important to acknowledge that data from in vitro attachment experiments alone do not directly address whether one particular population would be more resistant or suscep-tible to norovirus infection and that these hypotheses are based only on the results obtained from human challenge studies of NV susceptibility (32, 39). Only human challenge experiments with other norovirus strains will provide results to conclusively resolve the role of histo-blood group antigens in human sus-ceptibility to other noroviruses. However, results of attach-ment studies will allow the proposal of novel hypotheses of human susceptibility for other noroviruses, which can then be addressed using the human challenge model. For example, our data demonstrate that noroviruses from the M116 and M15/ Toronto genotypes can utilize Lebas an attachment molecule.
We therefore hypothesize that Lewis-negative and/or secretor-negative individuals might be more resistant to these norovirus strains than Lewis-positive and secretor-positive individuals, since either “negative” phenotype will result in the lack of Leb
production on mucosal cell surfaces of the gut. Of course, the logistics of norovirus human challenge experiments do not conveniently allow for challenge with many different norovirus strains. Epidemiology studies, however, could address these questions to provide some information about the role of histo-blood group antigens in the infectivity of other norovirus strains. In a recent report by Hennessy et al. (30), blood group O individuals appeared to be more susceptible to infection during a norovirus outbreak that occurred in a closed military hospital setting, providing indirect evidence of the role of histo-blood group antigens in the infectivity of this particular strain. Therefore, with the combination of human challenge experiments, epidemiological observations, and in vitro attach-ment studies, human susceptibility to a variety of norovirus strains can be systematically investigated.
on November 8, 2019 by guest
http://jvi.asm.org/
Identification of additional norovirus histo-blood group an-tigen binding patterns could also provide valuable insight into norovirus epidemiology and evolution. For example, the ability of noroviruses to bind to particular histo-blood group antigens could play a role in the establishment of norovirus strains selected and optimized to infect particular human populations, thus driving norovirus evolution. In fact, VLPs of Lordsdale virus, which is the genotype that has caused the majority of norovirus epidemics in recent years (9, 17), attach to at least five different specific histo-blood group antigens, listed here from highest to lowest binding avidity: H type 3, blood group B Ag, blood group A antigen, Leb, and H type 1 (unpublished
data). We speculate this particular strain has evolved to utilize a greater variety of histo-blood group antigens for attachment to enhance its infectivity in a broad range of human popula-tions.
In summary, this study has built on earlier observations of norovirus attachment and has provided the first examination of the attachment properties of wild-type noroviruses obtained from clinical stool specimens. In addition, we observed that
stool extracts could promote norovirus attachment, suggest-ing that a component of the extracellular environment might play a role in enhancing norovirus attachment and infectivity in vivo.
ACKNOWLEDGMENTS
We thank Martha Collier for producing packaged Venezuelan equine encephalitis VRPs for this study and Sjon Oudejans, Anna LoBue, Lisa Lindesmith, and Boyd Yount for helpful discussions and technical assistance.
This work was supported by grants from the U.S. Environmental Protection Agency (STAR grant R- 826139 and R-82936501) and the National Institutes of Health (AI056351, AI23946, and GM63228).
REFERENCES
1. Ando, T., J. S. Noel, and R. L. Fankhauser.2000. Genetic classification of
“Norwalk- like viruses”. J. Infect. Dis.181(Suppl. 2):S336–S348.
2. Arness, M. K., B. H. Feighner, M. L. Canham, D. N. Taylor, S. S. Monroe, T. J. Cieslak, E. L. Hoedebecke, C. S. Polyak, J. C. Cuthie, R. L. Fankhauser, C. D. Humphrey, T. L. Barker, C. D. Jenkins, and D. R. Skillman.2000. Norwalk-like viral gastroenteritis outbreak in U.S. Army trainees. Emerg.
Infect. Dis.6:204–207.
[image:10.603.50.537.74.411.2]3. Babiuk, L. A., K. Mohammed, L. Spence, M. Fauvel, and R. Petro.1977.
FIG. 10. Summary of norovirus attachment data in relation to genetic grouping. Genetic classifications of noroviruses used in this study (indicated by underlining) are based on previously described criteria and compared with known sequences of other noroviruses (genogroup.geno-type indicated) (23, 24, 51). The nucleotide sequences that were compared are from a 177-nucleotide region in the ORF2 gene (region D) that has been shown to reliably classify Norovirus genotypes (52). Specific histo-blood group antigen attachment data from this study compiled with results from other studies are indicated.1Unpublished data obtained using VLP reagents.2Previous results using HV VLPs alone showed no
binding to any carbohydrates tested (27), whereas results from this study demonstrated slight binding of VLPs to Leb, but only in the presence of
stool extract.3Observation made by Hutson et al. using baculovirus-expressed recombinant NV VLPs (33).4Based on the observation that SMV
VLPs attached specifically to saliva from secretor-positive, blood type B individuals (27).
on November 8, 2019 by guest
http://jvi.asm.org/
Rotavirus isolation and cultivation in the presence of trypsin. J. Clin.
Mi-crobiol.6:610–617.
4. Ball, J. M., M. E. Hardy, R. L. Atmar, M. E. Conner, and M. K. Estes.1998. Oral immunization with recombinant Norwalk virus-like particles induces a
systemic and mucosal immune response in mice. J. Virol.72:1345–1353.
5. Baric, R. S., B. Yount, L. Lindesmith, P. R. Harrington, S. R. Greene, F. C. Tseng, N. Davis, R. E. Johnston, D. G. Klapper, and C. L. Moe.2002. Expression and self- assembly of norwalk virus capsid protein from
venezu-elan equine encephalitis virus replicons. J. Virol.76:3023–3030.
6. Barnett, B. B., R. S. Spendlove, and M. L. Clark.1979. Effect of enzymes on
rotavirus infectivity. J. Clin. Microbiol.10:111–113.
7. Bass, D. M., D. Bodkin, R. Dambrauskas, J. S. Trier, B. N. Fields, and J. L. Wolf.1990. Intraluminal proteolytic activation plays an important role in replication of type 1 reovirus in the intestines of neonatal mice. J. Virol.
64:1830–1833.
8. Bodkin, D. K., M. L. Nibert, and B. N. Fields.1989. Proteolytic digestion of
reovirus in the intestinal lumens of neonatal mice. J. Virol.63:4676–4681.
9. Centers for Disease Control and Prevention.2003. Norovirus activity—
United States, 2002. Morb. Mortal. Wkly. Rep.52:41–45.
10. Centers for Disease Control and Prevention.2002. Outbreaks of gastroen-teritis associated with noroviruses on cruise ships—United States, 2002.
Morb. Mortal. Wkly. Rep.51:1112–1115.
11. Chang, K. O., Y. Kim, K. Y. Green, and L. J. Saif.2002. Cell-culture propagation of porcine enteric calicivirus mediated by intestinal contents is
dependent on the cyclic AMP signaling pathway. Virology304:302–310.
12. Clark, S. M., J. R. Roth, M. L. Clark, B. B. Barnett, and R. S. Spendlove.
1981. Trypsin enhancement of rotavirus infectivity: mechanism of
enhance-ment. J. Virol.39:816–822.
13. Crawford, S. E., S. K. Mukherjee, M. K. Estes, J. A. Lawton, A. L. Shaw, R. F. Ramig, and B. V. Prasad.2001. Trypsin cleavage stabilizes the rotavirus
VP4 spike. J. Virol.75:6052–6061.
14. Cubitt, W. D., K. Y. Green, and P. Payment.1998. Prevalence of antibodies to the Hawaii strain of human calicivirus as measured by a recombinant
protein based immunoassay. J. Med. Virol.54:135–139.
15. De Leon, R., S. M. Matsui, R. S. Baric, J. E. Herrmann, N. R. Blacklow, H. B. Greenberg, and M. D. Sobsey.1992. Detection of Norwalk virus in stool specimens by reverse transcriptase-polymerase chain reaction and
nonradio-active oligoprobes. J. Clin. Microbiol.30:3151–3157.
16. Dimitrov, D. H., S. A. Dashti, J. M. Ball, E. Bishbishi, K. Alsaeid, X. Jiang, and M. K. Estes.1997. Prevalence of antibodies to human caliciviruses (HuCVs) in Kuwait established by ELISA using baculovirus-expressed
cap-sid antigens representing two genogroups of HuCVs. J. Med. Virol.51:115–
118.
17. Fankhauser, R. L., S. S. Monroe, J. S. Noel, C. D. Humphrey, J. S. Bresee, U. D. Parashar, T. Ando, and R. I. Glass.2002. Epidemiologic and molecular trends of “Norwalk-like viruses” associated with outbreaks of gastroenteritis
in the United States. J. Infect. Dis.186:1–7.
18. Fankhauser, R. L., J. S. Noel, S. S. Monroe, T. Ando, and R. I. Glass.1998. Molecular epidemiology of “Norwalk-like viruses” in outbreaks of
gastroen-teritis in the United States. J. Infect. Dis.178:1571–1578.
19. Flynn, W. T., and L. J. Saif.1988. Serial propagation of porcine enteric calicivirus-like virus in primary porcine kidney cell cultures. J. Clin.
Micro-biol.26:206–212.
20. Gilpatrick, S. G., K. J. Schwab, M. K. Estes, and R. L. Atmar.2000. Devel-opment of an immunomagnetic capture reverse transcription-PCR assay for
the detection of Norwalk virus. J. Virol. Methods90:69–78.
21. Glass, P. J., L. J. White, J. M. Ball, I. Leparc-Goffart, M. E. Hardy, and M. K. Estes.2000. Norwalk virus open reading frame 3 encodes a minor
structural protein. J. Virol.74:6581–6591.
22. Glass, R. I., J. Noel, T. Ando, R. Fankhauser, G. Belliot, A. Mounts, U. D. Parashar, J. S. Bresee, and S. S. Monroe.2000. The epidemiology of enteric caliciviruses from humans: a reassessment using new diagnostics. J. Infect.
Dis.181(Suppl. 2):S254–S261.
23. Green, J., J. Vinje, C. I. Gallimore, M. Koopmans, A. Hale, D. W. Brown, J. C. Clegg, and J. Chamberlain.2000. Capsid protein diversity among
Norwalk-like viruses. Virus Genes.20:227–236.
24. Green, K. Y., A. Z. Kapikian, and R. M. Chanock.2001. Human caliciviruses,
p. 841–874.InD. M. Knipe, P. M. Howley, D. E. Griffin, et al. (ed.), Fields
virology, 4th ed. Lippincott-Raven, Philadelphia, Pa.
25. Guerrero, R. A., J. M. Ball, S. S. Krater, S. E. Pacheco, J. D. Clements, and M. K. Estes.2001. Recombinant norwalk virus-like particles administered intranasally to mice induce systemic and mucosal (fecal and vaginal) immune
responses. J. Virol.75:9713–9722.
26. Halstead, S. B.1988. Pathogenesis of dengue: challenges to molecular
biol-ogy. Science239:476–481.
27. Harrington, P. R., L. Lindesmith, B. Yount, C. L. Moe, and R. S. Baric.2002. Binding of Norwalk virus-like particles to ABH histo-blood group antigens is blocked by antisera from infected human volunteers or experimentally
vac-cinated mice. J. Virol.76:12335–12343.
28. Harrington, P. R., L. Lindesmith, B. Yount, C. L. Moe, J. LePendu, and R. S.
Baric.2003. Norovirus attachment, susceptibility, and vaccine design. Recent
Res. Dev. Virol.5:19–44.
29. Harrington, P. R., B. Yount, R. E. Johnston, N. Davis, C. Moe, and R. S. Baric.2002. Systemic, mucosal, and heterotypic immune induction in mice inoculated with Venezuelan equine encephalitis replicons expressing
Nor-walk virus-like particles. J. Virol.76:730–742.
30. Hennessy, E. P., A. D. Green, M. P. Connor, R. Darby, and P. MacDonald.
2003. Norwalk virus infection and disease is associated with ABO histo-blood
group type. J. Infect. Dis.188:176–177.
31. Huang, P., T. Farkas, S. Marionneau, W. Zhong, N. Ruvoen-Clouet, A. L. Morrow, M. Altaye, L. K. Pickering, D. S. Newburg, J. LePendu, and X. Jiang.2003. Noroviruses bind to human ABO, Lewis, and secretor histo-blood group antigens: identification of 4 distinct strain-specific patterns.
J. Infect. Dis.188:19–31.
32. Hutson, A. M., R. L. Atmar, D. Y. Graham, and M. K. Estes.2002. Norwalk virus infection and disease is associated with ABO histo-blood group type.
J. Infect. Dis.185:1335–1337.
33. Hutson, A. M., R. L. Atmar, D. M. Marcus, and M. K. Estes.2003. Norwalk virus- like particle hemagglutination by binding to h histo-blood group
an-tigens. J. Virol.77:405–415.
34. Jiang, X., M. Wang, D. Y. Graham, and M. K. Estes.1992. Expression, self-assembly, and antigenicity of the Norwalk virus capsid protein. J. Virol.
66:6527–6532.
35. Jiang, X., M. Wang, K. Wang, and M. K. Estes.1993. Sequence and genomic
organization of Norwalk virus. Virology195:51–61.
36. Jiang, X., N. Wilton, W. M. Zhong, T. Farkas, P. W. Huang, E. Barrett, M. Guerrero, G. Ruiz-Palacios, K. Y. Green, J. Green, A. D. Hale, M. K. Estes, L. K. Pickering, and D. O. Matson.2000. Diagnosis of human caliciviruses by
use of enzyme immunoassays. J. Infect. Dis.181(Suppl. 2):S349–S359.
37. Karst, S. M., C. E. Wobus, M. Lay, J. Davidson, and H. W. t. Virgin.2003.
STAT1- dependent innate immunity to a Norwalk-like virus. Science299:
1575–1578.
38. Koopmans, M., C. H. von Bonsdorff, J. Vinje, D. de Medici, and S. Monroe.
2002. Foodborne viruses. FEMS Microbiol Rev.26:187–205.
39. Lindesmith, L., C. Moe, S. Marionneau, N. Ruvoen, X. Jiang, L. Lindblad, P. Stewart, J. LePendu, and R. Baric.2003. Human susceptibility and
resis-tance to Norwalk virus infection. Nat. Med.9:548–553.
40. Lopman, B. A., G. K. Adak, M. H. Reacher, and D. W. Brown.2003. Two epidemiologic patterns of norovirus outbreaks: surveillance in England and
Wales, 1992–2000. Emerg. Infect. Dis.9:71–77.
41. Marionneau, S., A. Cailleau-Thomas, J. Rocher, B. Le Moullac-Vaidye, N. Ruvoen, M. Clement, and J. Le Pendu.2001. ABH and Lewis histo-blood group antigens, a model for the meaning of oligosaccharide diversity in the
face of a changing world. Biochimie83:565–573.
42. Marionneau, S., N. Ruvoen, B. Le Moullac-Vaidye, M. Clement, A. Cailleau-Thomas, G. Ruiz-Palacois, P. Huang, X. Jiang, and J. Le Pendu.2002. Norwalk virus binds to histo-blood group antigens present on
gastroduode-nal epithelial cells of secretor individuals. Gastroenterology122:1967–1977.
43. Mead, P. S., L. Slutsker, V. Dietz, L. F. McCaig, J. S. Bresee, C. Shapiro, P. M. Griffin, and R. V. Tauxe.1999. Food-related illness and death in the
United States. Emerg. Infect. Dis.5:607–625.
44. Morens, D. M.1994. Antibody-dependent enhancement of infection and the
pathogenesis of viral disease. Clin. Infect. Dis.19:500–512.
45. Parker, S. P., W. D. Cubitt, X. J. Jiang, and M. K. Estes.1994. Seropreva-lence studies using a recombinant Norwalk virus protein enzyme
immuno-assay. J. Med. Virol.42:146–150.
46. Parwani, A. V., W. T. Flynn, K. L. Gadfield, and L. J. Saif.1991. Serial propagation of porcine enteric calicivirus in a continuous cell line. Effect of medium supplementation with intestinal contents or enzymes. Arch. Virol.
120:115–122.
47. Ravn, V., and E. Dabelsteen.2000. Tissue distribution of histo-blood group
antigens. Apmis108:1–28.
48. Smiley, J. R., K. O. Chang, J. Hayes, J. Vinje, and L. J. Saif.2002. Charac-terization of an enteropathogenic bovine calicivirus representing a
poten-tially new calicivirus genus. J. Virol.76:10089–10098.
49. Sullivan, N. J.2001. Antibody-mediated enhancement of viral disease. Curr.
Top. Microbiol. Immunol.260:145–169.
50. Tamura, M., K. Natori, M. Kobayashi, T. Miyamura, and N. Takeda.2000. Interaction of recombinant norwalk virus particles with the 105-kilodalton cellular binding protein, a candidate receptor molecule for virus attachment.
J. Virol.74:11589–11597.
51. Vinje, J., J. Green, D. C. Lewis, C. I. Gallimore, D. W. Brown, and M. P. Koopmans.2000. Genetic polymorphism across regions of the three open
reading frames of “Norwalk-like viruses”. Arch. Virol.145:223–241.
52. Vinje, J., R. A. Hamidjaja, and M. D. Sobsey.2004. Development and application of a capsid VP1 (region D) based reverse transcription PCR assay for genotyping of genogroup I and II Noroviruses. J. Virol. Methods
116:109–117.
53. White, L. J., J. M. Ball, M. E. Hardy, T. N. Tanaka, N. Kitamoto, and M. K. Estes.1996. Attachment and entry of recombinant Norwalk virus capsids to
cultured human and animal cell lines. J. Virol.70:6589–6597.