Copyright© 1994, AmericanSocietyforMicrobiology
Sequence Requirements for Stable
Binding and Function
of
Rep68
on
the Adeno-Associated Virus
Type 2
Inverted Terminal Repeats
JOHNA. CHIORINI,l* STEPHENM.WIENER,' ROLAND A. OWENS,2 SIRKKA R. M. KYOSTIO,2 ROBERT M.KOTIN,73 ANDBRIANSAFER'
MolecularHematology Branch, NationalHeart, Lung, andBloodInstitute,1andLaboratory of Molecular and CellularBiology, NationalInstituteofDiabetesandDigestiveandKidney Diseases, 2
Bethesda, and GeneticTherapy, Inc., Gaithersburg, Maryland
Received 26May1994/Accepted 8 August1994
Replication of the palindromic inverted terminalrepeats (ITRs) of adeno-associated virus type 2 requires several functions of the viral nonstructural Rep proteins. These include bindingto theITR, nickingofthe double-stranded replication intermediate atthe terminal resolution site (trs), and then stranddisplacement andsynthesis from the nick. This report demonstrates the ability of both recombinant fusionmaltose-binding protein (MBP)-Rep68A produced in Escherichia coli and wild-type (wt) Rep68 to bindto alinear truncated form of the ITR, A57 ITR, with similar affinity as to the wt hairpin ITR. A dissociation constant for MBP-Rep68Aof approximately 8 x 10-10 Mwasdetermined for thewtITR and 457ITRprobes. Truncation of 457 ITRto generate 428ITR, which retains the GCTCrepeatmotif but not the trs, boundatleast 10 times less efficiently than 457 ITR. Extension of A28 ITR with nonspecific sequence restored the ability of MBP-Rep684 tobindtoA28 ITR. Thus, high-affinity binding wouldappeartorequire stabilization byflanking sequence aswellasthe intact GCTC repeatmotif.Cleavage of theA57ITRprobe with DdeI, which truncates the flankingsequence andwas previously shown to inhibit binding by Rep68, also inhibited the binding of MBP-Rep684. The requirements for stablebindingwerefurther defined withaseriesofoligonucleotide probes which spanned the region protected by MBP-Rep78 in DNase I footprinting. The binding activity ofeither MBP-Rep684 orwtRep68tohairpin ITRorA57ITRwasindistinguishable. However, the binding activity of MBP-Rep684 toDNA does not appeartocorrelate withtrs endonucleaseactivity. The nicking and covalent linkage ofMBP-Rep68Atothenonhairpin 457 ITRwasapproximately 100-fold less efficient than its linkage toahairpin-containing ITR. Therefore, although the hairpin portion of the ITR doesnotappeartoplayarole inrecognition and stabilization of MBP-Rep684 binding, itspresencedoes affect the trs cleavageactivity of the protein.
Adeno-associatedvirustype 2(AAV2)isahuman parvovi-ruswhich requires thepresence ofahelpervirus toreplicate efficiently.Thissingle-stranded DNA viruscontainstwoopen
readingframes (ORFs). The leftORFencodes the nonstruc-tural Rep proteins, which are involved in regulation of repli-cation and transcription in addition to the production of single-strandedprogenygenomes (6-8, 14, 32, 40, 42,45-47). Furthermore, two of the Rep proteins have been associated with the preferential integration of AAV genomes into a
regionoftheqarmof humanchromosome 19(21-23, 41,48). The right ORF encodes the capsid proteins, VP1, VP2, and VP3. ThesetwoORFs areflankedby145-nucleotide inverted terminalrepeats(ITRs)whichcanformhairpinstructuresthat
serve asprimers during replication (44). Thetwo larger Rep proteinstranscribed fromthep5promoter,Rep68andRep78,
are referred to by their apparent masses as determined by sodium dodecyl sulfate (SDS)-polyacrylamide gel electro-phoresis.Theseproteins have been showntobindtoITRs(1, 8, 16-18, 35-37, 44) and stimulate replication bothin vivo(25, 26, 45)and invitro (8, 15,33). Furthermore,Rep68 and Rep78 have been shown to nick the duplex ITR in a site- and strand-specific manner. This cleavage, which occurs at the
*Correspondingauthor.Mailingaddress:NIH/NHLBI/MHB, Bldg.
10/7D18, 10 Center Dr.,MSC 1654, Bethesda, MD 20892-1654.Phone: (301)496-1594.Fax: (301) 496-9985.
terminal resolution site (trs), permits the replication of the hairpinstructureinaprocessreferredtoasterminal resolution (42).
In addition to their role in replication, Rep68 and Rep78 have been shown invivo to regulate the transcriptionofthe AAV p5,
p19,
and p40 promoters (2, 24, 27, 30, 46). The p5-initiated Rep proteinscan also inhibit the transcriptionofheterologous promoters aswell ascellular transformation by papillomavirus or by adenovirus Ela plus an activated ras oncogene(11-13, 20, 25, 26). Rep68andRep78wererecently showntobindtoaregiononhumanchromosome 19 that had been identifiedas a preferredsite for AAV DNAintegration (21-23, 48). Furthermore, Rep68 was shown to mediate an interaction betweenthe humanintegrationlocus and the ITR of AAV, suggesting a role for this protein in the targeted integrationof AAV(48).
Previous literature hassuggestedthat in order forRep68to bindtotheITR,it hadtobe in ahairpinform(1, 16).Recent worksupportsthisideabydemonstratingthatanITR cleaved withDdeI, which removes the cross armsof the ITRhairpin and seven bases of the stem, would not bind Rep68 (43). Therefore,therecentobservationsthatRep68andRep78will bind to a linear fragment from human chromosome 19 (48) andarecombinantRep68canbind toafull-length ITRstem (8) (seeFig.1B)appeartobecontradictorytotherequirement forahairpinstructureforbinding. However,other studiesby Berns and colleagues have shown the importance of the
7448
on November 9, 2019 by guest
http://jvi.asm.org/
A. wt ITR A C
c
bA
5'AGGAACCCCTAGTGATGGAG-'tGGCCACTCCCTCTCTGCCCGCCCCACTGAGGCC
TCCTTGGGGATCACTACCTCAACGGTGAGGGAGAGACGCGCGAGClGAllGllGllAllGllT
TCCTTGGGGATCACTACCTCAACCGGTGAGGGiAGAGACGCGCGAGCGAGCGAGTGACTCCGG
D A'
B
B. A57 ITR
c c c G c T G G T
5'
eA
G-C G-C G-C
T
CG-CG GC-GC GC-GC GC-GC
T
GATCAGTGATGGAG1TGGCCACTCCCTCcTGCGcGcCYGCEAC'GAGGCC
TCACTACCTCAACCGGTGAGGGAGAGACGCGCGAGCGAGCGAGTGACTCCGGCTAG
D A'
A
C. A28 ITR TCTGCG#GCACGGAGGC(
AGACGCGCGAGCGAGCGAGTGACTCCG
A'
A
TGCtCE?CF?GCCA
D. Al8 ITR ...
ACGCGCGAGCGAGCGAGT
A'
E. ITR T
c A
A A G C _G G -C
c G-C C
G-C C -G C -G C -G G-C
G
c C -G C -G C -G G -C n C -G n
T -A G -C G -C T T
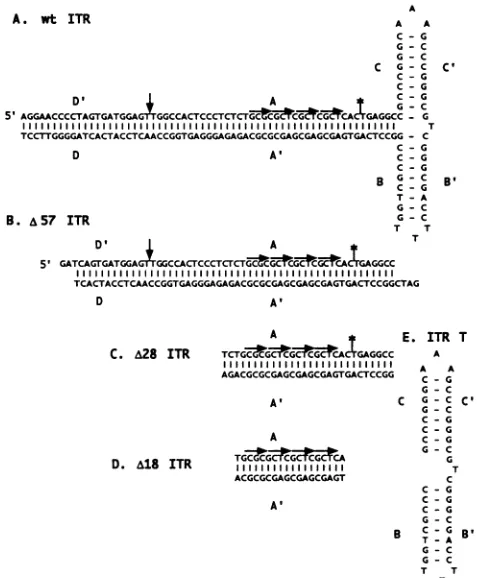
FIG. 1. Sequencesofprobes.wtITR(A)was madeaspreviously described(1). A57 ITR(B)is anannealedoligonucleotide pairof 57 bases inlength correspondingtoall of theA/A' andpartof theD/D' regionofwt ITR with a GATC 5' overhang. Also shown are three further truncations of theITR,A28 ITR(C), A18ITR(D),andITR T(E). Sequences are alignedtotheGCTC imperfectrepeats (hori-zontalarrows). Thetrs is indicated with avertical arrow. TheDdeI
cleavagesite in thestemof thewtITR is indicatedbyanasterisk.
hairpin structure for origin function in vivo (4, 5, 29, 39). Mutations which maintained the structure were shown to retainorigin activity.
Characterization ofthemechanismof action ofRep68and Rep78 has been limited by the inability to produce these proteins in large quantities. We have recently succeeded in expressing Rep68A and Rep78 as fusion proteins with the maltose-binding protein (MBP) in Escherichia coli and have demonstrated that the recombinant MBP-Rep68A can
per-form all of the in vitro activities described for thewild-type (wt) Rep protein (8). In this study, the DNA sequence and structuralrequirementsforMBP-Rep68A bindingare charac-terized. Thisreportdemonstrates thatMBP-Rep68Aisable to bind to a linear truncated form of the ITR, A57 ITR, with similaraffinityasto wthairpinITR.Furthermore, bindingtoa linear ITR is afunction notonly ofMBP-Rep68A but of wt Rep68 as well. Similar specific DNAbinding activities were obtained for MBP-Rep68A and Rep68. However, while the hairpin is not required for Rep binding to the ITR, it does
appeartohaveastimulatoryeffectontrsendonucleaseactivity by Rep proteins in vitro.
MATERIALS AND METHODS
Probes. DNAprobes aresummarizedinFig. 1, 6, and 7A. Oligonucleotide probes were synthesized with a model 392 DNA/RNA synthesizer (Applied Biosystems).The
oligonucle-otides usedto
produce
theA57
ITRwerepurified
ondenatur-ing polyacrylamide gels.
Allotheroligonucleotides
werepuri-fied
by
NAP25gel
filtrationchromatography
(Pharmacia).
5
'-32P-labeled
wtITRprobe
wasproduced
by XbaI digestion
ofpSub2O1 (38). Following
calf intestinalalkalinephosphatase
treatment,the 5'terminuswaslabeledwith
[-y-32P]ATP
(3,000
Ci/mmol) by polynucleotide kinase,
and the labeledfragment
was released
by
PvuIIdigestion.
The linearprobe
was then converted to the wt ITR conformationby
boiling, rapid
chilling,
andfilling
in the 5'overhang
with Klenowfragment
andcold
deoxynucleoside triphosphates.
The unlabeledcom-plementary
strand releasedby
Xba-PvuIIdigestion
cannotbe extendedby
this treatment.Therefore,
thewt5'-labeled ITR canbeseparated
from the shortercomplementary
strandby
electrophoresis
on a 6% Tris-borate-EDTAacrylamide gel,
excised from the
gel
and eluted into lx Tris-EDTA buffer. 3'-labeled wt ITR wasprepared by boiling
andquickly
cooling
an unlabeledXbaI-PvuIIfragment
and thenfilling
in the 5'overhang
withKlenowfragment
and[a-32P]dCTP.
To obtain 3'-labeled ITRprobe
ofthesamespecific activity
asA57 ITR, unlabeled dCTP was added to thelabeling
cocktail to compensatefor the greaternumber ofcytosine
residues in the 3'-labeledprobe.
The A ITR
probes
were madeby
5' endlabeling
eitheroligonucleotide
andannealing
with theunlabeledcomplemen-tary
oligonucleotide.
A57 ITRwas 3' endlabeledby
filling
in the5'overhang
createdby
theBamHIrestriction site included in theoligonucleotides.
DdeI truncation of A57 ITR wasgenerated by
cleavage
of5'-end-labeled A57 ITRprobe
with DdeI restriction enzyme and shown to be greater than 90% cleavedby
analysis
on adenaturing acrylamide gel.
Preparation
ofwtRep68
from transfected 293 cells. Thepreparation
ofwtRep68
has been describedpreviously
(37).
Briefly,
aplasmid
expressing Rep68
fromthe humanimmuno-deficiency
viruslong
terminal repeat(pSK9)
was transfected into human293cells,
and the cellswereharvested 48 hlater;
600 mM NaCl nuclear extracts were
prepared
as describedpreviously
(37).
Asnegative controls,
similar nuclear extracts weremadefrom 293 cells transfected withaplasmid encoding
anonfunctionaltruncationmutantform ofRep68
(Repam238)
(37)
orpBR322.
MBP-Rep68A
andMBP-Rep78.
Theconstruction,
expres-sion,
andpurification
of theseproteins
have been describedpreviously
(8).
Briefly,
PCR was used toamplify
theRep68
ORF from AAV nucleotides 327 to 1882. This
fragment
wasthen clonedinto
plasmid pPR997
(New England
Biolabs).
Thefull-length
Rep78
gene(nucleotides
327to2186)
was cloned into the same vector toproduce
MBP-Rep78.
The fusionproteins
werepurified by
amylose
affinity chromatography.
The elutedMBP-Rep68A
andMBP-Rep78
protein
fractionswerefoundtoconsist of90%
full-length
molecules and contain a minor fraction ofprematurely
truncatedMBP-Rep68A
andMBP-Rep78,
as determinedby
Coomassie bluestaining.
Theprematurely
truncatedfragments
were shown to lacksignifi-cant DNA
binding activity
by
theirseparation
from thefull-length molecules,
using
ITR-derived DNAaffinity
columnchromatography
(data
notshown).
Immunoblotting.
Westernimmunoblotanalyses
ofwtRep68
and
MBP-Rep68A
wereperformed
as described elsewhere(24). Briefly,
serial dilutions ofRep68-containing
293 cell nuclear extracts oramylose
column-purified
MBP-Rep68A
samples
wereelectrophoresed
on a12.5%polyacrylamide gel
containing
0.1% SDS. Theproteins
wereelectroblotted(Mul-tiphor
II NovaBlotelectrophoretic
transferapparatus; Phar-macia LKBBiotechnology,
Piscataway,
N.J.)
ontopolyvinyli-dene difluoride membranes
(Millipore
Corp.,
Bedford,
Mass.).
t
--f
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.55.294.73.362.2]The primary antibody was a rabbit antibody (anti-Rep78.93) raisedagainstatruncatedRep78expressedin E. coli (47).The secondary antibodywasbiotinylated goatanti-rabbit immuno-globulin G. The blots were then treated with streptavidin-alkaline phosphatase and incubated with achemiluminescent alkaline phosphatase substrate (PhotoBlot chemiluminescent systemforprotein
detection;
LifeTechnologies Inc.,Gaithers-burg,
Md.)
andexposed
toX-ray film. Protein concentrations intheoriginal
samplesweredeterminedby usingthe bicincho-ninic acidprotein
assayreagent(Pierce,
Rockford, Ill.)accord-ing
to the manufacturer's instructions. The amount ofRep68protein
contained in eachsample
wasmeasuredby densitom-etry of thefull-length Rep protein band.Electrophoretic mobility shift assays (EMSAs). DNA-pro-tein complexes were detected by their reduced mobility on nondenaturing polyacrylamide gels. The assays were per-formedaspreviously described (9, 10),with themodifications described
by
Jacob et al.(19). Briefly,
radiolabeled probeswere incubated withproteinfractions at30°Cfor 15 min in 25
RI.
The reaction mixture contained 10mMTris-Cl(pH
7.5), 1 mM EDTA, 10 mMP-mercaptoethanol,
0.1% Triton X-100, 4%glycerol,
and 0.5pg
ofpoly(dI-dC)
whenindicated.Specific activity
determination. The specific activities ofMBP-Rep68A
andwt Rep68 produced from 293 cells trans-fected withpSK9
were determinedasfollows. The amountof shiftcomplex
formed in Fig. 2A for MBP-Rep68A and wtRep68
was determined byPhosphorlmager
(MolecularDy-namics)
analysis usinga5'-end-labeledprobe.The shift bandsproduced
byMBP-Rep68A
per 100 ngofproteinwerefourfold greater than the complexes formed by 170 ng ofwt Rep68-containingcellextract.The ratio ofDNA toproteinwasshown in otherexperiments
to be within the linear range, and this difference inbinding activitywasdeterminedatseveralDNA/protein
ratios(Fig.
4C). Thus,MBP-Rep68A
hadapproxi-mately
seventimesasmuchgel shifting activitypermicrogram of totalprotein
as the 293 nuclear extract did. However,quantitation
ofthefull-length Rep68 proteinscontained inthe twoextractsby densitometryofaWesternblot(Fig.3, lanes 3 to 5 and 7 to9)
indicated thatMBP-RepA
contained 8to 16 times more Rep68 protein molecules per microgram of totalprotein.
Therefore, thespecific
DNAbindingactivities of theRep68 proteins
inthepreparationsofMBP-Rep68A
andpSK9 extractswerewithintwofold of each other.Dissociation constant determination. The Kd was deter-minedby measuringtheamountofDNA-MBP-Rep68A
com-plex
formationas afunction ofincreasingDNAconcentration,using
aPhosphorlmager.
One hundred nanograms ofamylose-purified
MBP-Rep68A
was incubated in an EMSA within-creasing
amountsof 5'-end-labeledwtITR orA57ITR(60to 400fmol).
Thespecific
activityof eachprobewasdeterminedby separating
the labeled probe from free32P-nucleoside
triphosphate by
gel filtration followed by quantitation vialiquid
scintillationcounting.wtITRand A57ITRprobes were found tohave thesamespecific activityof2 x 105 cpm/pmol. For the purpose of conversion from PhosphorImager units(PIUs)
tocountsperminute,standardswereincluded on eachgel.
The conversion factors are 46 PIU/cpm of probe forA57 ITR and 38 PIU/cpm ofprobefor wt ITR. The difference in PIUfor thetwoprobesistheresultof differentexposure timeson the
Phosphorlmager
for the experiments. Eadie-Hofsteeanalysis
ofthe data (boundversusbound/free) resulted in astraight
line(slope
= -1.8 x 105PIUfor A57ITRand -1.87 X 105 PIU forwt ITR) and thus a single value for the slope,-Kd,
where Kd is the dissociation constant (Fig. 4). As a confirmation of the value obtainedby this method, a second determination was madeby the method outlined byMeister-ernstet al. (31). The twomethods generatedKd values in the same range.
DNase I footprinting. The substrates used for DNase I protection experimentsconsisted of the synthetic oligonucleo-tideA57ITR,which contains the AD' orA'D sequence of the ITRshown in Fig. 1B. The oligonucleotides were individually 5' end labeled and annealed as described above. The condi-tions for DNase I footprinting were as described previously (16), with thefollowing modifications. About 5 X
104
cpm of probe wasusedwith 0.5,ugofpoly(dI-dC) and 2,ugofprotein. The digestion time was decreased to30 s. The reactions were terminated in 2 volumes of stop buffer (10 mM Tris-Cl[pH
7.9], 10 mM NaCl,0.5% SDS, 0.2 mg of yeast tRNA perml, 2 mg of proteinase K per ml). The samples were extracted by phenol-chloroform and ethanol precipitated. The products were then resuspended in 80% formamide loading buffer, denatured by heating to95°Cfor 5 min, and then fractionated on a 6% polyacrylamide-8 M urea sequencing gel.
Covalent linkage trs endonuclease assay. Activity was as-sayedby performing the trsendonuclease assay with 3'-labeled probes as previously described (8, 17, 18), with the following modifications. After incubation of 100 ng of MBP-Rep68A with either wt ITR orA57 ITR (50 or 500 fmol,respectively), the reactions wereterminated by the addition of 1 volume of 2x sample buffer (120 mM Tris-HCl [pH 6.8], 10% SDS, 2% dithiothreitol, 40% sucrose), and the samples were heated to 95°C for 5 min. The 3'-radiolabeled probe covalently boundto MBP-Rep68A was separated from the free probe by electro-phoresis on anSDS-10% polyacrylamide gel.
RESULTS
MBP-Rep68A and wt Rep68 bind to A57 ITR. Previous work has shown that the recombinantMBP-Rep68A is able to bind to a linear form of the AAV ITR (8). A57 ITR is the entire AAV ITRA/A' duplex region plus 10 bp of sequence down-stream of the trs (D/D') (Fig. 1B). When MBP-Rep68A and 293 cell nuclear extracts containing wt Rep68 were incubated with the A57 ITR probe, a cluster of three reduced mobility bands was detected (Fig. 2A, lanes 2 and 3, respectively). In contrast, these bands are not obtained with a non-DNA-binding truncation mutant of Rep68, Repam238 (lane4), or in thepresence of a 10-fold excess of unlabeled competitor (lanes 5 and6). The difference in mobility of theMBP-Rep68A and the wt Rep68-DNA complexes is the result of a difference in the molecular masses of the two proteins (106 and 68 kDa, respectively). Some additional bands that are not produced withMBP-Rep68A are present when theA57 ITR is incubated with 293 cellnuclear extract containing wt Rep68. However, thesebands are produced when the Repam238 mutant is used and are thought to be theresult of cellular proteins presentin the 293 nuclear extract.
Therefore, the ability of eitherMBP-Rep68A or wt Rep68 to bind to the ITRdoes not require the presence of the hairpin cross arm (B/B' C/C'). Furthermore, binding to aA57 ITR is a function not only ofMBP-Rep68A but of wt Rep68 as well, and no binding is detected with the MBP moiety fused to LacZ (datanot shown).
The ability ofRep68 to bind to a linear form of the ITR,A57 ITR, directly contradicts several previousobservations (1, 16, 43). To resolvethese discrepancies, we truncated theA57 ITR at theDdeI site, which was previously shown to inhibit binding to the wt ITR (43). Truncation at this position significantly reduced binding of MBP-Rep68A compared with the full-length A57 ITR probe (Fig. 2B, lanes 1 and 2). Competition experiments indicated that theDdeI-truncated fragment bound
on November 9, 2019 by guest
http://jvi.asm.org/
A28 ITR Competitor
*_
C.6nuDu
o co CO CZ)
°~ mc F cmYc0c
o co co
co aco<
29t co
N N
a-c cs
3 Cells
1 F E. coil N
-j
Rep68
CLMBP-Rep6BA
E.~.
-.gp; .-N -MBP-Rep68A
Bound ~ ~ ..
A57 ITR
...
Free A57 ITR
1 2 3 4 5 6 7
a) 57 Dde
i
-Bound
-Free
1 2 3 4 5 6 7 8
FIG. 2. BothwtRep68 andrecombinantMBP-Rep68AbindtoA57 ITR.(A)Approximately 10,000cpmof5'-end-labeled A57 ITRprobe wasincubatedwith either 100 ngofMBP-Rep68A or 170ng ofwt
Rep68nuclearextractproteinorRepam238nuclearextractprotein. The positionsof the free probe and boundprotein-DNA complexes areindicated atthe left. Rep68-DNA complexeswere detected only withMBP-Rep68A&andwtRep68 producedfromplasmid pSK9(pSK9 Rep 68; lanes2and3, respectively). Nospecificbindingwasdetected bythe negative controls (probe only [lane 1],MBP-LacZ [data not
shown], anda truncated ambermutation, Repam238 [lane 4]). The MBP-Rep68AandwtRep68shiftbands could becompetedforbythe addition ofa10-foldexcessofunlabeled A28 ITRcompetitor (lanes5
and 6). (B) 5'-end-labeled A57 ITRprobe was cut with DdeI and showntobe greaterthan 90%digested by separationonadenaturing gel. One hundred nanograms ofMBP-Rep68A was incubated with approximately0.1ngof either DdeI(lane 1)orA57 ITRprobe (lane 2) under standard EMSA conditions. The reaction mixtures were incubatedatroomtemperature,and thegelwasrunatroom temper-atureasreportedpreviously (43).Forcompetition,a5-, 10-,or15-fold
excessof unlabeledduplex oligonucleotideswasaddedtothe reaction
mixture with labeled A57 ITRprobe(lanes3to8).
_10W A, - --*- Rep68
1 2 3 4 5 6 7 8 9
FIG. 3. Westernblot comparison ofwtRep68- and MBP-Rep68A-containing extracts. The amounts of Rep68 protein contained in pSK9-transfected293 cell nuclearextractandamylose column-purified MBP-Rep68Awerecomparedby immunoblotting using anti-Rep78.93 antibody(47). Three negative controlswereincluded:4,ug of nuclear
extract protein from293 cells transfected with pBR322 (lane 1) or pHIVRepam (Repam238; lane 2) and 4 ,ug of amylose column-purified MBP-LacZ produced in E. coli (lane 6). Lanes 3 to 5 contained4, 2,and 1 ,ug, respectively, of pSK9 Rep68nuclearextract. Lanes 7to9contained 250, 125, and 62ng,respectively, of amylose column-purified MBP-Rep68A. The positions ofpSK9-produced wt
Rep68(Rep68) and MBP-Rep68Aareindicatedatthe right.
MBP-Rep68A at least 10 times less efficiently than the full-length A57ITR(lanes 3to8).This result is inagreementwith the other report of Rep68 binding activity (43). However, bindingisobservedonthefull-lengthA57 ITR(lane 2),which indicates theimportance of this regionin the formation of a stable complex and not necessarily the requirement of a
hairpin conformation for stable binding as previously
sug-gested.
Determination of the DNA binding activities of MBP-Rep68AandwtRep68.Tocomparethe DNAbindingactivities ofMBP-Rep68A andwtRep68,the ITRbinding activitywas normalized totheamountofRep68 proteinasdeterminedby Western analysis. MBP-Rep68A bound approximately seven times more DNA per microgram of total protein than wt
Rep68(Fig. 2A,lanes 2 and3)when valueswerecorrectedfor the amount ofprotein used in each lane. The DNA/protein ratiowas shown in other experiments tobe within the linear
range of detection. This difference in binding activity was determined atseveralDNA/proteinratios and is shownagain inFig.4Aand B.Westernblotanalysiswithanti-Repantisera showed that MBP-Rep68A contained approximately 8 to 16 timesmorefull-length Rep68than 293 cellextractsdid(Fig. 3, lanes 3 to5 and 7to9). Therefore,thespecific activity of the Rep68 proteinsinthepreparationsofMBP-Rep68AandpSK9 extracts are within twofold of each other, indicating that MBP-Rep68AandwtRep68haveasimilar abilitiestobindthe AAVITR.
Since comparable binding activities to A57 ITR were ob-tained forwtRep68andMBP-Rep68A,the relativeaffinities of these protein preparations for hairpin ITR, A57 ITR, or a further truncation of A57 ITR, A28 ITR (Fig. 1),were
com-pared. The binding affinities ofwtRep68 and MBP-Rep68A
wereverysimilarfor thewtITRand A57 ITRprobes(Fig.4A
and B, lanes 1 to 5 and 6 to 10, respectively) but were
significantlyreducedupon further truncation of the A57 ITR probeto A28 ITR(lanes 11 to 15).
The data presented in Fig. 4A and B were obtained at a
A
.... MM'M
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.331.548.76.224.2] [image:4.612.101.260.92.516.2]A. wtITR A57ITR A28 ITR
Bound F MBP-6 RepB8L
B.
wtITR A57ITR A28 ITRBound _ pSK9 Rep68 L
wtITR-
"
_w
.,z. d
A57 ITR
-W < A28ITR
1 2 3 4 5 6 7 8 9 10 11 12131415 1 2 3
C. *wa_ W.W_ lowVW
BE ,.4 K.Sj j i [ "i
F-
_S
4 8 12 16 20 30 40 50 60 78 80 90 110 LINEAR ITR kcpm
D
700000-600000
500000
bound
(PIU) 400000
300000
200000 U w
* a
free (10 } (PIU)
* 0 E 80
70- 60-50
-E 40. E
30
-20
-rITR
ITD
~~10
-2
k
ItL
-I ' I a
4 5 6 7 8 9 10 11 12 13 1415
0
0
*_
*_
* ITR * AITR
B/F
wt ITR--
w
I.:.-,
X57 ITR
-A28ITR
-tw
IIIH
on November 9, 2019 by guest
http://jvi.asm.org/
FIG. 4. wt Rep68 and MBP-Rep68A bind to wt ITR and AITR with similar affinities. The relative binding affinities of recombinant MBP-Rep68A (A) andwtRep68(B)werecompared for three probes, wt ITR,A57ITR, andA28ITR. Increasing amounts of probe (10, 20, 30, 40,and 50 fmol)wereaddedtoa constant amountofprotein (100 ng).Lanes 1 to5 inbothpanelscontain wt ITR as the probe; lanes 6 to 10 contain A57ITR;lanes 11 to 15 containA28ITR.(C) To determine the Kd for the different protein-DNA complexes, the amount of bound and free probe wasmeasured over a range of amounts from 60 to 400 fmol with a constant amount of protein. Bound (B) and free (F) probe bands were quantitated withaPhosphorlmager (MolecularDynamics), and the data were used to determine the dissociation constants for each of the probes withMBP-Rep68A.The dots at theright were used to develop a standard curve for converting from PIU to counts per minute. Graphs with plots of boundversusfree probes(A57ITR orwtITR) presentedin PIU(D)and bound versusbound/free(B/F) (Eadie-Hofstee plot; E) are shown.
relatively lowRep/probe ratio; hence, the amount of complex that could be formed was limited despite the addition of increasing amounts of probe(e.g., lanes 6 to 10). While these results demonstratethe similar affinities of MBP-Rep68A and wtRep68for both the wt ITR and A57 ITR probes, the data do not allow for more precise Kd determinations. The binding experimentswerethereforerepeatedover ahigherRep/probe ratio.Figure 4Cshows thatas afixed amount of Rep is titrated with increasing amounts of A57 ITR probe, the amount of complex formed increases. Similar results were obtained with bothwtRepandMBP-Rep68A.
The affinities ofwtRep68 and MBP-Rep68A for wt ITR and A57 ITR were determinedby measuring the amount ofbound and free probe over a range of protein/DNA ratios (see MaterialsandMethods), anda curvewasfitted tothese values (Fig. 4D). Kdvalues for MBP-Rep68A of approximately 8 x
10`1
and 9 x1010
Mfor A57 ITRandwtITR,respectively, weredetermined byusing an Eadie-Hofstee plot(Fig. 4E).The difference in the yintercepts (Fig. 4D) mayresult from small variations in the amount of protein added to the reaction mixtures. This is in agreementwith the shift in thecurveinFig. 4D.A.
- Control Rep78 Lac Z
A M.
CTI
C_
1 2 3 4 5 6 7 8 9 10 11
Binding
affinity
ofMBP-Rep68A for a (GCTC)4 imperfect repeat motif is increased by nonspecific flanking sequence. MBP-Rep68AandwtRep68bind toA57ITRwith equivalent affinities. Further truncation of A57 ITR to A28 ITR or cleavage with DdeI resulted in asignificantdecrease in binding affinity (Fig. 4A and B, lanes 11 to 15; Fig. 2B, lane 1). To determine the basis for thereduced binding affinity, DNaseI protection analysiswasperformed with MBP-Rep78 and A57 ITR (Fig. 5). On the trs-containing strand of A57 ITR, protection extends over a 24-nucleotide sequence that is centered over an imperfect (GCTC)4 repeat (lanes 5 to 7). Truncation with DdeI cleaves within thisprotectedregion [5'(C/T)GAG]
andfurther supports theimportance ofthisregion in complex formation. Protection of the non-trs-containing strandcoversthecomplementof theGCTCrepeatmotif,but a total of 32 nucleotides are protected and the repeats are foundatthe 5' endoftheprotectedregion(lanes13to15).No protection was found on the trs region of either strand. To determine the minimal DNA sequence that would support the stablebinding ofMBP-Rep68A,severalduplexoligonucleotide probes spanning the GCTC repeat region were tested for binding activityand fortheabilitytocompetewithA57ITRforB.
- Rep 78 Lac Z Control
Gn: 11 11
~---
I U)A G
G o
A A
A
.MaX'1
G~~~~~
G :d.*
ClG
i
12 1314 15 16 17 18 19 20 21 22
C.
D' A
i~~ -" _ b
AGGAACCCCTAGTGATGGAGTTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCC TCCTTGGGGATCACTACCTCAACCGGTGAGGGAGAGACGCGCGAGCGAGCGAGTGACTCCGG
D A'
FIG. 5. DNase I footprintingofA57ITRwithMBP-Rep78protectsa GCTCrepeatin theA/A' regionoftheITR.The regionofcontact
betweenRepproteinand DNAwasidentifiedbyusingaDNase Iprotectionassay. Thecoding(trs-containing) (A)ornoncoding (B)strand of the5'-end-labeled A57 ITRwasincubatedwith 2 ,ug ofMBP-Rep78followedbyincreasingamountsofDNase I(lanes5to7and13to15).As acontrol,
A57
ITRwasincubated with DNaseIonly (lanes2to4and19 to21)orwithMBP-LacZ(lanes8to10and 16to18).Lanes1, 11,12,and 22areG+Aladders. (C)Schematicrepresentationof theprotectedregionsonthecodingandnoncodingstrands,withhorizontallinesover theprotected regionsonthecodingandnoncodingstrands. The GCTC repeat motif isindicatedbyfour horizontalarrows.Thetrs is indicated byaverticalarrow.
7m
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.144.468.418.662.2]BOUND COMPETE
ATCCGCCGCTCGrCTCT
GCGCGCTCGCTCGCT+
A
- ND
++ ++
A46 ITR
ND
FIG. 6. Minimum binding site determination for MBP-Rep68A. To determine the minimum DNAsequencerequired for binding, several probes which spanned the protectedareafoundby DNase I footprint-ingweresynthesized. Only the trs-containing strandsareshown. The GCTCimperfectrepeatregion in each probeisoutlinedbyabox. The
trsis indicated byaverticalarrow.
A
46M ITR
D' A
.AT'GGAGTTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGC
li l1 1 1 1 1 1IJi:1 1 1 llll!lllllllllllll|I
,C CAAC GTGAbbAGAGACGCCGAGC GAGCGATGACTCCG
D A'
D0' IA
W A
GATGGAGTrGGC(CACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGC ITACTCACCGGT!AGG!I!A!II II iCIGIIGCIaGGII
CTACCTCAC(CICGGAGbGGGAGACGCGCGAGCGAGCGAGTGACTCCG
A
binding (Fig. 6). Only the trs-containing strandsequences are
shown. All of the oligonucleotide probes tested had signifi-cantly weaker bindingactivity comparedwith A57 ITR. Oligo-nucleotideprobes thatcontained both therepeatsand flanking
sequences wereabletobindMBP-Rep68Aaswellascompete for MBP-Rep68A. Binding activity was proportional to the length of flankingsequencecontained in the probe: the fewer
the number of flanking nucleotides, the weaker the binding activity. Oligonucleotides1and6,whichcontainedatleast 3 bp offlanking sequence onbothsides ofthe imperfect (GCTC)4 repeat, hadthe strongest binding activity. As the number of flanking nucleotides was reduced, activity was decreased.
Truncation of therepeat motif (Fig. 6, 2, and 5) resulted ina total loss ofbinding activity.
These resultssupport the DNase Iprotection assayresults,
which showed protection extending beyond the immediate GCTC repeat region. To determine the extent of flanking
sequence needed to obtain comparable binding activityto wt ITR,A57ITRwasfurther truncatedtoA46 ITR (Fig. 7A).No loss of binding activitywasobserved with this probe (datanot
shown).Toinvestigate the possibility that interaction with the trswasrequired for stable MBP-Rep68A binding, a thymine-to-guanine transversionwasmadeatthetrsof A46MITR (Fig. 7A). Although trs endonuclease activitywas abolished (data notshown), binding ofMBP-Rep68Awasnotaffected (Fig. 7B, lanes 7 to 10). This finding is in agreement with recent
published observations (43). Binding activity of a A28 ITR
probe extended toward the trs with additional base pairs of randomsequence (A46RITR) (Fig. 7A) restored the binding activitytothelevel obtainedwith A57 ITR(Fig. 7B, lanes 12to 15). Thus, the increased binding affinity ofMBP-Rep68A for A57ITRcompared with A28 ITR is the result of nonspecific
sequence stabilization of the complex by flanking DNA se-quences.
Binding affinityofMBP-Rep68AtoDNA isnotproportional to trs endonuclease activity. The datapresented above
dem-onstratethatMBP-Rep68A andwtRep68are abletobindto
GCTCimperfectrepeatswithin linearduplex substrates in the A/A' region of the ITRwithan affinitysimilar to that of the hairpin ITR. Since the hairpin structureisrequired for DNA replication (4, 5, 28, 29, 39),the effect of the hairpinsequence
ontrs endonuclease activitywasexamined. The amount of 5' covalentlyattached MBP-Rep68A-DNA that results from the ATP,Mg2+-dependentendonuclease activitywasmeasured by
A46R
ITR
D' A
5 t
CaGATGTCCATGGGCAGTCTCTCTGCGCGCTCGCTCGCTCACTGAGGC
GIGI Gi IIIA IiGIGIAIGIAilGlIGA CIi
CGThCAGGTA C C CTCAGAGA GACSCG6CGAG CGA GCCAGGA CTC C
D B
A'
A 46 TR A46M ITR 546R ITR
MBPRep
-
Bound--F
Free
-"0.,.bomb " . - t .Iftwit - - - -,.'o46.6 as L .
I I k.-A
"W
Iw w
w .v n
:
[image:7.612.61.301.78.205.2]1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
FIG. 7. MBP-Rep68 binding is stabilized by nonspecific flanking
sequence, and trs is not required for stable binding. (A) Three
oligonucleotide probesweremadetoaddress thedifferencesinbinding affinitybetween A57 ITR andA28 ITR. A46M ITR is identicaltothe
wt sequence except for a point mutation at the trs site (T-to-G
substitution; highlightedandunderlined).A46R ITR isthesameprobe asA28ITRexceptthat it is extended withrandom DNAtobe thesame lengthas A46ITR.The GCTCrepeatsareindicated withhorizontal
arrows.ThetrsisindicatedfortheA46 ITR and A46M ITRprobes,
and itspotentialsite in theA46R ITRprobeisindicatedbyavertical
arrow. (B) Lanes 1, 6, and 11 contain no protein. All other lanes
containaconstant amountofMBP-Rep68A and increasingamounts
(10, 20, 30,and 40fmol)of A46 ITR(lanes2to5),A46M ITR(lanes 7 to 10), and A46R ITR(lanes 12to 15) probes. All three probes bound MBP-Rep68A with the same affinity, indicating that the
in-creaseinbinding affinityis the result ofincreasedprobe lengthandnot
specific protein-DNAinteractions.
2
3
4
6
+++ S+F
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.319.558.92.553.2]A MgCI2 -
-+-ATP - +
-MBP-Rep 68 - + + + +
Covalently Linked- 4
Rep:wtITR
Free wt
ITR-B wtITR.157ITR
Linked- 4
Free-1 2 345
FIG. 8. Covalent linkage ofMBP-Rep68Ato wtITRandA57ITR.
The standardtrsendonucleaseassays wereperformed asdescribedby
Im and Muzyczka (17), with the following modifications. After the incubationof 100ngofprotein with 50 fmol of3'-labeledwtITR intrs buffer,SDS sample bufferwasadded, the samplewasheated, and the
probecovalently boundto MPB-Rep68Awasseparated from the free probe byelectrophoresis on anSDS-10%polyacrylamide gel. (A) To
confirm the specificityof thetrsendonuclease assay,the dependence ofwtITRretarded mobilityonATP,MgCl2, andMPB-Rep68A was
examined. Formation of the ITR-MPB-Rep68A retarded complex is dependent onATP, MgCl2, and MBP-Rep68A. (B) By comparing the
amounts of free probeversusthat covalently linked to the MBP-Rep
protein, the efficiency with which the two probeswere cleaved was
determined. The probes used in thisexperimentwere3' labeledtothe
samespecific activity of 2x105 cpm/pmol.Lane 1containswtITRas
a substrate for MBP-Rep68A, and lane 2 contains A57 ITR. The covalentlylinkedDNA-MBP-Rep68Acomplex (linked) and free DNA probe (free) are indicated by arrows at the left. Since the covalent linkageoccurredatagreatly reducedefficiencyontheA57ITRprobe comparedwith thewtITRprobe, 500fmol of A57 ITR probewasused
inthis experiment, compared with 50fmol of thewtITRprobe.
the formation of alower-mobility band on an
SDS-polyacryl-amide gel upon incubation of MBP-Rep68A with 3'-radiola-beled ITRinatrsendonucleaseassay(Fig. 8A). The substrate
was3' end labeled so that the label would be present on the
fragment which was covalently linked to MBP-Rep68A. The reduced mobility of this complex compared with the unbound fragment would allow for determination of the efficiency of linkage.
The resultsof this experiment (Fig. 8B) indicate that hairpin ITRis cleavedatleast100timesmoreefficiently thanA57ITR.
Because of the inefficiencyoflinkagetothe A57ITRprobe, 10 times the molar amount of probe was added to the reaction mixture compared with the amount ofwt ITR probe. Thus, while the hairpin portion of the ITR does not play a role in binding of Rep to the ITR, its presence does stimulate trs
endonuclease activity.
Previously,DNaseIprotectionby Rep68 has beenreported in the B/B' palindromicregion of the ITR (16). It is possible that interaction with this portion of the ITR could act to
stimulatethe covalentlinkage of Rep68totheDNAsubstrate. However, addition intrans of increasingamountsofasynthetic
hairpin cross arm(Fig.1E),whichwasshowntobe folded into thehairpinstructure by digestion with SmaI, hadno effecton the amountof covalently boundMBP-Rep68A detected. Fur-thermore, addition ofthe hairpin cross arm toan EMSA had
noeffectonbindingand itself could notbind Rep68(datanot
shown).
DISCUSSION
Rep68is acritical proteinin the life cycle of AAV. Previous work has shown that a recombinant version of this protein
produced in E. coli can catalyze all of the known in vitro
activities associated with wt Rep68 (8). This report
demon-strates that an additional function of wt Rep68 is its ability to bind to a linear form of the ITR, A57 ITR, with the same affinity as to wthairpin ITR. MBP-Rep68A and wt Rep68 also appear to have similar specific DNA binding activities. A dissociation constant of approximately 8 x
10-
M was determined for the wt ITR and A57 ITR probes with MBP-Rep68A. However, a truncated version of A57 ITR, A28 ITR, bound 10 times less efficiently than the full-length fragment. Truncation ofA57ITR withDdeI
alsoinhibited binding, which is in agreementwith previouslypublished data (43). Footprint-ing analysis indicated that theDdeI
truncation cuts within the protected region. This observation may explain why truncation withDdeI
resulted in an inhibition of binding activity and indicates the importance of this region in the formation of a stable complex. Extension of A28 ITRwith random sequence restored the ability ofMBP-Rep68Ato bind toA28ITR. Thus, the increase in binding would appear to be the result of nonspecific sequence stabilization ofMBP-Rep68Abinding. It is not clear whether extension of theDdeI-truncated
A57 ITR with random sequence would have the same stabilizing effect or if that sequence is important for stable binding. Mutationof the trs had no effect on binding, which is in agreement with other published data (43).To further define the MBP-Rep68A binding site, we con-structed a series of oligonucleotide probes which spanned the region protected by MBP-Rep78 in a DNase I footprinting experiment. Two 18-bp probes which contained the imperfect (GCTC)4 repeat motif were found to be sufficient for the binding. While these data show that Rep68 is able to bind to nonhairpin derivatives of the ITR and identify some of the features required for binding of Rep68 to DNA, this ability of Rep68 to bind to linear DNA does not appear to correlate with trsendonuclease activity. The covalent linkage of Rep68 to the non-hairpin-containing ITR, A57 ITR, is approximately 100-fold less efficient than linkage to a hairpin-containing ITR. This effect of the hairpin portion of the ITR was observed only when the hairpin was part of the probe, not when it was supplied in trans. This difference was confirmed by the stan-dard trs endonuclease assay (8). Thus, while the hairpin portion of the ITR does not appear to play a role in the stabilization of Rep68 binding, its presence is required for efficient trs endonuclease activity. The features of the hairpin structure that enhance cleavage activity are not clear. The presence of the hairpin structure may force the DNA into a conformation that is more favorable for cleavage. However, previous work showed that digestion of the hairpin structure with the restriction enzyme
SmaI,
which removed most of the C/C' duplex, had little or no effect on trs endonuclease activity by Rep68 (43). Since trs endonuclease activity is thought to be central for the replication of AAV DNA, these data could indicate the importance of structure in this region for efficient trs endonuclease activity.The data presented in this report indicate that a hairpin conformation is not required for binding of wt Rep68 or MBP-Rep68A to the ITR. Footprinting experiments indicate that the
DdeI
site is protected, and therefore truncation at the DdeI site would remove a portion of the binding site and inhibit binding. Studies in the other reports of Rep68 not binding to linear DNA used linear wt ITR as eithera probe or a competitor. This fragment is a more complex substrate, and lack of binding is not as easily interpretable. The linear ITR contains two binding sites for Rep68 in close proximity and may not bind Rep68 because of interference between these sites. Analternatehypothesis is that the palindromes which are normally in a fully hairpin conformation in the wt ITR are nowon November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.53.298.77.176.2]in a linear conformation. This may create a partial hairpin structure that isinhibitoryfor Rep68 binding.
Our observation that Rep canbind tolinear DNA clarifies
some of the effects thatp5-initiated Rep proteins have onthe
expression of heterologous promoters and the apparent cyto-static effects of overexpression of the Rep proteins. These effects maybe duetodirect Rep-DNA interaction withingene
regulatory elements. It has been recently reported that a sequencewithhomologytothe GCTCrepeatmotif ispresent in the longterminal repeat promoter of human immunodefi-ciency virustype 1andmaybeinvolvedinthe inhibitionof this
virus by the Rep proteins (34). The binding of Rep68 and Rep78 to cellular promoters could have a negative effect on gene expression suchas occurswith the p5 andp19 promoter ofAAV2 (2, 3, 24, 27). Binding ofp5Rep proteins could also
havea positive effect and stimulate the expression of silenced genesinthe cell oroverexpression ofgenes involved in down regulation of the cell. Such stimulatory effects of Rep have been reported for the p40 promoter of AAV2 (30, 46).
The results of methylation interference indicated that the guanine residuesinthe GCTCrepeatsof the AAVintegration locus, AAVS1, areinvolved in binding (48). However,what is notknownis the importance of theindividual baseswithin the
repeatsorthecontribution of the flankingsequencein forming astable Rep-DNA complex. Thisinformation isnecessaryfor
definingaconsensus sequencefor Rep68 andidentifyinggenes
thatcould be regulated by Rep.
The observation that binding of MBP-Rep68A is distinct fromtrsendonuclease activity is important whenoneconsiders
the lifecycle ofthevirus. Duringa lytic infection, trs endonu-clease activityhastobe efficienttocomplete thereplication of the large number of viral particles forming in the cell. The inefficienttrsendonuclease activityonnonhairpinordefective
ITRs maybe a way of ensuring that only full-length genomes are replicated. Nicking of the DNA during the process of rescue mayneedtooccuronlyonceandthereforecanaffordto
be inefficient. Furthermore, cleavage of the viral and cellular promoterswhich are Rep responsive is not necessary for its
transcriptionalregulatory effects.
The ability of Rep68 to stably bind to a GCTC repeat sequence present in both the AAV ITR and the integration locuswithout thenecessity ofanadjacentpalindromic hairpin structure allows forspeculation that such bindingmayplayan essential role in theprocess of targeted AAVintegration (8,
48). The presence of stable Rep binding sites at both loci suggestthatRep-Repinteractionmayalign the preintegration site locus with the AAV ITR. It is unclear if other GCTC repeatsequencesidentifiedatother loci arecapableof partic-ipatingasalternateAAVintegration sites. The data presented in this report showing that the hairpin is not required for binding of Rep68 and the identification of the binding site shouldfacilitate theunderstanding of the positive and negative effects of p5-promoted Rep proteins on the expression of
heterologous promoters.
ACKNOWLEDGMENTS
WethankLindaYang forexperttechnical assistance and perform-ing the DNase I footprinting assay and Cathy Ciatto for excellent
administrative skills. We thankNancyNossal for helpful discussion andcriticalreview of this manuscript.
ADDENDUM
A recentpublication has confirmedourearlier observation that Rep68 is able to bind to alinear truncated form of the ITR. McCarty et al. (30a) report that binding to a linear
truncated ITR stem is 125-fold lowerthan to wt ITR, which is contrast to our observations of similar affinities. The data presentedin our report demonstrate that probeswhich do not extend beyond the DdeI site in the stem of the ITR and are less than 46 nucleotides in length do not bind Rep68 compared with the full-length stem. While the A-stem probe used by McCarty et al. is ofsufficient length to allow stable binding, it does not extend beyond the DdeI site and therefore is a poor substrate forRep68bindingcomparedwith thewtITR.
REFERENCES
1. Ashktorab, H., and A. Srivastava. 1989.Identification of nuclear proteins thatspecificallyinteract with adeno-associated virustype 2inverted terminal repeat hairpin DNA. J. Virol. 63:3034-3039. 2. Beaton, A., P. Palumbo, andK. I. Berns. 1989.Expression from
the adeno-associated virus p5 and p19 promoters is negatively regulated in trans by the rep protein. J. Virol. 63:4450-4454. 3. Berns, K.I.,R.M. Kotin, and M. A.Labow. 1988. Regulation of
adeno-associated virus DNA replication. Biochim.Biophys. Acta 951:425-429.
4. Bohenzky, R A., and K.I.Berns. 1989. Interactions between the termini ofadeno-associatedvirus DNA. J.Mol. Biol.206:91-100. 5. Bohenzky, R. A., R. B. LeFebvre, andK.I.Berns. 1988. Sequence and symmetry requirements within the internal palindromic se-quences ofthe adeno-associatedvirusterminal repeat. Virology 166:316-327.
6. Chejanovsky, N., and B. J. Carter. 1989. Mutagenesis of an AUG codon in theadeno-associated virus rep gene: effects on viral DNA replication. Virology 173:120-128.
7. Chejanovsky, N., and B. J. Carter. 1989. Replication of a human parvovirus nonsense mutant in mammalian cells containing an inducible amber suppressor. Virology 171:239-247.
8. Chiorini, J. A., M. D. Weitzman, R A. Owens, E. Urcelay, B. Safer, and R. M. Kotin. 1994. Biologically active Rep proteins of adeno-associated virus type 2 produced as fusion proteins in Escherichiacoli.J. Virol.68:797-804.
9. Fried, M., and D. M. Crothers. 1981. Equilibria and kinetics of lac repressor-operator interactions by polyacrylamide gel electro-phoresis. Nucleic Acids Res. 9:6505-6525.
10. Garner, M. M., and A. Revzin. 1981. A gel electrophoresis method forquantifying the binding of proteins to specific DNA regions: applicationstocomponents of the Escherichia coli lactose operon regulatory system. Nucleic Acids Res. 9:3047-3060.
11. Hermonat, P. L. 1989. The adeno-associated virus Rep78 gene inhibits cellular transformation induced by bovine papillomavirus. Virology172:253-261.
12. Hermonat, P. L. 1991. Inhibition of H-ras expression by the adeno-associated virus Rep78 transformation suppressor gene product. Cancer Res. 51:3373-3377.
13. Hermonat, P. L. 1992. Inhibition of bovine papillomavirus plasmid DNA replication by adeno-associated virus. Virology 189:329-333.
14. Hermonat, P. L., M. A. Labow, R. Wright, K I. Berns, and N. Muzyczka.1984. Genetics ofadeno-associated virus: isolation and preliminary characterization of adeno-associated virus type 2 mutants. J. Virol. 51:329-339.
15. Hong, G., P. Ward, and K I. Berns. 1992. In vitroreplication of adeno-associated virus DNA. Proc. Natl. Acad. Sci. USA 89:4673-4677.
16. Im, D. S., and N. Muzyczka. 1989. Factors that bind to adeno-associated virus terminal repeats. J. Virol. 63:3095-3104. 17. Im, D.-S., and N. Muzyczka. 1990. The AAV origin binding
protein Rep68 is an ATP-dependent site-specific endonuclease with DNAhelicase activity. Cell 61:447-457.
18. Im, D. S., and N. Muzyczka. 1992. Partial purification of adeno-associated virus Rep78,RepS2,andRep4Oand theirbiochemical characterization. J. Virol. 66:1119-1128.
19. Jacob, W. F., T. A. Silverman, R. B.Cohen, and B. Safer. 1989. Identificationand characterization of a novel transcription factor participatingin the expression of eukaryotic initiationfactor 2 a. J. Biol. Chem.264:20372-20384.
20. Khleif, S. N., T. Myers, B. J. Carter, and J. P.Trempe. 1991.
on November 9, 2019 by guest
http://jvi.asm.org/
Inhibition of cellulartransformation by the adeno-associated virus rep gene.Virology181:738-741.
21. Kotin, R M., R M. Linden, and K. I. Berns. 1992. Characteriza-tionof apreferred site onhumanchromosome 19q for integration of adeno-associated virus DNAbynon-homologous recombina-tion. EMBOJ.11:5071-5078.
22. Kotin, R M., J. C. Menninger, D. C. Ward, and K. .Berns. 1991.
Mapping and direct visualization of a region-specific viral DNA integration site on chromosome 19ql3-qter. Genomics 10:831-834.
23. Kotin, R M., M.Siniscalco, R J. Samulski, X. Zhu, L. Hunter, C.A.Laughlin,S.McLaughlin,N.Muzyczka,M.Rocchi, and K I. Berns. 1990. Site-specific integration by adeno-associated virus. Proc.Natl. Acad. Sci.USA87:2211-2215.
24. Kyostio,S. R M., R A.Owens,M. D.Weitzman, B. A. Antoni, N. Chejanovsky,and B.J.Carter. 1994.Analysis of adeno-associated virus(AAV) wild-type andmutantRepproteins fortheirabilities tonegatively regulateAAV p5 andp19mRNA levels. J. Virol. 68:2947-2957.
25. Labow, M. A., and K I. Berns. 1988.Theadeno-associatedvirus rep gene inhibitsreplication of an adeno-associated virus/simian virus40hybridgenomein cos-7cells.J.Virol. 62:1705-1712. 26. Labow, M. A., L. H. Graf, Jr., and K. I. Berns. 1987.
Adeno-associated virus geneexpression inhibits cellulartransformationby heterologousgenes.Mol. Cell. Biol. 7:1320-1325.
27. Labow, M. A., P. L. Hermonat, andK. .Berns.1986.Positiveand
negative autoregulation of the adeno-associated virus type 2 genome.J.Virol. 60:251-258.
28. LeFebvre, R B., and K. .Berns.1984.Uniqueeventsin
parvovi-rusreplication. Microbiol. Sci. 1:163-167.
29. LeFebvre, R. B., S. Riva, and K. .Berns. 1984. Conformation
takesprecedenceover sequence in adeno-associated virusDNA replication. Mol. Cell. Biol.4:1416-1419.
30. McCarty, D. M., M. Christensen, and N. Muzyczka. 1991. Se-quencesrequired for the coordinate induction of adeno-associated virus p19 and p40 promoters by the Rep protein. J. Virol. 65:2936-2945.
30a.McCarty, D. M., J. H. Ryan, S. Zolotukhin, X. Zhou, and N. Muzyczka. 1994. Interaction of the adeno-associated virus Rep protein with a sequence within the A palindrome of the viral terminal repeat. J. Virol. 68:4998-5006.
31. Meisterernst,M., I.Gander,L.Rogge,andE. L Winnacker. 1988. A quantitative analysis of nuclear factor I/DNA interactions. Nucleic Acids Res. 16:4419-4435.
32. Mendelson, E., J. P. Trempe, and B. J. Carter. 1986.Identification of the trans-acting Rep proteins of adeno-associated virus by antibodiesto asyntheticoligopeptide. J.Virol.60:823-832. 33. Ni, T.-H., X. Zhou, D. M. McCarty, I. Zolotukhin, and N.
Muzyczka. 1994. In vitro replication of adeno-associatedvirus DNA.J.Virol.68:1128-1138.
34. Oelze,I., K.Rittner,andG. Sczakiel.1994.Adeno-associatedvirus type 2 repgene-mediated inhibition of basal expression ofhuman immunodeficiency virus type 1 involves its negative regulatory functions. J.Virol. 68:1229-1233.
35. Owens, R. A., and B. J. Carter. 1992. In vitro resolution of adeno-associated virus DNA hairpin termini by wild-type Rep protein is inhibitedby a dominant-negative mutant of rep. J. Virol. 66:1236-1240.
36. Owens, R A., J. P. Trempe, N. Chejanovsky, and B. J. Carter. 1991.Adeno-associatedvirus repproteins produced ininsect and mammalianexpressionsystems:wild-type and dominant-negative mutant proteins bind to the viral replication origin. Virology 184:14-22.
37. Owens, R. A., M. D. Weitzman, S. R M. Kyostio, and B. J. Carter. 1993. Identification of a DNA-binding domain in the amino terminusof adeno-associated virusRepproteins.J.Virol. 67:997-1005.
38. Samulski,RJ., L. S.Chang, and T. Shenk 1987. A recombinant plasmid from whichaninfectious adeno-associated virusgenome can be excised invitro and its use to study viral replication. J. Virol. 61:3096-3101.
39. Samulski,RJ., A.Srivastava,K I.Berns,and N.Muzyczka.1983. Rescue of adeno-associated virus from recombinant plasmids: genecorrection within the terminalrepeatsofAAV.Cell 33:135-143.
40. Senapathy,P., J. D.Tratschin,and B.J. Carter.1984.Replication of adeno-associated virus DNA. Complementation of naturally occurringrep- mutantsbyawild-typegenome or an ori- mutant and correction of terminal palindrome deletions. J. Mol. Biol. 179:1-20.
41. Shelling, A. N., and M. G. Smith. 1994. Targeted integration of transfected andinfected adeno-associated virusvectorscontaining theneomycin resistancegene.Gene Ther.2:1-5.
42. Snyder,R O., D. S. Im, and N. Muzyczka. 1990. Evidence for covalent attachment of the adeno-associated virus (AAV) Rep proteintothe ends of theAAV genome. J. Virol. 64:6204-6213. 43. Snyder, R O., D.-S. Im,T. Ni,X.Xiao, R J. Samulski,andN. Muzyczka. 1993. Features of the adeno-associated virus origin involvedinsubstraterecognition bythe viralRepprotein.J. Virol. 67:6096-6104.
44. Srivastava, A., E. W. Lusby, and K. .Berns. 1983. Nucleotide
sequence and organization of the adeno-associated virus 2 ge-nome.J. Virol. 45:555-564.
45. Tratschin, J. D., L. L. Miller, and B. J. Carter. 1984. Genetic analysisofadeno-associated virus:properties of deletionmutants constructed invitro and evidence for anadeno-associated virus replication function.J.Virol. 51:611-619.
46. Trempe, J. P., and B. J. Carter. 1988. Regulation of adeno-associatedvirus gene expression in 293 cells: control of mRNA abundance andtranslation.J.Virol.62:68-74.
47. Trempe, J. P.,E. Mendelson,and B.J.Carter. 1987. Character-ization ofadeno-associated virus repproteins in humancellsby antibodies raisedagainstrepexpressedinEscherichiacoli. Virol-ogy161:18-28.
48. Weitzman, M. D., S. R M.Kyostio,R M.Kotin,and R A. Owens. 1994.Adeno-associated virus(AAV) Rep proteins mediate
com-plex formation between AAV DNA and itsintegration site in human DNA. Proc. Natl. Acad. Sci. USA 91:5808-5812.
![FIG. 2.gel.withwasbymixtureincubatedexcessshownapproximately2)atureTheshown],additionandareRepMBP-Rep68AITR.Rep68 under the Both wt Rep68 and recombinant MBP-Rep68A bind to A57 (A) Approximately 10,000 cpm of 5'-end-labeled A57 ITR probe incubated with eit](https://thumb-us.123doks.com/thumbv2/123dok_us/1290515.81924/4.612.101.260.92.516/withwasbymixtureincubatedexcessshownapproximately-aturetheshown-additionandarerepmbp-aitr-recombinant-approximately-labeled-incubated.webp)



