Infection after Local Generation of Mucosal IgA
Uri Lopatin,a* Sarah E. Blutt,b,cMargaret E. Conner,b,cBrian L. Kelsalla
Mucosal Immunobiology Section, Laboratory of Molecular Immunology, National Institute of Allergy and Infectious Diseases, NIH, Bethesda, Maryland, USAa; Department
of Molecular Virology and Microbiology, Baylor College of Medicine, Houston, Texas, USAb; Michael E. DeBakey Veterans Affairs Medical Center, Houston, Texas, USAc
Rotavirus is a major cause of pediatric diarrheal illness worldwide. To explore the role of organized intestinal lymphoid tissues in infection by and immunity to rotavirus, lymphotoxin alpha-deficient (LT␣ⴚ/ⴚ) mice that lack Peyer’s patches and mesenteric
lymph nodes were orally infected with murine rotavirus. Systemic rotavirus was cleared within 10 days in both LT␣ⴚ/ⴚand
wild-type mice, and both strains developed early and sustained serum antirotavirus antibody responses. However, unlike wild-wild-type mice, which resolved the intestinal infection within 10 days, LT␣ⴚ/ⴚmice shed fecal virus for approximately 50 days after
inocu-lation. The resolution of fecal virus shedding occurred concurrently with induction of intestinal rotavirus-specific IgA in both mouse strains. Induction of intestinal rotavirus-specific IgA in LT␣ⴚ/ⴚmice correlated with the (late) appearance of
IgA-pro-ducing plasma cells in the small intestine. This, together with the absence of rotavirus-specific serum IgA, implies that secretory rotavirus-specific IgA was produced locally. These findings indicate that serum IgG responses are insufficient and imply that local intestinal IgA responses are important for the clearance of rotavirus from intestinal tissues. Furthermore, they show that while LT␣-dependent lymphoid tissues are important for the generation of IgA-producing B cells in the intestine, they are not absolutely required in the setting of rotavirus infection. Moreover, the induction of local IgA-producing B cell responses can occur late after infection and in an LT␣-independent manner.
R
otavirus is the leading cause of pediatric viral gastroenteritis worldwide. Virtually all children are infected in their first year of life, and rotavirus infection accounted for the deaths of approx-imately 450,000 children under the age of 5 years, primarily in developing countries, in 2008, prior to the introduction of mass vaccination campaigns of yet-unclear efficacy (1). Children with rotavirus gastroenteritis experience a wide spectrum of clinical symptoms ranging from asymptomatic infection to dehydrating diarrhea and vomiting requiring hospitalization. The cause of this variation in disease presentation remains unknown. One possibil-ity is that children experiencing more severe disease may have an altered immune response to the virus. Unfortunately, this is diffi-cult to assess since “normal” immune responses to rotavirus in humans are not yet well characterized.Indeed, much of what is known about rotavirus immunity has been obtained from mice, particularly those lacking specific im-mune system components. SCID and Rag 2 knockout mice, de-void of both T and B lymphocytes, are unable to resolve a primary rotavirus infection (2–5), while mice possessing either B cells or T cells clear a primary rotavirus infection with near-wild-type kinet-ics (2–9). Mice lacking B cells are, however, susceptible to reinfec-tion (3,6,7), indicating that B cells make a significant contribu-tion to protective immunity. In support of a role for B cells in protective immunity, protection from reinfection with rotavirus in humans correlates with the development of a rotavirus-specific intestinal IgA response (10,11). Furthermore, it has recently been shown that IgA in mice is important for clearance of rotavirus infection (12); however, where this IgA is induced is not yet clear, and in particular, the requirement for the gut-associated lym-phoid tissues in IgA responses to rotavirus has not been addressed. Humans and animals infected with rotavirus exhibit both in-testinal and systemic infection (13,14). It has been proposed that infection of gut-associated lymphoid tissues (GALT), which in-clude Peyer’s patches (PPs), mesenteric lymph nodes (MLNs),
and isolated lymphoid follicles (ILFs) in the small intestine and colon may contribute to viral spread (15,16). However, the pre-cise role that the GALT plays in viral dissemination, clearance, and protection from reinfection has not been directly addressed. To explore this role, we investigated the initial site of rotavirus infec-tion in the intestine and in particular the relainfec-tionship of this initial infection to GALT, by immunohistochemical analysis of infected intestinal tissues from mice sacrificed early after infection. Next, we sought to explore the kinetics of infection, as well as the induc-tion of rotavirus-specific antibody responses in the absence of GALT, by infecting lymphotoxin alpha-deficient (LT␣⫺/⫺) mice. These mice lack PPs, ILFs, and MLNs and are notably deficient in their capacity to produce IgA (17–19).
MATERIALS AND METHODS
Mouse strains.BALB/c inbred and CD1 outbred mice (Charles River, Boston, MA) were housed at Baylor College of Medicine (BCM), and LT␣⫺/⫺and C57BL/6 mice (Jackson Laboratory, Bar Harbor, ME) were
housed at the NIH in specific-pathogen-free animal facilities. The day prior to gavage with rotavirus, the mice were transferred to biosafety level 2 (BSL2) or BSL3 facilities, respectively, for the duration of all studies. All experiments were conducted with adult female mice greater than 4 weeks of age. All animal procedures and protocols were carried out in accor-dance with NIH and BCM standard approved procedures, with
institu-Received2 July 2012 Accepted16 October 2012
Published ahead of print24 October 2012
Address correspondence to Brian L. Kelsall, [email protected].
* Present address: Uri Lopatin, Clinical and Translational Research, Gilead Sciences, Foster City, California, USA.
U.L. and S.E.B. contributed equally to this work.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01801-12
on November 7, 2019 by guest
http://jvi.asm.org/
tional review board approval, and under veterinary surveillance. In some experiments, mice were housed individually and cages changed daily to decrease the likelihood of cross infection or reinfection.
Rotavirus infection and sample collection.Murine wild-type rotavi-rus ECwt(gift of Harry Greenberg, Stanford University) was serially pas-saged in suckling CD-1 mice, and virus stocks were produced at BCM by homogenization of intestines as described previously (20). The 50% in-fective dose (ID50) of the ECwtvirus stock was determined for BALB/c, CD1, and C57BL/6 mice (21). Primary and challenge virus inoculation doses were diluted to the indicated ID50in sterile phosphate-buffered saline (PBS) immediately prior to gavage. For confocal studies, mice were gavaged with 105ID
50ECwtin 100l PBS. For serial shedding and rechal-lenge studies, mice were inoculated with 103ID
50of ECwtrotavirus in 100 l PBS (21). On the day of inoculation (day 0) and at the indicated time points, at least two stool pellets were collected from individual mice and stored at⫺80°C until assayed. Blood was collected by either tail bleed or cardiac puncture. Serum was separated by centrifugation in SST Micro-tainer tubes (Becton Dickenson, Franklin Lakes, NJ) per the manufactur-er’s instructions and stored at⫺80°C until analysis.
ELISAs.Rotavirus antigen and rotavirus-specific antibody enzyme-linked immunosorbent assays (ELISAs) were performed in duplicate at BCM. All measurements were performed in linear ranges of the assay, using standard methods (22).
Immunohistochemistry. Tissues were snap-frozen in Tissue-Tek O.C.T. (optimal cutting temperature) compound (Sakura Fintek USA, Inc., Torrance, CA) at BCM, cut into 8-m sections at the NIH, and stained for immunofluorescence microscopy with antibodies against CD11c (clone N418; E-Biosciences, San Diego, CA), rotavirus (rabbit polyclonal antirotavirus) antibody (23), and/or IgA (rat anti-mouse IgA, clone 11-44-2; E-Biosciences, San Diego, CA), using techniques previ-ously described (24). Briefly, slides were fixed with acetone at⫺20°C for 3 min, prior to rehydration in PBS-Tween 20. Endogenous peroxidase was quenched using peroxidase-blocking reagent (Dako North Amer-ica, Inc., Carpinteria, CA) per the manufacturer’s instructions. Sec-tions were blocked with TNB buffer (Invitrogen) and then stained for 1 h with primary antibody. Primary antibodies were detected using horseradish peroxidase (HRP)-conjugated secondary antibodies (Jackson ImmunoResearch, West Grove, PA) and a tyramide fluores-cent-amplification system (Invitrogen, Carlsbad, CA) prior to nuclear staining (Hoechst 33258; Sigma-Aldrich, St. Louis, MO). For double staining, primary HRP was deactivated by peroxidase blocking reagent (Dako), incubated with the second biotinylated secondary antibody for 1 h, washed, incubated with streptavidin-HRP, and detected with fluorescein isothiocyanate (FITC)-tyramide. Slides were mounted with Fluoromount G (Southern Biotechnology Associates, Inc., Bir-mingham, AL) and analyzed by confocal microscopy.
Confocal microscopy.Images were collected on a Leica SP2-UV 405 confocal microscope (Leica Microsystems, Exton, PA) using a 20⫻ nu-merical aperture (NA) 1.25 or 40⫻or 63⫻NA 1.4 oil immersion objec-tive. Fluorochromes were excited using a 488-nm laser for FITC and a 594-nm laser for Alexa 594. DAPI (4=,6-diamidino-2-phenylindole) was excited using a 405-nm diode laser. Detector slits were configured to min-imize diffusion of signal between channels. Images were collected sepa-rately and later superimposed. Images were processed using Leica TCS-SP software (version 2.1537), Imaris 3.3.2 (Bitplane AG, Zurich, Switzer-land), and Adobe Photoshop CS2 (Adobe Systems, San Jose, CA).
Statistics.For all figures, statistics were analyzed using the SPSS Statistics software, version 17.0 (IBM, Inc., Armonk, NY). The Mann-Whitney U non-parametric test was applied to all figures, and data are shown as means⫾ standard errors of the means (SEM).
RESULTS
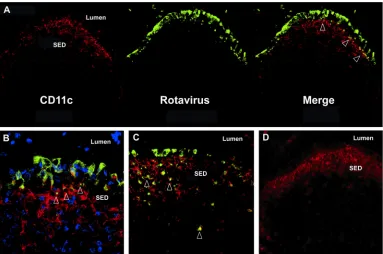
Rotavirus antigen is found in the Peyer’s patch follicle-associ-ated epithelium early in infection and colocalizes with CD11cⴙ cells in the subepithelial cell dome.Previous work in suckling
mice indicates that rotavirus antigens colocalize to the subepithe-lial dome region (SED) of the Peyer’s patches within 72 h after viral inoculation (15). To determine whether dendritic cells in the SED contain rotavirus antigen early after infection, adult CD-1 outbred mice were orally inoculated with murine ECwtrotavirus and PPs were examined by immunohistochemistry. Rotavirus an-tigen was detected in the follicle-associated epithelium (FAE) of the PPs as early as 24 h after inoculation (Fig. 1A). Additionally, on confocal analysis of the FAE, CD11c⫹cells, most likely dendritic cells (DCs), were observed interdigitating with infected epithelial cells (Fig. 1B) during the first 72 h of infection. Colocalization of CD11c⫹cells and rotaviral antigen was primarily restricted to the SED 72 h after inoculation (Fig. 1C, arrows, and data not shown). As expected, beyond 72 h, rotaviral antigen could also be found throughout the intestinal villous epithelium, as well as in associa-tion with CD11c⫹cells in T cell zones of PPs (data not shown).
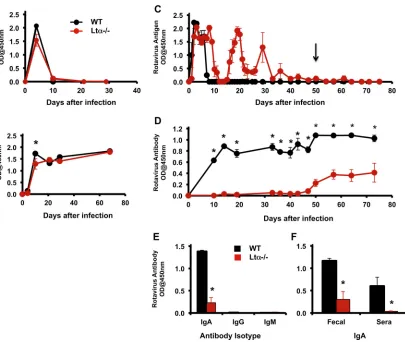
Lymphotoxin alpha-deficient mice are infected with rotavi-rus and clear systemic infection with normal kinetics.Early co-localization of rotavirus with PP CD11c⫹cells suggested that the PPs might play a critical role in extraintestinal dissemination of the virus (25, 26). To examine this possibility, we determined whether the kinetics of systemic rotavirus infection was altered in LT␣⫺/⫺mice that lack PPs, MLNs, and ILFs. We hypothesized that these mice might not become systemically infected or would do so more slowly. Instead, rotavirus antigen was detected in the blood of both LT␣⫺/⫺and wild-type mice within 48 h after viral inoculation but was no longer present in the serum beyond day 10 (Fig. 2Aand data not shown). The decline in rotavirus antigen in the serum of LT␣⫺/⫺mice was associated with an increase in rotavirus-specific serum antibody to levels similar to those of wild-type mice (Fig. 2B). These data indicate that organized mu-cosal lymphoid tissue is not required for systemic viral spread or for the induction of systemic antibody responses.
Lymphotoxin alpha-deficient mice have prolonged intesti-nal virus infection.In addition to evaluating the effect of LT␣ deficiency on systemic rotavirus clearance, we investigated whether the absence of these mucosal immune tissues affected mucosal clearance of rotavirus infection and the generation of rotavirus-specific intestinal IgA. Interestingly, resolution of intes-tinal viral infection was significantly delayed in mice lacking LT␣ expression. While C57BL/6 mice ceased shedding detectable viral antigen in the feces by day 10, LT␣⫺/⫺mice continued to shed significant quantities of virus until day 40, and complete resolu-tion of viral shedding did not occur until approximately day 50 (Fig. 2C). The mean number of time points that were rotavirus antigen positive from 0 to 75 days following primary infection was significantly greater for mice lacking LT␣(mean, 23; range, 22 to 24 time points) than for wild-type mice (mean, 6.5; range, 6 to 7) (P⫽0.018). Resolution of viral shedding was tested by oral chal-lenge of highly susceptible naïve BALB/c mice with fecal pellets from day 50 of “cured” LT␣⫺/⫺mice (Fig. 2C, arrow). None of the BALB/c mice challenged orally became infected (data not shown), confirming the absence of infectious virus.
Lymphotoxin alpha-deficient mice generate a delayed intes-tinal IgA response to rotavirus.Since LT␣⫺/⫺mice exhibited a substantial delay in viral clearance, we next tested for the presence of rotavirus-specific antibodies at multiple time points after viral inoculation. Unlike wild-type mice, which generated rotavirus-specific antibodies in the stool within 10 days of infection, consis-tent with prior studies (13), fecal antirotaviral antibodies were not
on November 7, 2019 by guest
http://jvi.asm.org/
detected in LT␣⫺/⫺mice until approximately 7 weeks postinfec-tion (Fig. 2D). The mean number of days until antibody was first detected was significantly greater for mice lacking LT␣⫺/⫺(mean, 49.3 days; range, 47 to 50) than for wild-type mice (mean, 10 days; range, 10 days) (P⫽0.011). Additionally, when fecal antibody was assayed at week 7 for immunoglobulin isotypes, the only antibody found was IgA in both LT␣⫺/⫺and C57BL/6 mice (Fig. 2E). These data indicated that the PPs and MLNs play a significant role in the early intestinal antibody response to rotavirus infection. However, late after infection, even in the absence of PPs and MLNs in the LT␣⫺/⫺mice, intestinal rotavirus-specific IgA was induced and coincided with the resolution of intestinal rotavirus shedding (Fig. 2CandD).
Since LT␣⫺/⫺mice do not produce significant amounts of IgA, either to commensal bacteria or following immunization (17–19), we suspected that fecal IgA might have been produced in the spleen or bone marrow and secreted into the intestinal lumen across the liver or intestinal epithelium following systemic circu-lation. To test this, serum and feces collected simultaneously were assayed for antirotavirus antibodies at a late time point when viral shedding had declined and antirotavirus IgA was readily detect-able in the stool of both wild-type and LT␣⫺/⫺mice (after day 80). While antirotavirus-specific IgA was detectable in the feces of both control and LT␣⫺/⫺mice (Fig. 2E), at no time point could serum antirotavirus-specific IgA be detected in LT␣⫺/⫺mice (Fig. 2Fand data not shown). This implied that the antirotaviral IgA was pro-duced locally in the intestine. Consistent with this hypothesis, wild-type C57BL/6 mice had abundant IgA-producing plasma cells in the lamina propria (LP) early after infection, a time point where no IgA⫹cells were detectable in the LP of LT␣⫺/⫺mice
(Fig. 3). However, beginning at 5 weeks and persisting through at least 12 weeks after infection, we found IgA-producing plasma cells scattered throughout the LP of LT␣⫺/⫺mice, though fewer than were found in control age-matched C57BL/6 mice (Fig. 3and data not shown). To determine whether LT␣⫺/⫺mice developed protective immunity against secondary infection after primary in-fection resolution, both LT␣⫺/⫺mice and control mice were re-challenged with ECwtat week 7. Neither LT␣⫺/⫺nor control mice shed intestinal rotavirus after challenge (data not shown), and infection did not boost antirotavirus stool IgA levels, confirming the development of protective immunity in LT␣⫺/⫺mice.
DISCUSSION
Gut-associated lymphoid tissues (GALT) are important for the induction of intestinal IgA responses to intestinal pathogens and also play a role in the systemic spread of many intestinal patho-gens, particularly those that enter the body via M cells (25). We first determined the initial site of rotaviral infection in the intes-tine and its relationship to GALT and then explored whether GALT was critical either for the systemic spread of rotavirus or for the induction of rotavirus-specific antibody responses in LT␣⫺/⫺ mice that lack PPs, ILFs, and MLNs. While our data support the idea that a major initial site of rotavirus interaction with GALT is in PPs, with rotavirus antigen found in the FAE in PPs as early as 24 h after infection, they additionally suggest several novel con-clusions.
First, it was clear that local intestinal lymphoid tissues were not required for either local or systemic rotavirus infection. These data suggest that direct apical infection of villus epithelial cells, rather than systemic spread to epithelial cells, is of primary
impor-FIG 1Rotavirus antigens are associated with follicle-associated epithelium and subepithelial dendritic cells early after viral inoculation. Mice were orally inoculated with 105ID
50of rotavirus ECwt, and PP sections were examined by immunofluorescence at 24 h after infection. Rotavirus is shown in green (A to C), CD11c is shown in red (A to D), and cell nuclei are shown in blue (B). Overlap of rotavirus and CD11c is shown in yellow in the merged images, with examples indicated by arrows. A section stained with anti-CD11c and control polyclonal antibody is shown in panel D. All images are representative of results seen with at least three individual mice. A, C, and D, magnification of⫻200; B, magnification of⫻400.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.101.484.66.320.2]tance in establishing intestinal infection and that viremia is not dependent on viral spread from organized intestinal lymphoid tissues.
Second, the direct uptake of rotavirus by CD11c⫹cells, most likely DCs, in the PP has not been widely appreciated. Showing results similar to those of the present study, Dharakul and col-leagues demonstrated the early presence of rotavirus antigen in the murine PPs as early as 3 days postinfection in epithelial cells and Ia⫹CD11b⫺cells in the SED and interfollicular region (IFR) as well as within cells in the MLN over the same time period, which likely represented DCs (15). In the present study, early after infec-tion, CD11c⫹cells appeared to migrate or extend dendrites into the intestinal epithelium. Shortly thereafter, there was clear asso-ciation of rotavirus antigen with CD11c⫹cells in the SED region of the PPs. This likely demonstrates direct infection of epithelial cells in the FAE and uptake by CD11c⫹cells, which is very similar
to the function of CD11c⫹DCs during infection with type 1 reo-virus (27). Whether this uptake by CD11c⫹DCs during homolo-gous rotavirus infection represents uptake of apoptotic cells, as occurs with reovirus (27), or involves direct DC infection is not yet clear. However, Lopez-Guerrero and colleagues did not find rota-virus NSP5 protein associated with CD11c⫹cells in the PPs of adult mice infected with rotavirus, which argues against direct DC infection (28). Recent studies have described a population of CD11c⫹lysozyme-containing cells (“Lyso-DCs”) in murine PPs, which extend dendrites through transcellular pores in M cells to capture antigens, pathogenic bacteria, and dead cells (29, 30). Whether these cells are the same CD11c⫹cells that capture rota-virus antigens is also not known.
Direct uptake of rotavirus by CD11b⫹cells that are not DCs, such as macrophages, within the organized lymphoid follicles may also occur. In this regard, in addition to the studies mentioned
FIG 2LT␣⫺/⫺mice produce systemic rotavirus-specific antibody and clear systemic rotavirus infection, but their production of intestinal IgA and intestinal
clearance of rotavirus are significantly delayed. C57BL/6 (WT, black) and LT␣⫺/⫺(red) mice were orally inoculated with 103ID
50of rotavirus (ECwt). Systemic rotavirus antigens (A) and total rotavirus-specific antibody (including IgA, IgG, and IgM isotypes) in the serum (B) were measured by ELISA. Each point in panels A and B represents the average optical density (OD) reading at 450 nm of samples from four or five individual mice⫾SEM. No significant differences in mean OD values for antigen or antibody were observed at any time point in panel A or B, except for the initial antibody levels at day 10 (P⫽0.043). Fecal levels of rotavirus antigen (C) or antibody (D) were measured over time by ELISA. The mean number of time points that were rotavirus antigen positive (C) and the mean day postinfection that rotavirus-specific antibody was first detected in stool (D) were significantly greater for LT␣⫺/⫺than wild-type mice (Pⱕ0.02). Each
point in panels C and D represents the average OD reading at 450 nm of samples from four or five individual mice⫾SEM. The arrow (C) represents the time point 50 days after infection at which fecal and serum samples were assayed for infectivity (data not shown) and immunoglobulin content (E). Fecal samples from mice infected for 50 days were assessed for the presence of rotavirus-specific IgA, IgG, and IgM (E). Only IgA was detectable. Paired serum and fecal samples from C57BL/6 mice (black bars) and LT␣⫺/⫺mice (red bars) from mice infected forⱖ80 days were assayed for rotavirus-specific IgA (F). Data represent average OD
readings at 450 nm of samples from four individual mice⫾SEM. *,Pⱕ0.05. In LT␣⫺/⫺mice, rotavirus-specific IgA was detected only in fecal samples and not
in any serum samples. Representative data of experiments repeatedⱖ3 times are shown.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.93.498.63.405.2]above, rotavirus antigen has been found within CD11b⫹CD11c⫺ cells that most likely represent macrophages in intestinal lym-phoid tissues (31). Furthermore, bone marrow-derived DCs from mice and monocyte-derived DCs from humans can take up and become activated by rotavirus (32, 33). Whether rotavirus can directly activate DCsin vivo, however, is an important issue, as DCs derivedin vitrofrom cultured bone marrow or blood mono-cytes are quite different from DCs found in tissues (34). Indeed, one recent report indicates that CCR6⫹PP DCs can accumulate and become activated in the SED following rotavirus infection in mice (28). Regardless, the present demonstration that rotavirus antigens are directly taken up by CD11c⫹cells, most likely DCs, in PPs is of interest for understanding the mechanisms by which immune responses are induced to rotavirusin vivo.
Third, the induction of the primary early IgA responses to ro-tavirus was dependent on the presence of LT␣and organized lym-phoid tissues. This early dependence of IgA induction on orga-nized lymphoid structures is not surprising, as B cells producing secretory IgA are induced in PPs and in ILFs, particularly follow-ing infections and immunizations, and LT␣⫺/⫺mice lack these structures (35). In addition, as mentioned above, rotavirus ap-pears to infect the FAE of PPs and can enter the PPs via transport across M cells. Furthermore, rotavirus induces a robust B cell ex-pansion in the PPs of immunocompetent mice (13). Finally, stud-ies show that PP DCs can directly induce IgA B cell switching and differentiation via a mechanism involving their production of
ret-inoic acid (36,37), suggesting that DC presentation of rotavirus to B cells in PPs may be important for the induction of rotavirus-specific IgAin vivo. Therefore, the early induction of IgA to rota-virus appears to require LT␣expression.
Fourth, despite the lack of IgA induced early in rotavirus infec-tion in the LT␣⫺/⫺mice, rotavirus-specific intestinal IgA was pro-duced late after infection, which correlated with both the appear-ance of IgA⫹B cells in the intestinal LP and viral clearance. While prior studies have now demonstrated the importance of IgA for the clearance of rotavirus infection in mice (12), the current data are important in that they provide directin vivoevidence that IgA responses to rotavirus in the intestine can indeed occur in the absence of PPs, MLNs, or ILFs. The source of these local IgA⫹B cells is not yet clear. However, recent data suggest that IgA B cell switching may occur in the intestinal LP, through a process that involves the local production of BAFF/BlyS and APRIL by epithe-lial cells (38,39). It would be of interest to determine whether the late production of IgA in LT␣⫺/⫺ mice is due to direct B cell switching in the LP. If this is the case, then an additional question of interest is why it does not occur at earlier time points. Addition-ally, the fact that LT␣⫺/⫺mice can make IgA responses to this intestinal pathogen is of particular interest given that LT␣⫺/⫺ mice are thought to be IgA deficient and do not make IgA against commensal bacteria (26).
This late IgA could theoretically be derived from serum, as in rodents marginal-zone B cells can contribute to IgA production (39). However, LT␣⫺/⫺mice have been reported to lack marginal-zone B cells (40), serum levels of specific IgA remain remarkably low after infection, and the appearance of IgA in the feces corre-lates with staining for IgA B cells in the intestinal LP. This suggests that late IgA is derived from local IgA-producing plasma cells. Furthermore, the fact that serum antigen was cleared coinciden-tally with the induction of serum antibody responses while intes-tinal infection persisted indicates that serum antibody responses are less likely involved in clearance from the intestine. It is also possible that IgA B cells in this late response are primed within nonmucosal lymphoid tissues and migrate slowly to the intestinal LP, as data support the importance of localization of rotavirus-specific B cells to the intestinal LP for viral clearance (41). Since B cells may obtain specific receptors important for homing to the small intestine (e.g.,␣47 and CCR9) from interaction with den-dritic cells producing retinoic acid in the PPs (and likely the MLNs) (36,37,42), priming of IgA B cells outside these orga-nized mucosal tissues would likely result in lower levels of traf-ficking to, and thus slower accumulation in, the intestine. Where this nonmucosal IgA switching may occur is not clear, but in rodents spleen marginal-zone B cells or peritoneal B1 B cells may contribute to IgA B cell responses in the absence of GALT (39).
Fifth and finally, most murine models evaluating the contribu-tion of B and T cells to clearance of rotaviral infeccontribu-tion have utilized mouse strains deficient in either B cells, T cells, or both. To our knowledge, this is the first time that the immune response to rotavirus has been evaluated in LT␣-deficient mice, which are replete with both B and T cells. In our model, clearance of virus correlated with the induction of antibody responses in the serum and intestine, suggesting that antirotavirus antibody was respon-sible. This is consistent with recent studies with IgA-deficient mice showing that IgA is important for viral clearance (12). However, it is not clear how T cell responses are affected in the LT␣⫺/⫺mice.
FIG 3Late production of antirotavirus IgA in the feces of LT␣⫺/⫺mice
cor-relates with the presence of IgA-positive plasma cells in the intestinal lamina propria. Small intestinal tissue sections from rotavirus-infected mice eutha-nized at day 10 (A, B) or day 60 (C, D) after oral inoculation were stained for IgA (green) and cell nuclei (blue). Sections from wild-type (A) and LT␣⫺/⫺
(B) mice euthanized 10 days after infection indicate that LT␣⫺/⫺mice have no
detectable IgA-secreting plasma cells in their LP within the first 10 days of infection. In contrast, sections from mice 60 days after infection (C, D) indi-cate that IgA-secreting plasma cells are present in small numbers in LT␣⫺/⫺
mice (D), while wild-type mice have plentiful IgA-secreting cells at both time points (A, C). Figures are representative of two mice at each time point.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.44.284.65.309.2]We speculate that CD8⫹T cells contribute to the kinetics of viral clearance seen in the LT␣⫺/⫺mice because mice treated with CD8 and NK cell-depleting antibodies had persistently high viral titers in their stool, which only declined several weeks after infection (data not shown). In comparison, untreated LT␣⫺/⫺mice (replete with CD8 and NK cells) underwent characteristic cyclic declines in viral shedding. The cyclic nature of the rotaviral shedding sug-gested that mice were clearing rotavirus infection and becoming reinfected by other mice in the cage. However, the same pattern was seen with daily cage changes and individualized housing of the mice (data not shown). Other potential explanations include cy-clical viral replication within an individual mouse or cyclic entry (and departure) of CD8 or NK cells into infected LP. At this time, we cannot provide a definitive explanation for this interesting observation, although it does suggest that CD8 and NK responses, despite their ability to contribute to viral control in LT␣⫺/⫺, are not sufficient for durable clearance.
Therefore, the data provided here indicate that rotavirus rap-idly infects the epithelium overlying PPs and give the first direct evidence of rotavirus uptake by DCs in the PPsin vivo. Further-more, while PP infection is not required for either subsequent intestinal epithelial cell or systemic infection by rotavirus, local lymphoid tissues are essential for early induction of secretory an-tibodies that results in viral clearance. In contrast, chronic local infection is sufficient to induce local IgA B cell responses in the absence of organized mucosal tissues late in infection, possibly through a mechanism that involves B cell switching directly in the intestinal lamina or outside the gastrointestinal tract. Finally, sys-temic antibody responses were compartmentalized from local IgA responses to rotavirus and are insufficient for intestinal clearance. These findings have implications for understanding immunity to rotavirus, general mechanisms of IgA induction, and the develop-ment of mucosal vaccines.
ACKNOWLEDGMENTS
This work was supported by the Intramural Research Program of the National Institute of Allergy and Infectious Diseases, National Institutes of Health, National Institute of Allergy and Infectious Diseases grant R01 AI24998 (to M.E.C. and S.E.B) and a merit review grant from the Office of Research and Development, Medical Research Service, Department of Veterans Affairs (to M.E.C. and S.E.B.).
REFERENCES
1.Tate JE, Burton AH, Boschi-Pinto C, Steele AD, Duque J, Parashar UD.
2012. 2008 estimate of worldwide rotavirus-associated mortality in chil-dren younger than 5 years before the introduction of universal rotavirus vaccination programmes: a systematic review and meta-analysis. Lancet Infect. Dis.12:136 –141.
2.Dharakul T, Rott L, Greenberg HB.1990. Recovery from chronic rota-virus infection in mice with severe combined immunodeficiency: rota-virus clearance mediated by adoptive transfer of immune CD8⫹T lympho-cytes. J. Virol.64:4375– 4382.
3.Feng N, Franco MA, Greenberg HB.1997. Murine model of rotavirus infection. Adv. Exp. Med. Biol.412:233–240.
4.Franco MA, Greenberg HB.1997. Immunity to rotavirus in T cell defi-cient mice. Virology238:169 –179.
5.Riepenhoff-Talty M, Dharakul T, Kowalski E, Michalak S, Ogra PL.
1987. Persistent rotavirus infection in mice with severe combined immu-nodeficiency. J. Virol.61:3345–3348.
6.Franco MA, Greenberg HB.1995. Role of B cells and cytotoxic T lym-phocytes in clearance of and immunity to rotavirus infection in mice. J. Virol.69:7800 –7806.
7.Franco MA, Tin C, Greenberg HB. 1997. CD8⫹T cells can mediate almost complete short-term and partial long-term immunity to rotavirus in mice. J. Virol.71:4165– 4170.
8.McNeal MM, Rae MN, Ward RL.1997. Evidence that resolution of rotavirus infection in mice is due to both CD4 and CD8 cell-dependent activities. J. Virol.71:8735– 8742.
9.VanCott JL, McNeal MM, Flint J, Bailey SA, Choi AH, Ward RL.2001. Role for T cell-independent B cell activity in the resolution of primary rotavirus infection in mice. Eur. J. Immunol.31:3380 –3387.
10. Coulson BS, Grimwood K, Hudson IL, Barnes GL, Bishop RF.1992. Role of coproantibody in clinical protection of children during reinfection with rotavirus. J. Clin. Microbiol.30:1678 –1684.
11. Matson DO, O’Ryan ML, Herrera I, Pickering LK, Estes MK.1993. Fecal antibody responses to symptomatic and asymptomatic rotavirus in-fections. J. Infect. Dis.167:577–583.
12. Blutt SE, Miller AD, Salmon SL, Metzger DW, Conner ME.2012. IgA is important for clearance and critical for protection from rotavirus infec-tion. Mucosal Immunol. doi:10.1038/mi. 2012.51.
13. Blutt SE, Kirkwood CD, Parreno V, Warfield KL, Ciarlet M, Estes MK, Bok K, Bishop RF, Conner ME.2003. Rotavirus antigenaemia and vi-raemia: a common event? Lancet362:1445–1449.
14. Blutt SE, Matson DO, Crawford SE, Staat MA, Azimi P, Bennett BL, Piedra PA, Conner ME.2007. Rotavirus antigenemia in children is asso-ciated with viremia. PLoS Med. 4:e121. doi:10.1371/journal. pmed.0040121.
15. Dharakul T, Riepenhoff-Talty M, Albini B, Ogra PL.1988. Distribution of rotavirus antigen in intestinal lymphoid tissues: potential role in devel-opment of the mucosal immune response to rotavirus. Clin. Exp. Immu-nol.74:14 –19.
16. Mossel EC, Ramig RF.2003. A lymphatic mechanism of rotavirus ex-traintestinal spread in the neonatal mouse. J. Virol.77:12352–12356. 17. Davis IA, Rouse BT. 1998. Immune responsiveness of
lymphotoxin-alpha-deficient mice: two reconstitution models. Cell. Immunol.189: 116 –124.
18. Fagarasan S.2008. Evolution, development, mechanism and function of IgA in the gut. Curr. Opin. Immunol.20:170 –177.
19. Macpherson AJ, Gatto D, Sainsbury E, Harriman GR, Hengartner H, Zinkernagel RM.2000. A primitive T cell-independent mechanism of intestinal mucosal IgA responses to commensal bacteria. Science288: 2222–2226.
20. Burns JW, Krishnaney AA, Vo PT, Rouse RV, Anderson LJ, Greenberg HB.1995. Analyses of homologous rotavirus infection in the mouse model. Virology207:143–153.
21. Blutt SE, Warfield KL, O’Neal CM, Estes MK, Conner ME.2006. Host, viral, and vaccine factors that determine protective efficacy induced by rotavirus and virus-like particles (VLPs). Vaccine24:1170 –1179. 22. O’Neal CM, Crawford SE, Estes MK, Conner ME. 1997. Rotavirus
virus-like particles administered mucosally induce protective immunity. J. Virol.71:8707– 8717.
23. Ciarlet M, Crawford SE, Estes MK.2001. Differential infection of polar-ized epithelial cell lines by sialic dependent and sialic acid-independent rotavirus strains. J. Virol.75:11834 –11850.
24. Iwasaki A, Kelsall BL.2000. Localization of distinct Peyer’s patch den-dritic cell subsets and their recruitment by chemokines macrophage in-flammatory protein (MIP)-3alpha, MIP-3beta, and secondary lymphoid organ chemokine. J. Exp. Med.191:1381–1394.
25. Cossart P, Sansonetti PJ.2004. Bacterial invasion: the paradigms of enteroinvasive pathogens. Science304:242–248.
26. Kwa SF, Beverley P, Smith AL.2006. Peyer’s patches are required for the induction of rapid Th1 responses in the gut and mesenteric lymph nodes during an enteric infection. J. Immunol.176:7533–7541.
27. Fleeton MN, Contractor N, Leon F, Wetzel JD, Dermody TS, Kelsall BL.2004. Peyer’s patch dendritic cells process viral antigen from apoptotic epithelial cells in the intestine of reovirus-infected mice. J. Exp. Med.
200:235–245.
28. Lopez-Guerrero DV, Meza-Perez S, Ramirez-Pliego O, Santana-Calderon MA, Espino-Solis P, Gutierrez-Xicotencatl L, Flores-Romo L, Esquivel-Guadarrama FR.2010. Rotavirus infection activates dendritic cells from Peyer’s patches in adult mice. J. Virol.84:1856 –1866. 29. Lelouard H, Fallet M, de Bovis B, Meresse S, Gorvel JP.2012. Peyer’s
patch dendritic cells sample antigens by extending dendrites through M cell-specific transcellular pores. Gastroenterology142:592– 601.e3. 30. Lelouard H, Henri S, De Bovis B, Mugnier B, Chollat-Namy A,
Mal-issen B, Meresse S, Gorvel JP.2010. Pathogenic bacteria and dead cells are internalized by a unique subset of Peyer’s patch dendritic cells that express lysozyme. Gastroenterology138:173–184.e1-3.
on November 7, 2019 by guest
http://jvi.asm.org/
31. Brown KA, Offit PA.1998. Rotavirus-specific proteins are detected in murine macrophages in both intestinal and extraintestinal lymphoid tis-sues. Microb. Pathog.24:327–331.
32. Istrate C, Douagi I, Charpilienne A, McInerney GM, Hidmark A, Johansen K, Larsson M, Magnusson KE, Poncet D, Svensson L, Hinkula J.2007. Bone marrow dendritic cells internalize live RF-81 bovine rotavi-rus and rotavirotavi-rus-like particles (RF 2/6-GFP-VLP and RF 8*2/6/7-VLP) but are only activated by live bovine rotavirus. Scand. J. Immunol.65: 494 –502.
33. Narvaez CF, Angel J, Franco MA.2005. Interaction of rotavirus with human myeloid dendritic cells. J. Virol.79:14526 –14535.
34. Shortman K, Naik SH.2007. Steady-state and inflammatory dendritic-cell development. Nat. Rev. Immunol.7:19 –30.
35. Tsuji M, Suzuki K, Kitamura H, Maruya M, Kinoshita K, Ivanov II, Itoh K, Littman DR, Fagarasan S. 2008. Requirement for lymphoid tissue-inducer cells in isolated follicle formation and T cell-independent immunoglobulin A generation in the gut. Immunity29:261–271. 36. Mora JR, Iwata M, Eksteen B, Song SY, Junt T, Senman B, Otipoby KL,
Yokota A, Takeuchi H, Ricciardi-Castagnoli P, Rajewsky K, Adams DH,
von Andrian UH.2006. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science314:1157–1160.
37. Mora JR, von Andrian UH.2008. Differentiation and homing of IgA-secreting cells. Mucosal Immunol.1:96 –109.
38. Cerutti A.2008. Location, location, location: B-cell differentiation in the gut lamina propria. Mucosal Immunol.1:8 –10.
39. Macpherson AJ, McCoy KD, Johansen FE, Brandtzaeg P.2008. The immune geography of IgA induction and function. Mucosal Immunol.
1:11–22.
40. Banks TA, Rouse BT, Kerley MK, Blair PJ, Godfrey VL, Kuklin NA, Bouley DM, Thomas J, Kanangat S, Mucenski ML.1995. Lymphotoxin-alpha-deficient mice. Effects on secondary lymphoid organ development and humoral immune responsiveness. J. Immunol.155:1685–1693. 41. Youngman KR, Franco MA, Kuklin NA, Rott LS, Butcher EC,
Green-berg HB.2002. Correlation of tissue distribution, developmental pheno-type, and intestinal homing receptor expression of antigen-specific B cells during the murine anti-rotavirus immune response. J. Immunol.168: 2173–2181.
42. Mora JR, von Andrian UH.2009. Role of retinoic acid in the imprinting of gut-homing IgA-secreting cells. Semin. Immunol.21:28 –35.